
Stability of Nuclear and Mitochondrial Reference Genes in Selected Tissues of the Ambrosia Beetle Xylosandrus germanus

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
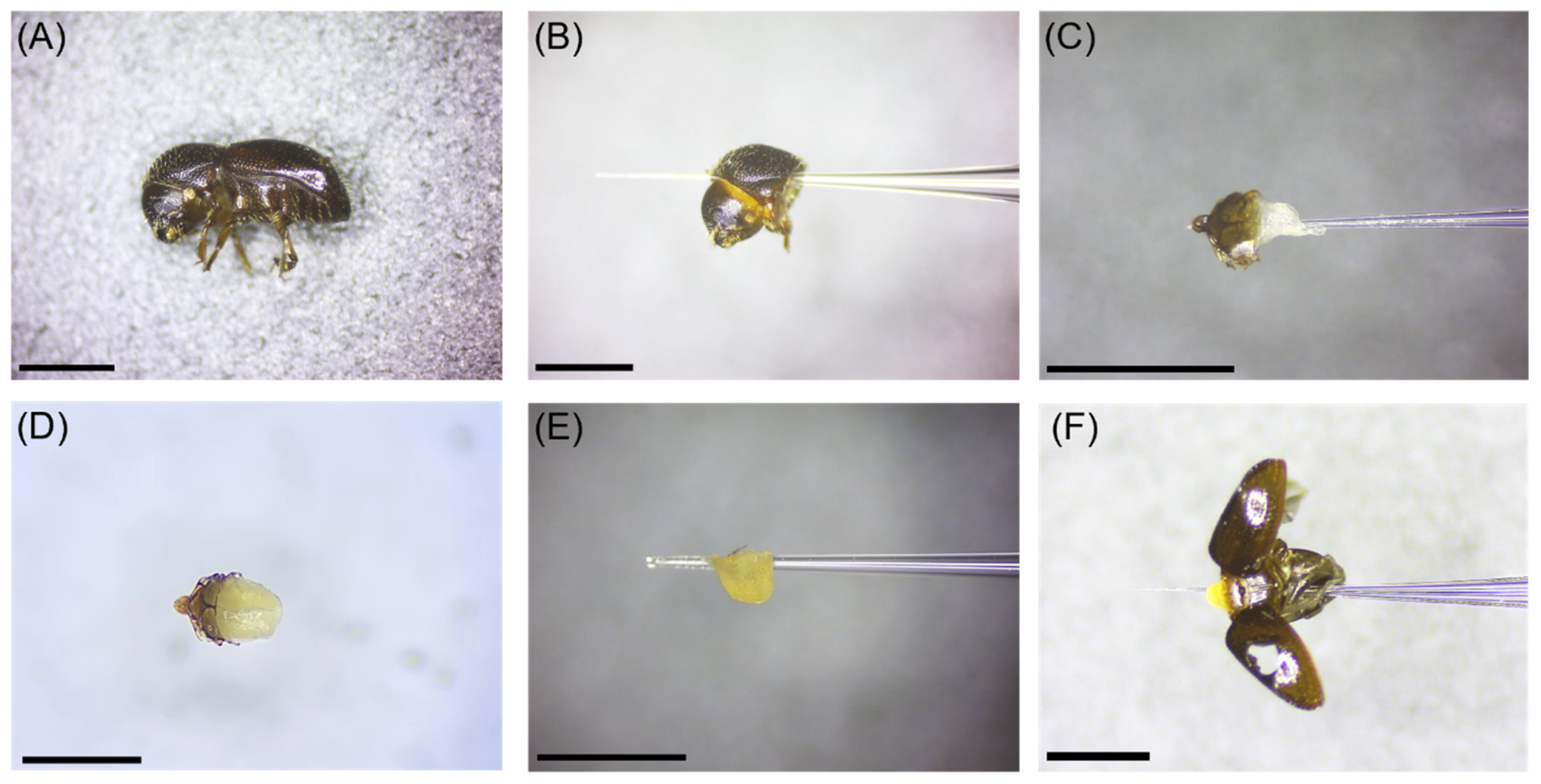
2.1. Insect Specimens and Tissue Types
2.2. RNA Isolation, Reverse Transcription, and Primer Design
2.3. Real-Time PCR
2.4. Stability and Statistical Analysis of Candidate Reference Genes
3. Results and Discussion
3.1. Primer Specificity and Efficiency of Reference Genes
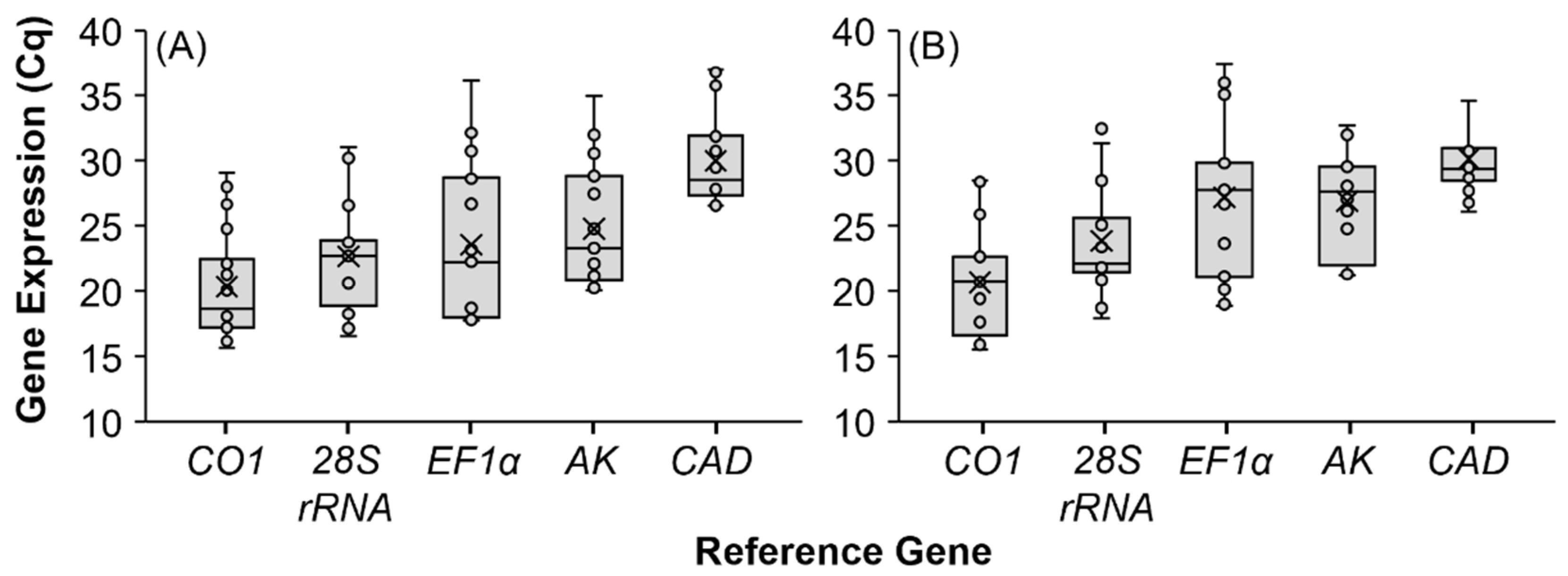
3.2. Expression Profiling of Reference Genes
3.3. Stability Analysis of Reference Genes
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hulcr, J.; Stelinski, L.L. The ambrosia symbiosis: From evolutionary ecology to practical management. Annu. Rev. Entomol. 2017, 62, 285–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranger, C.M.; Reding, M.E.; Schultz, P.B.; Oliver, J.B.; Frank, S.D.; Addesso, K.M.; Chong, J.H.; Sampson, B.; Werle, C.; Gill, S.; et al. Biology, ecology, and management of nonnative ambrosia beetles (Coleoptera: Curculionidae: Scolytinae) in ornamental plant nurseries. J. Integr. Pest Manag. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Agnello, A.M.; Breth, D.I.; Tee, E.M.; Cox, K.D.; Villani, S.M.; Ayer, K.M.; Wallis, A.E.; Donahue, D.J.; Combs, D.B.; Davis, A.E.; et al. Xylosandrus germanus (Coleoptera: Curculionidae: Scolytinae) occurrence, fungal associations, and management trials in New York apple orchards. J. Econ. Entomol. 2017, 110, 2149–2164. [Google Scholar] [CrossRef] [PubMed]
- Monterrosa, A.; Acebes, A.L.; Blaauw, B.; Joseph, S.V. Effects of trap, and ethanol lure type and age on attraction of ambrosia beetles (Coleoptera: Curculionidae). J. Econ. Entomol. 2021, 114, 1647–1654. [Google Scholar] [CrossRef]
- Gomez, D.F.; Rabaglia, R.J.; Fairbanks, K.E.; Hulcr, J. North American Xyleborini north of Mexico: A review and key to genera and species (Coleoptera, Curculionidae, Scolytinae). ZooKeys 2018, 768, 19–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CABI (Commonwealth Agricultural Bureaux International) Invasive Species Compendium. Xylosandrus germanus (Black Timber Bark Beetle); CABI Head Office: Wallingford, UK, 2019; Available online: https://www.cabi.org/isc/datasheet/57237 (accessed on 21 October 2021).
- Mayers, C.G.; McNew, D.L.; Harrington, T.C.; Roeper, R.A.; Fraedrich, S.W.; Biedermann, P.H.; Castrillo, L.A.; Reed, S.E. Three genera in the Ceratocystidaceae are the respective symbionts of three independent lineages of ambrosia beetles with large, complex mycangia. Fungal Biol. 2015, 119, 1075–1092. [Google Scholar] [CrossRef] [Green Version]
- Harrington, T.C.; Aghayeva, D.N.; Fraedrich, S. New combinations in Raffaelea, Ambrosiella, and Hyalorhinocladiella, and four new species from the redbay ambrosia beetle, Xyleborus glabratus. Mycotaxon 2010, 111, 337–361. [Google Scholar] [CrossRef] [Green Version]
- Hulcr, J.; Cognato, A.I. Repeated evolution of crop theft in fungus-farming ambrosia beetles. Evol. Intern. J. Org. Evol. 2010, 64, 3205–3212. [Google Scholar] [CrossRef]
- Li, Y.; Ruan, Y.Y.; Stanley, E.L.; Skelton, J.; Hulcr, J. Plasticity of mycangia in Xylosandrus ambrosia beetles. Insect Sci. 2019, 26, 732–742. [Google Scholar] [CrossRef] [Green Version]
- Kostovcik, M.; Bateman, C.C.; Kolarik, M.; Stelinski, L.L.; Jordal, B.H.; Hulcr, J. The ambrosia symbiosis is specific in some species and promiscuous in others: Evidence from community pyrosequencing. ISME J. 2015, 9, 126–138. [Google Scholar] [CrossRef] [Green Version]
- Ito, M.; Kajimura, H. Landscape-scale genetic differentiation of a mycangial fungus associated with the ambrosia beetle, Xylosandrus germanus (Blandford) (Curculionidae: Scolytinae) in Japan. Ecol. Evol. 2017, 7, 9203–9221. [Google Scholar] [CrossRef] [PubMed]
- Stone, W.D.; Nebeker, T.E.; Monroe, W.A.; MacGown, J.A. Ultrastructure of the mesonotal mycangium of Xylosandrus mutilatus (Coleoptera: Curculionidae). Can. J. Zool. 2007, 85, 232–238. [Google Scholar] [CrossRef]
- Schneider, I.A.; Rudinsky, J.A. Mycetangial glands and their seasonal changes in Gnathotrichus retusus and G. sulcatus. Ann. Entomol. Soc. Am. 1969, 62, 39–43. [Google Scholar] [CrossRef]
- Schneider, I.; Rudinsky, J.A. Anatomical and histological changes in internal organs of adult Trypodendron lineatum, Gnalhotrichus retusus, and G. sulcatus (Coleoptera: Scolytidae). Ann. Entomol. Soc. Am. 1969, 62, 995–1003. [Google Scholar] [CrossRef]
- Norris, D.M. The Mutualistic Fungi of the Xyleborini beetles. In Insect-Fungus Symbiosis: Nutrition, Mutualism, and Commensalism; Batra, L.R., Ed.; Wiley: Hoboken, NY, USA, 1979; p. 5363. [Google Scholar]
- Spahr, E.; Kasson, M.T.; Kijimoto, T. Micro-computed tomography permits enhanced visualization of mycangia across development and between sexes in Euwallacea ambrosia beetles. PLoS ONE 2020, 15, e0236653. [Google Scholar] [CrossRef] [PubMed]
- Joseph, R.; Keyhani, N.O. Fungal mutualisms and pathosystems: Life and death in the ambrosia beetle mycangia. Appl. Microbiol. Biotechnol. 2021, 105, 3393–3410. [Google Scholar] [CrossRef]
- Hruz, T.; Wyss, M.; Docquier, M.; Pfaffl, M.W.; Masanetz, S.; Borghi, L.; Verbrugghe, P.; Kalaydjieva, L.; Bleuler, S.; Laule, O.; et al. RefGenes: Identification of reliable and condition specific reference genes for RT-qPCR data normalization. BMC Genom. 2011, 12, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozera, B.; Rapacz, M. Reference genes in real-time PCR. J. Appl. Gen. 2013, 54, 391–406. [Google Scholar] [CrossRef] [Green Version]
- Castrillo, L.A.; Griggs, M.H.; Vandenberg, J.D. Brood production by Xylosandrus germanus (Coleoptera: Curculionidae) and growth of its fungal symbiont on artificial diet based on sawdust of different tree species. Environ. Entomol. 2012, 41, 822–827. [Google Scholar] [CrossRef]
- Ranger, C.M.; Biedermann, P.H.; Phuntumart, V.; Beligala, G.U.; Ghosh, S.; Palmquist, D.E.; Mueller, R.; Barnett, J.; Schultz, P.B.; Reding, M.E.; et al. Symbiont selection via alcohol benefits fungus farming by ambrosia beetles. Proc. Natl. Acad. Sci. USA 2018, 115, 4447–4452. [Google Scholar] [CrossRef] [Green Version]
- Ranger, C.M.; Reding, M.E.; Persad, A.B.; Herms, D.A. Ability of stress-related volatiles to attract and induce attacks by Xylosandrus germanus and other ambrosia beetles. Agric. For. Entomol. 2010, 12, 177–185. [Google Scholar] [CrossRef]
- Ranger, C.M.; Schultz, P.B.; Frank, S.D.; Chong, J.H.; Reding, M.E. Non-native ambrosia beetles as opportunistic exploiters of living but weakened trees. PLoS ONE 2015, 10, e0131496. [Google Scholar] [CrossRef] [PubMed]
- Dole, S.A.; Jordal, B.H.; Cognato, A.I. Polyphyly of Xylosandrus Reitter inferred from nuclear and mitochondrial genes (Coleoptera: Curculionidae: Scolytinae). Mol. Phylogenet. Evol. 2010, 54, 773–782. [Google Scholar] [CrossRef]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; deCastro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E.; et al. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012, 40, W597–W603. [Google Scholar] [CrossRef]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for general users and for biologist programmers. Methods Mol. Biol. 2000, 132, 365–386. [Google Scholar]
- Radonić, A.; Thulke, S.; Mackay, I.M.; Landt, O.; Siegert, W.; Nitsche, A. Guideline to reference gene selection for quantitative real-time PCR. Biochem. Biophys. Res. Commun. 2004, 313, 856–862. [Google Scholar] [CrossRef]
- Vandesompele, J.; de Preter, K.; Pattyn, F.; Poppe, B.; van Roy, N.; de Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Andersen, C.L.; Jensen, J.L.; Orntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.; Tichopad, A.; Prgomet, C.; Neuvians, T. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 47. [Google Scholar] [CrossRef]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR. BMC Mol. Biol. 2006, 7, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Spiegelaere, W.; Dern-Wieloch, J.; Weigel, R.; Schumacher, V.; Schorle, H.; Nettersheim, D.; Bergmann, M.; Brehm, R.; Kliesch, S.; Vandekerckhove, L.; et al. Reference gene validation for RT-qPCR, a note on different available software packages. PLoS ONE 2015, 10, e0122515. [Google Scholar]
- Lü, J.; Chen, S.; Guo, M.; Ye, C.; Qiu, B.; Wu, J.; Yang, C.; Pan, H. Selection and validation of reference genes for RT-qPCR analysis of the ladybird beetle Henosepilachna vigintioctomaculata. Front. Physiol. 2018, 9, 1614. [Google Scholar] [CrossRef] [Green Version]
- Barros Rodrigues, T.; Khajuria, C.; Wang, H.; Matz, N.; Cardoso, D.C.; Valicente, F.H.; Zhou, X.; Siegfried, B. Validation of reference housekeeping genes for gene expression studies in western corn rootworm (Diabrotica virgifera virgifera). PLoS ONE 2014, 9, e109825. [Google Scholar]
- Kyre, B.R.; Rodrigues, T.B.; Rieske, L.K. RNA interference and validation of reference genes for gene expression analyses using qPCR in southern pine beetle, Dendroctonus frontalis. Sci. Rep. 2019, 9, 5640. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Villalba, A.; Ruijter, J.M.; van den Hoff, M.J. Use and misuse of Cq in qPCR data analysis and reporting. Life 2021, 11, 496. [Google Scholar] [CrossRef] [PubMed]
- Ridgeway, J.A.; Timm, A.E. Reference gene selection for quantitative real-time PCR normalization in larvae of three species of Grapholitini (Lepidoptera: Tortricidae). PLoS ONE 2015, 10, e0129026. [Google Scholar]
- Sang, W.; He, L.; Wang, X.P.; Zhu-Salzman, K.; Lei, C.L. Evaluation of reference genes for RT-qPCR in Tribolium castaneum (Coleoptera: Tenebrionidae) under UVB stress. Environ. Entomol. 2015, 44, 418–425. [Google Scholar] [CrossRef]
- Xue, J.L.; Salem, T.Z.; Turney, C.M.; Cheng, X.W. Strategy of the use of 28S rRNA as a housekeeping gene in real-time quantitative PCR analysis of gene transcription in insect cells infected by viruses. J. Virol. Methods 2010, 163, 210–215. [Google Scholar] [CrossRef]
- Salem, T.Z.; Allam, W.R.; Thiem, S.M. Verifying the stability of selected genes for normalization in Q PCR experiments of Spodoptera frugiperda cells during AcMNPV infection. PLoS ONE 2014, 9, e108516. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.W.; Chen, J.Y.; Lu, M.X.; Gao, Y.; Tian, Z.H.; Gong, W.R.; Zhu, W.; Du, Y.Z. Selection and validation of reference genes for quantitative real-time PCR analysis under different experimental conditions in the leafminer Liriomyza trifolii (Diptera: Agromyzidae). PLoS ONE 2017, 12, e0181862. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Gene Symboland Name | Primer Sequence (5′–3′) | Length (bp) | Efficiency (%) | R2 Regression |
|---|---|---|---|---|
| 28S rRNA 28S Ribosomal RNA | F: ACTCCTTGGTCCGTGTTTCA R: CTCTGGTGACTGTTGGCGA | 120 | 099.0 | 0.998 y = −3.346 + 31.033 |
| AK Arginine kinase | F: ACAAGTCTACCGTCGTCTGG R: GTTGGTTGGGCAGAAAGTGA | 103 | 105.8 | 0.999 y = −3.190 + 30.798 |
| CAD carbamoyl-phosphate synthetase 2-aspartate transcarbamylase-dihydroorotase | F: GAACCACCGCCATAAACGTT R: TGGACAGTCGGGCATTAACT | 120 | 108.7 | 0.998 y = −3.129 + 38.058 |
| CO1 Cytochrome oxidase subunit 1 | F: TTCCTCCTGCTAAAACTGGC R: CCTCAATCCTTGGAGCAATCA | 150 | 104.0 | 0.999 y = −3.229 + 25.483 |
| EF1α Elongation factor-1 alpha | F: CCAACCAGAAATAGGCACGA R: CCACCGAACCACCCTACAG | 119 | 108.3 | 0.999 y = −3.138 + 28.178 |
| Tissue | Source | Gene | GeNorm | NormFinder | BestKeeper | ΔCt | RefFinder | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Stability | Rank | Stability | Rank | Stability | Rank | Stability | Rank | Stability | Final Rank | |||
| Whole Body | LR | CO1 | 0.137 | 1 | 0.069 | 1 | 0.24 | 2 | 0.40 | 1 | 1.19 | 1 |
| EF1α | 0.137 | 1 | 0.069 | 2 | 0.16 | 1 | 0.40 | 2 | 1.41 | 2 | ||
| CAD | 0.215 | 2 | 0.109 | 3 | 0.31 | 4 | 0.44 | 3 | 3.22 | 3 | ||
| AK | 0.305 | 3 | 0.516 | 4 | 0.30 | 3 | 0.58 | 4 | 3.72 | 4 | ||
| 28S | 0.544 | 4 | 0.886 | 5 | 0.91 | 5 | 0.90 | 5 | 5.00 | 5 | ||
| FC | CO1 | 0.136 | 1 | 0.068 | 1 | 0.16 | 2 | 0.78 | 1 | 1.19 | 1 | |
| AK | 0.136 | 1 | 0.389 | 2 | 0.14 | 1 | 0.81 | 2 | 1.41 | 2 | ||
| CAD | 0.870 | 3 | 0.908 | 3 | 1.00 | 4 | 1.06 | 3 | 3.46 | 3 | ||
| EF1α | 0.380 | 2 | 1.051 | 4 | 0.52 | 3 | 1.12 | 4 | 3.46 | 4 | ||
| 28S | 0.985 | 4 | 1.088 | 5 | 1.12 | 5 | 1.16 | 5 | 5.00 | 5 | ||
| Head + Pronotum | LR | CO1 | 0.784 | 2 | 0.468 | 1 | 0.84 | 3 | 1.19 | 1 | 1.73 | 1 |
| CAD | 0.181 | 1 | 1.068 | 3 | 0.21 | 1 | 1.32 | 3 | 1.73 | 2 | ||
| 28S | 0.181 | 1 | 1.264 | 4 | 0.26 | 2 | 1.42 | 4 | 2.38 | 3 | ||
| AK | 1.148 | 3 | 0.759 | 2 | 1.62 | 4 | 1.32 | 2 | 2.83 | 4 | ||
| EF1α | 1.411 | 5 | 1.710 | 5 | 2.19 | 5 | 1.81 | 5 | 5.00 | 5 | ||
| FC | CO1 | 1.308 | 2 | 0.849 | 1 | 1.83 | 3 | 1.85 | 1 | 1.73 | 1 | |
| CAD | 0.279 | 1 | 1.708 | 3 | 0.32 | 2 | 2.09 | 2 | 1.86 | 2 | ||
| 28S | 0.279 | 1 | 2.095 | 4 | 0.12 | 1 | 2.28 | 4 | 2.00 | 3 | ||
| AK | 1.958 | 3 | 1.565 | 2 | 2.60 | 4 | 2.12 | 3 | 3.13 | 4 | ||
| EF1α | 2.165 | 4 | 2.295 | 5 | 3.17 | 5 | 2.48 | 5 | 5.00 | 5 | ||
| Inflated Mycangium + Scutellum | LR | AK | 0.359 | 1 | 0.179 | 1 | 2.43 | 3 | 1.47 | 1 | 1.32 | 1 |
| CO1 | 0.359 | 1 | 0.610 | 2 | 2.71 | 4 | 1.56 | 2 | 2.00 | 2 | ||
| 28S | 1.546 | 3 | 1.251 | 3 | 1.25 | 2 | 1.98 | 3 | 2.91 | 3 | ||
| CAD | 1.985 | 4 | 2.512 | 5 | 0.55 | 1 | 2.64 | 5 | 3.34 | 4 | ||
| EF1α | 0.887 | 2 | 2.136 | 4 | 3.57 | 5 | 2.27 | 4 | 3.94 | 5 | ||
| FC | CAD | 0.315 | 1 | 0.989 | 2 | 0.13 | 1 | 1.21 | 2 | 1.41 | 1 | |
| CO1 | 0.527 | 2 | 0.296 | 1 | 0.60 | 3 | 1.01 | 1 | 1.73 | 2 | ||
| 28S | 0.315 | 1 | 1.092 | 4 | 0.21 | 2 | 1.26 | 3 | 2.21 | 3 | ||
| AK | 1.090 | 3 | 1.063 | 3 | 1.70 | 4 | 1.35 | 4 | 3.72 | 4 | ||
| EF1α | 1.275 | 4 | 1.456 | 5 | 1.90 | 5 | 1.55 | 5 | 5.00 | 5 | ||
| Inflated Mycangium | LR | 28S | 0.494 | 1 | 1.467 | 3 | 0.36 | 1 | 1.98 | 2 | 1.57 | 1 |
| CO1 | 1.161 | 2 | 0.702 | 1 | 0.84 | 3 | 1.71 | 1 | 1.73 | 2 | ||
| CAD | 0.494 | 1 | 1.854 | 4 | 0.50 | 2 | 2.16 | 3 | 2.21 | 3 | ||
| EF1α | 1.696 | 3 | 1.435 | 2 | 1.92 | 4 | 2.24 | 4 | 3.36 | 4 | ||
| AK | 2.216 | 4 | 2.798 | 5 | 2.66 | 5 | 3.00 | 5 | 5.00 | 5 | ||
| FC | CAD | 0.371 | 1 | 0.185 | 1 | 0.40 | 2 | 5.32 | 1 | 1.19 | 1 | |
| AK | 0.371 | 1 | 0.185 | 2 | 0.30 | 1 | 5.38 | 2 | 1.41 | 2 | ||
| CO1 | 0.832 | 2 | 0.338 | 3 | 1.13 | 3 | 5.43 | 3 | 3.00 | 3 | ||
| 28S | 1.029 | 3 | 3.140 | 4 | 1.52 | 4 | 5.81 | 4 | 4.00 | 4 | ||
| EF1α | 8.160 | 4 | 18.846 | 5 | 16.11 | 5 | 18.86 | 5 | 5.00 | 5 | ||
| Deflated Mycangium + Scutellum | LR | AK | 0.361 | 1 | 0.181 | 1 | 0.69 | 3 | 0.75 | 1 | 1.32 | 1 |
| 28S | 0.361 | 1 | 0.588 | 2 | 0.38 | 1 | 0.85 | 2 | 1.41 | 2 | ||
| CO1 | 0.560 | 2 | 0.902 | 4 | 0.45 | 2 | 1.04 | 4 | 3.13 | 3 | ||
| CAD | 0.844 | 3 | 0.723 | 3 | 1.33 | 4 | 0.94 | 3 | 3.46 | 4 | ||
| EF1α | 0.927 | 4 | 0.934 | 5 | 1.37 | 5 | 1.05 | 5 | 5.00 | 5 | ||
| Mycangium + Thorax (without pronotum) + Scutellum + Abdomen 1 | LR | EF1α | 0.121 | 1 | 0.207 | 1 | 0.38 | 3 | 0.52 | 1 | 1.32 | 1 |
| AK | 0.121 | 1 | 0.375 | 3 | 0.31 | 1 | 0.55 | 2 | 1.57 | 2 | ||
| CO1 | 0.138 | 2 | 0.379 | 4 | 0.32 | 2 | 0.56 | 3 | 2.91 | 3 | ||
| CAD | 0.331 | 3 | 0.243 | 2 | 0.77 | 4 | 0.64 | 4 | 3.36 | 4 | ||
| 28S | 0.707 | 4 | 1.253 | 5 | 1.52 | 5 | 1.27 | 5 | 5.00 | 5 | ||
| FC | AK | 0.118 | 1 | 0.047 | 1 | 0.38 | 2 | 0.65 | 1 | 1.19 | 1 | |
| CAD | 0.118 | 1 | 0.144 | 2 | 0.40 | 3 | 0.66 | 2 | 1.86 | 2 | ||
| EF1α | 0.309 | 2 | 0.188 | 3 | 0.29 | 1 | 0.69 | 3 | 2.28 | 3 | ||
| 28S | 0.650 | 3 | 1.053 | 4 | 0.81 | 5 | 1.15 | 4 | 4.23 | 4 | ||
| CO1 | 0.869 | 4 | 1.117 | 5 | 0.64 | 4 | 1.20 | 5 | 4.73 | 5 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patwa, N.; Ranger, C.M.; Lehenberger, M.; Biedermann, P.H.; Reding, M.E. Stability of Nuclear and Mitochondrial Reference Genes in Selected Tissues of the Ambrosia Beetle Xylosandrus germanus. Insects 2021, 12, 1125. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121125
Patwa N, Ranger CM, Lehenberger M, Biedermann PH, Reding ME. Stability of Nuclear and Mitochondrial Reference Genes in Selected Tissues of the Ambrosia Beetle Xylosandrus germanus. Insects. 2021; 12(12):1125. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121125
Chicago/Turabian StylePatwa, Nisha, Christopher M. Ranger, Maximilian Lehenberger, Peter H. Biedermann, and Michael E. Reding. 2021. "Stability of Nuclear and Mitochondrial Reference Genes in Selected Tissues of the Ambrosia Beetle Xylosandrus germanus" Insects 12, no. 12: 1125. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121125