
Diversity and Distribution of Forest Ants (Hymenoptera: Formicidae) in Nepal: Implications for Sustainable Forest Management

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
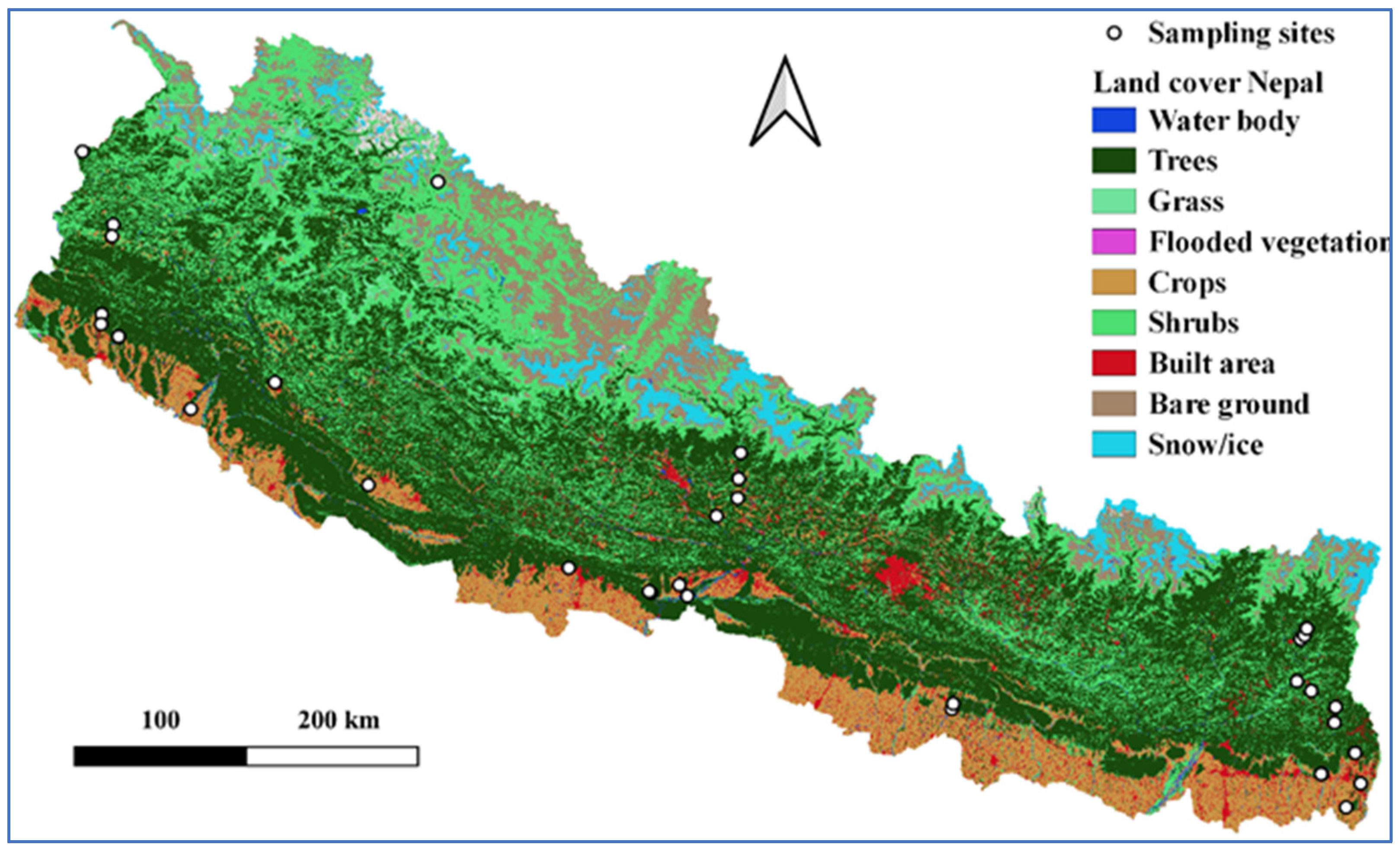
2.1. Field Sampling
2.2. Specimen Processing and Identification
2.3. Data Analysis
3. Results
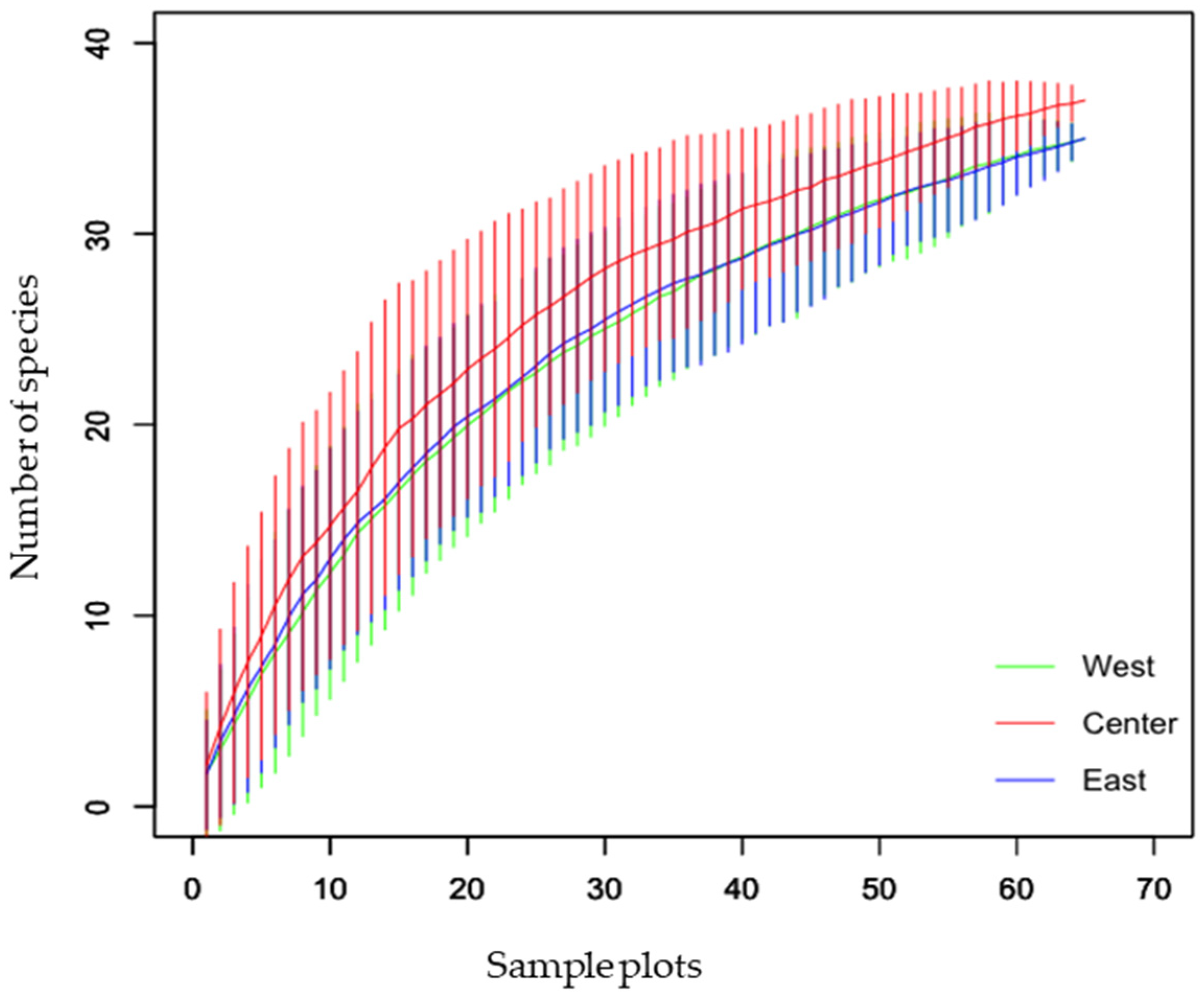
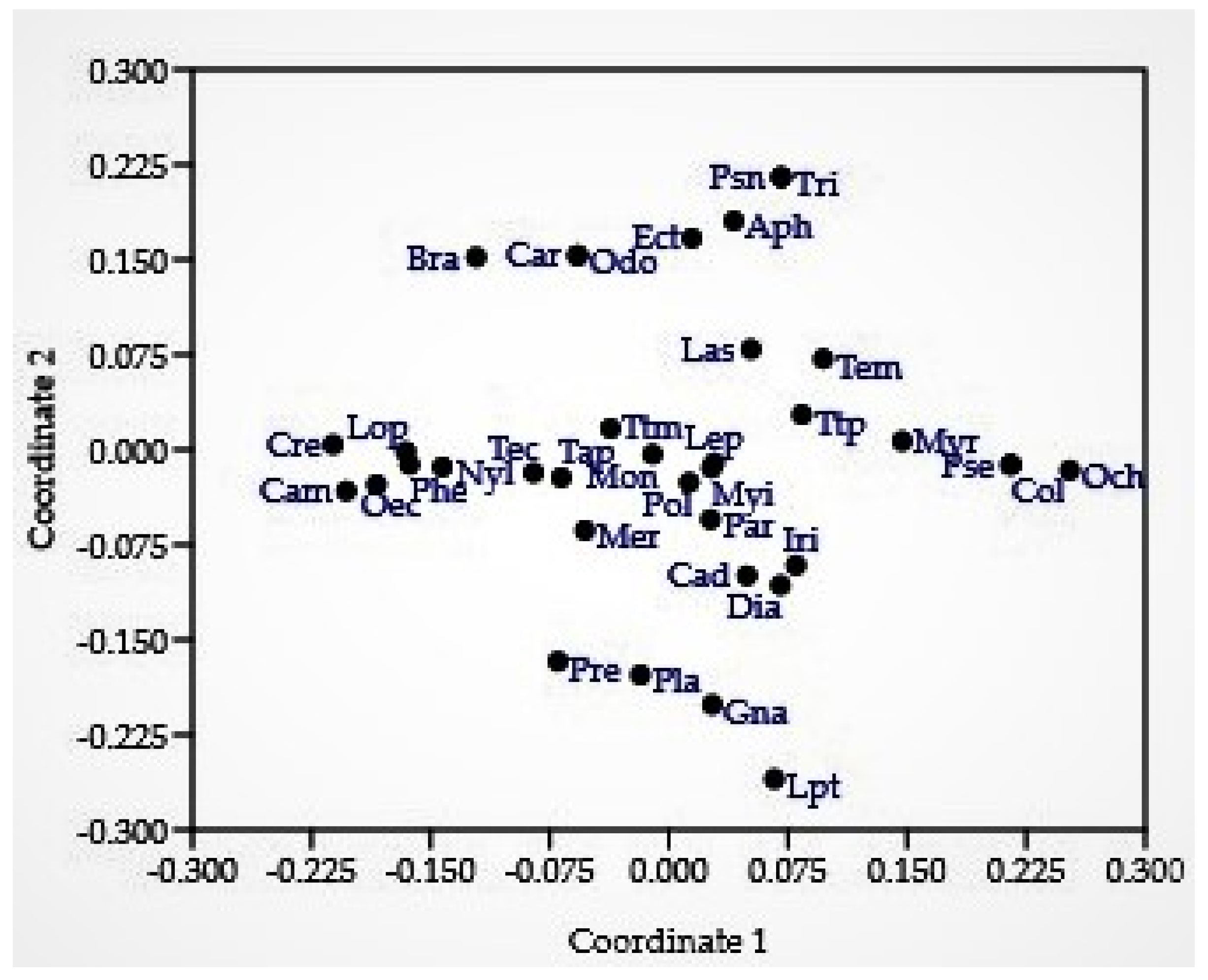
3.1. Ant Species Diversity with New Records for Nepal
3.2. Tramp and Invasive Ant Species
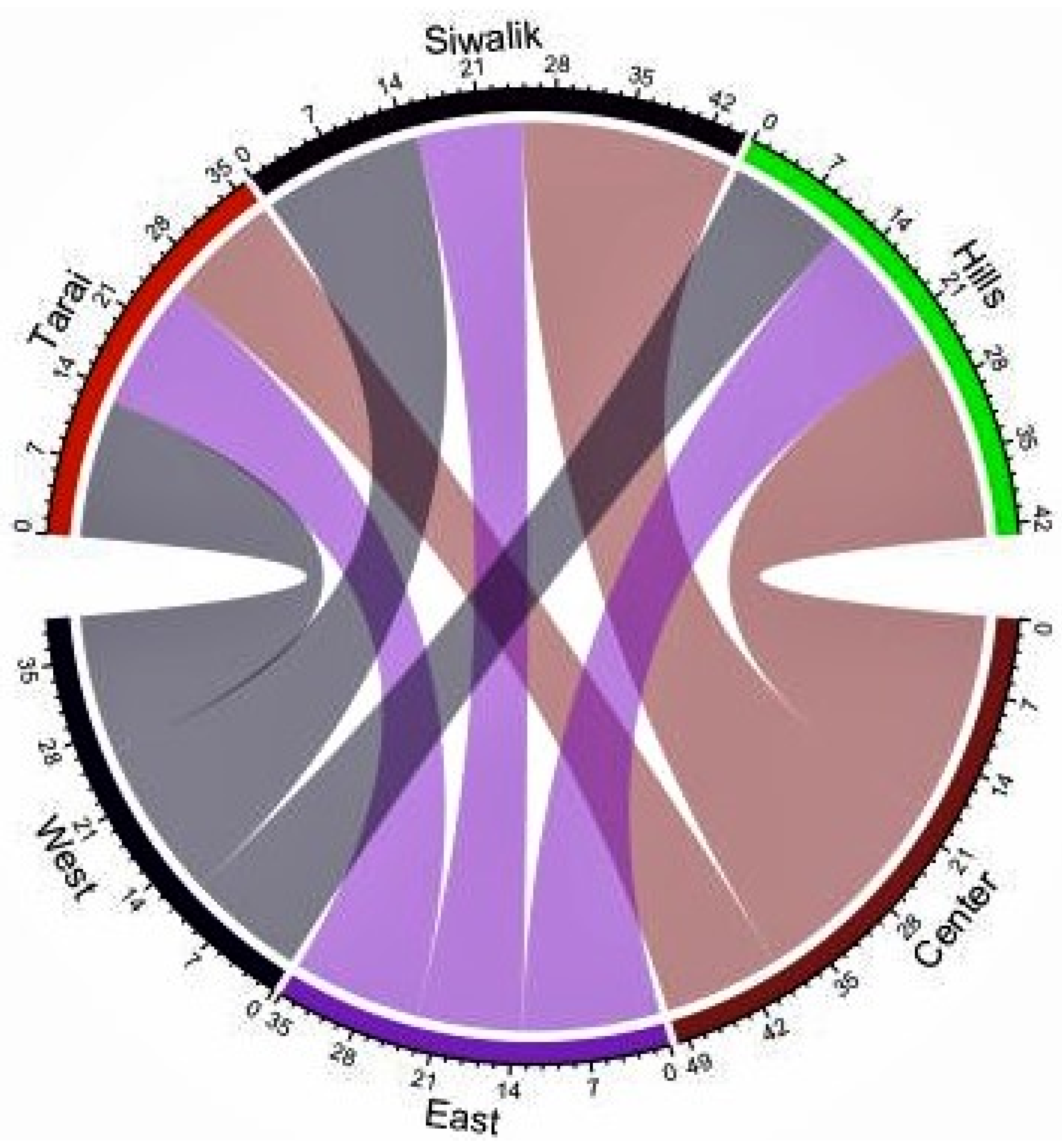
3.3. Distribution of Ant Species
4. Discussion
4.1. Ant Species Diversity
4.2. Distribution of Ants in Nepal
4.3. Ants and Forest Health
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guénard, B. An overview of the species and ecological diversity of ants. In eLS; John Wiley & Sons, Ltd.: Chichester, UK, 2013. [Google Scholar] [CrossRef]
- Thurman, J.H.; Northfield, T.D.; Snyder, W.E. Weaver ants provide ecosystem services to tropical tree crops. Front. Ecol. Evol. 2019, 7, 120. [Google Scholar] [CrossRef] [Green Version]
- Ohyama, L.; King, J.R.; Jenkins, D.G. Are tiny subterranean ants top predators affecting aboveground ant communities? Ecology 2020, 101, e03084. [Google Scholar] [CrossRef]
- Folgarait, P.J. Ant biodiversity and its relationship to ecosystem functioning: A review. Biodivers. Conserv. 1998, 7, 1221–1244. [Google Scholar] [CrossRef]
- Frouz, J.; Jilková, V. The effect of ants on soil properties and processes (Hymenoptera: Formicidae). Myrmecol. News 2008, 11, 191–199. [Google Scholar]
- Kovář, P.; Vojtíšek, P.; Zentsová, I. Ants as ecosystem engineers in natural restoration of human made habitats. J. Landsc. Ecol. 2013, 6, 18–31. [Google Scholar] [CrossRef] [Green Version]
- Warren, R.J.; Giladi, I. Ant-mediated seed dispersal: A few ant species (Hymenoptera: Formicidae) benefit many plants. Myrmecol. News 2014, 20, 129–140. [Google Scholar]
- Anjos, D.V.; Andersen, A.N.; Carvalho, R.L.; Sousa, R.M.; Del-Claro, K. Switching roles from antagonist to mutualist: A harvester ant as a key seed disperser of a myrmecochorous plant. Ecol. Entomol. 2020, 45, 1063–1070. [Google Scholar] [CrossRef]
- Offenberg, J.; Cuc, N.T.T.; Wiwatwitaya, D. The effectiveness of weaver ant (Oecophylla smaragdina) biocontrol in Southeast Asian citrus and mango. Asian Myrmecol. 2013, 5, 139–149. [Google Scholar]
- Wetterer, J.K. Geographic distribution of the weaver ant Oecophylla smaragdina. Asian Myrmecol. 2017, 9, e009004. [Google Scholar]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species: A selection from the Global Invasive Species Database; Invasive Species Specialist Group: Auckland, New Zealand, 2000; Volume 12. [Google Scholar]
- Wilkie, K.T.R.; Mertl, A.L.; Traniello, J.F. Diversity of ground-dwelling ants (Hymenoptera: Formicidae) in primary and secondary forests in Amazonian Ecuador. Myrmecol. News 2009, 12, 139–147. [Google Scholar]
- Stephens, S.S.; Wagner, M.R. Using ground foraging ant (Hymenoptera: Formicidae) functional groups as bioindicators of forest health in northern Arizona ponderosa pine forests. Environ. Entomol. 2006, 35, 937–949. [Google Scholar] [CrossRef] [Green Version]
- Majer, J.D.; Orabi, G.; Bisevac, L. Ants (Hymenoptera: Formicidae) pass the bioindicator scorecard. Myrmecol. News 2007, 10, 69–76. [Google Scholar]
- Marathe, A.; Priyadarsanan, D.R.; Krishnaswamy, J.; Shanker, K. Spatial and climatic variables independently drive elevational gradients in ant species richness in the Eastern Himalaya. PLoS ONE 2020, 15, e0227628. [Google Scholar] [CrossRef]
- Bharti, H.; Sharma, Y.P.; Bharti, M.; Pfeiffer, M. Ant species richness, endemicity and functional groups, along an elevational gradient in the Himalayas. Asian Myrmecol. 2013, 5, 79–101. [Google Scholar]
- Subedi, I.P.; Budha, P.B.; Bharti, H.; Alonso, L. An updated checklist of Nepalese ants (Hymenoptera, Formicidae). ZooKeys 2020, 1006, 99–136. [Google Scholar] [CrossRef] [PubMed]
- Subedi, I.P.; Budha, P.B.; Bharti, H.; Alonso, L.; Yamane, S. First Record of the Ant Subgenus Orthonotomyrmex of the Genus Camponotus from Nepal (Hymenoptera, Formicidae). Zoodiversity 2021, 55, 279–284. [Google Scholar] [CrossRef]
- Forel, A. Les fourmis de l’Himalaya. Bull. De La Société Vaud. Des. Sci. Nat. 1906, 42, 79–94. [Google Scholar]
- Collingwood, C. Formicidae (Hymenoptera: Aculeata) from Nepal. Khumbu Himal 1970, 3, 371–387. [Google Scholar]
- Lindenmayer, D.; Franklin, J.; Fischer, J. General management principles and a checklist of strategies to guide forest biodiversity conservation. Biol. Conserv. 2006, 131, 433–445. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Margules, C.R.; Botkin, D.B. Indicators of biodiversity for ecologically sustainable forest management. Conserv. Biol. 2000, 14, 941–950. [Google Scholar] [CrossRef]
- Junninen, K.; Penttilä, R.; Martikainen, P. Fallen retention aspen trees on clear-cuts can be important habitats for red-listed polypores: A case study in Finland. Biodivers. Conserv. 2007, 16, 475–490. [Google Scholar] [CrossRef]
- Ohsawa, M. Species richness and composition of Curculionidae (Coleoptera) in a conifer plantation, secondary forest, and old-growth forest in the central mountainous region of Japan. Ecol. Res. 2005, 20, 632–645. [Google Scholar] [CrossRef]
- Hartley, M.J. Rationale and methods for conserving biodiversity in plantation forests. For. Ecol. Manag. 2002, 155, 81–95. [Google Scholar] [CrossRef]
- Ferris, R.; Humphrey, J. A review of potential biodiversity indicators for application in British forests. Forestry 1999, 72, 313–328. [Google Scholar] [CrossRef]
- Majer, J. Ants: Bio-indicators of minesite rehabilitation, land-use, and land conservation. Environ. Manage 1983, 7, 375–383. [Google Scholar] [CrossRef]
- Hoffmann, B.D.; Griffiths, A.D.; Andersen, A.N. Responses of ant communities to dry sulfur deposition from mining emissions in semi-arid tropical Australia, with implications for the use of functional groups. Austral Ecol. 2000, 25, 653–663. [Google Scholar] [CrossRef]
- Agosti, D.; Alonso, L.E. The ALL protocol: A standard protocol for the collection of ground-dwelling ants. In Ants. Standard Methods for Measuring and Monitoring Biodiversity; Agosti, D., Majer, J.D., Alonso, L.E., Schultz, T.R., Eds.; Smithsonian Institution Press: Washington, DC, USA, 2000; pp. 204–206. [Google Scholar]
- Alonso, L.E. Ants as indicators of diversity. In Ants. Standard Methods for Measuring and Monitoring Biodiversity; Agosti, D., Majer, J.D., Alonso, L.E., Schultz, T.R., Eds.; Smithsonian Institution Press: Washington, DC, USA, 2000; pp. 80–88. [Google Scholar]
- Underwood, E.C.; Fisher, B.L. The role of ants in conservation monitoring: If, when, and how. Biol. Conserv. 2006, 132, 166–182. [Google Scholar] [CrossRef]
- Ribas, C.R.; Schoereder, J.H. Ant communities, environmental characteristics and their implications for conservation in the Brazilian Pantanal. Biodivers.Conserv. 2007, 16, 1511–1520. [Google Scholar] [CrossRef]
- Holway, D.A.; Lach, L.; Suarez, A.V.; Tsutsui, N.D.; Case, T.J. The causes and consequences of ant invasions. Annu. Rev. Ecol. Syst. 2002, 33, 181–233. [Google Scholar] [CrossRef] [Green Version]
- Karra, K.; Kontigis, C.; Statman-Weil, Z.; Mazzariello, J.; Mathis, M.; Brumby, S. Global land use/land cover with Sentinel-2 and deep learning. In Proceedings of the 2021 IEEE International Geoscience and Remote Sensing Symposium, Brussels, Belgium, 11–16 July 2021; pp. 4704–4707. [Google Scholar]
- QGIS Development Team. QGIS Geographic Information System. Open Source Geospatial Foundation Project. Available online: http://qgis.osgeo.org (accessed on 17 September 2020).
- Bingham, C.T. The Fauna of British India, Including Ceylon and Burma. Hymenoptera, Vol. II. Ants and Cuckoo-Wasps; Taylor and Francis: London, UK, 1903. [Google Scholar]
- Bolton, B. Identification Guide to the Ant Genera of the World; Harvard University Press: Cambridge, MA, USA, 1994. [Google Scholar]
- Schmidt, C.A.; Shattuck, S.O. The higher classification of the ant subfamily Ponerinae (Hymenoptera: Formicidae), with a review of ponerine ecology and behavior. Zootaxa 2014, 3817, 1–242. [Google Scholar] [CrossRef] [Green Version]
- Rigato, F. Revision of the myrmicine ant genus Lophomyrmex, with a review of its taxonomic position (Hymenoptera: Formicidae). Syst. Entomol. 1994, 19, 47–60. [Google Scholar] [CrossRef]
- Ward, P.S. Taxonomy, phylogeny and biogeography of the ant genus Tetraponera (Hymenoptera: Formicidae) in the Oriental and Australian regions. Invertebr. Syst. 2001, 15, 589–665. [Google Scholar] [CrossRef]
- Seifert, B. The ant genus Cardiocondyla (Insecta: Hymenoptera: Formicidae)-a taxonomic revision of the C. elegans, C. bulgarica, C. batesii, C. nuda, C. shuckardi, C. stambuloffii, C. wroughtonii, C. emeryi, and C. minutior species groups. Ann. Naturhist. Mus. Wien. Ser. B Für Bot. Und Zool. 2002, 104, 203–338. [Google Scholar]
- Karmaly, K. A new species and a key to species of Polyrhachis Smith (Hymenoptera: Formicidae) from India. Biosyst. Biodivers. 2004, 40, 539–551. [Google Scholar]
- Radchenko, A.; Elmes, G.W. Myrmica Ants (Hymenoptera: Formicidae) of the Old World; Natura Optima Dux Foundation Warszawa: Warszawa, Poland, 2010; p. 789. [Google Scholar]
- Heterick, B.E.; Shattuck, S. Revision of the ant genus Iridomyrmex (Hymenoptera: Formicidae). Zootaxa 2011, 2845, 1–175. [Google Scholar] [CrossRef]
- Bharti, H.; Kumar, R. Lophomyrmex terraceensis, a new ant species (Hymenoptera: Formicidae) in the bedoti group with a revised key. J. Asia Pac. Entomol. 2012, 15, 265–267. [Google Scholar] [CrossRef]
- Bharti, H.; Kumar, R. Taxonomic studies on genus Tetramorium Mayr (Hymenoptera, Formicidae) with report of two new species and three new records including a tramp species from India with a revised key. Zookeys 2012, 207, 11–35. [Google Scholar] [CrossRef]
- Bharti, H.; Wachkoo, A.A. Two new species of the ant genus Leptogenys (Hymenoptera: Formicidae) from India, with description of a plesiomorphic ergatogyne. Asian Myrmecol. 2013, 5, 11–19. [Google Scholar]
- Bharti, H.; Akbar, S.A. Meranoplus periyarensis, a remarkable new ant species (Hymenoptera: Formicidae) from India. J. Asia Pac. Entomol. 2014, 17, 811–815. [Google Scholar] [CrossRef]
- LaPolla, J.S.; Fisher, B.L. Then there were five: A reexamination of the ant genus Paratrechina (Hymenoptera, Formicidae). ZooKeys 2014, 422, 35–48. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Lattke, J.E.; Shi, F.; Zhou, S. Three new species of the genus Gnamptogenys (Hymenoptera, Formicidae) from southern China with a key to the known Chinese species. J. Hymenop. Res. 2017, 54, 93–112. [Google Scholar] [CrossRef] [Green Version]
- Akbar, S.A.; Bharti, H. A new species of the ant genus Carebara westwood (Hymenoptera: Formicidae) from India. J. Entomol. Res. Soc. 2017, 19, 35–43. [Google Scholar]
- Williams, J.L.; Lapolla, J.S. Two new Prenolepis species (Hymenoptera: Formicidae) from Indomalaya and Australasia, with a redescription of P. dugasi from Vietnam. Zootaxa 2018, 4441, 171–180. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Colwell, R.K. Estimating Species Richness. Biological Diversity: Frontiers in Measurement and Assessment; Oxford University Press: Oxford, UK, 2011; Volume 12, pp. 39–54. [Google Scholar]
- R. Core Team. R: A Language and Environment for Statistical Computing; R Foundation of Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Pfeiffer, M.; Mezger, D.; Hosoishi, S.; Bakhtiar, E.; Kohout, R.J. The Formicidae of Borneo (Insecta: Hymenoptera): A preliminary species list. Asian Myrmecol. 2011, 4, 9–58. [Google Scholar]
- Fellowes, J.R. Ant (Hymenoptera: Formicidae) genera in southern China: Observations on the Oriental-Palaearctic boundary. Myrmecol. Nachr. 2006, 8, 239–249. [Google Scholar]
- Liu, C.; Guénard, B.; Hita Garcia, F.; Yamane, S.; Blanchard, B.; Yang, D.; Economo, E. New records of ant species from Yunnan, China. ZooKeys 2015, 477, 17–78. [Google Scholar] [CrossRef] [Green Version]
- Bharti, H.; Wachkoo, A.A.; Kumar, R. First inventory of ants (Hymenoptera: Formicidae) in northwestern Shivalik, India. Halteres 2017, 8, 33–68. [Google Scholar] [CrossRef]
- Fontanilla, A.M.; Nakamura, A.; Xu, Z.; Cao, M.; Kitching, R.L.; Tang, Y.; Burwell, C.J. Taxonomic and functional ant diversity along tropical, subtropical, and subalpine elevational transects in Southwest China. Insects 2019, 10, 128. [Google Scholar] [CrossRef] [Green Version]
- Wachkoo, A.A.; Akbar, S.A.; Jan, U.; Shah, G.M. Taxonomic inventory of ants (Hymenoptera: Formicidae) in Jammu and Kashmir state. In Biodiversity of the Himalaya: Jammu and Kashmir State; Springer Nature: Basingstoke, UK, 2020; pp. 733–747. [Google Scholar] [CrossRef]
- Wilson, E.O. Which are the most prevalent ant genera. Studia Entomol. 1976, 19, 187–200. [Google Scholar]
- Offenberg, J. Ants as tools in sustainable agriculture. J. Appl. Ecol. 2015, 52, 1197–1205. [Google Scholar] [CrossRef]
- Subedi, I.P.; Budha, P.B. Diversity and distribution patterns of ants along elevational gradients. Nepal. J. Zool. 2020, 4, 44–49. [Google Scholar] [CrossRef]
- Rahbek, C. The role of spatial scale and the perception of large-scale species-richness patterns. Ecol. Lett. 2005, 8, 224–239. [Google Scholar] [CrossRef]
- Brühl, C.A.; Mohamed, M.; Linsenmair, K.E. Altitudinal distribution of leaf litter ants along a transect in primary forests on Mount Kinabalu, Sabah, Malaysia. J. Trop. Ecol. 1999, 15, 265–277. [Google Scholar] [CrossRef] [Green Version]
- Karaman, M.G. Zoogeography, diversity and altitudinal distribution of ants (Hymenoptera: Formicidae) in the Mediterranean and the oro-Mediterranean parts of Montenegro. North West J. Zool. 2011, 7, 26–34. [Google Scholar]
- Burwell, C.J.; Nakamura, A. Distribution of ant species along an altitudinal transect in continuous rainforest in subtropical Queensland, Australia. Mem. Qld. Mus. 2011, 55, 391–411. [Google Scholar]
- Longino, J.T.; Colwell, R.K. Density compensation, species composition, and richness of ants on a neotropical elevational gradient. Ecosphere 2011, 2, 1–20. [Google Scholar] [CrossRef]
- Yusah, K.M.; Turner, E.C.; Yahya, B.E.; Fayle, T.M. An elevational gradient in litter-dwelling ant communities in Imbak Canyon, Sabah, Malaysia. J. Trop. Biol. Conserv. JTBC 2012, 9, 192–199. [Google Scholar]
- Liu, C.; Dudley, K.L.; Xu, Z.H.; Economo, E.P. Mountain metacommunities: Climate and spatial connectivity shape ant diversity in a complex landscape. Ecography 2018, 41, 101–112. [Google Scholar] [CrossRef]
- Bhattarai, K.R.; Vetaas, O.R. Can Rapoport’s rule explain tree species richness along the Himalayan elevation gradient, Nepal? Diver. Distrib. 2006, 12, 373–378. [Google Scholar] [CrossRef]
- Acharya, K.P.; Vetaas, O.R.; Birks, H. Orchid species richness along Himalayan elevational gradients. J. Biogeogr. 2011, 38, 1821–1833. [Google Scholar] [CrossRef]
- Manish, K.; Pandit, M.K. Phylogenetic diversity, structure and diversification patterns of endemic plants along the elevational gradient in the Eastern Himalaya. Plant Ecol. Divers. 2018, 11, 501–513. [Google Scholar] [CrossRef]
- Grau, O.; Grytnes, J.A.; Birks, H. A comparison of altitudinal species richness patterns of bryophytes with other plant groups in Nepal, Central Himalaya. J. Biogeogr. 2007, 34, 1907–1915. [Google Scholar] [CrossRef]
- Baniya, C.B.; Solhøy, T.; Gauslaa, Y.; Palmer, M.W. The elevation gradient of lichen species richness in Nepal. Lichenologist 2010, 42, 83–96. [Google Scholar] [CrossRef]
- Wetterer, J.K. Geographic origin and spread of cosmopolitan ants (Hymenoptera: Formicidae). Halteres 2015, 6, 66–78. [Google Scholar]
- Wetterer, J.K. Worldwide spread of the longhorn crazy ant, Paratrechina longicornis (Hymenoptera: Formicidae). Myrmecol. News 2008, 11, 137–149. [Google Scholar]
- McGlynn, T.P. The worldwide transfer of ants: Geographical distribution and ecological invasions. J. Biogeogr. 1999, 26, 535–548. [Google Scholar] [CrossRef]
- Del Toro, I.; Ribbons, R.R.; Pelini, S.L. The little things that run the world revisited: A review of ant-mediated ecosystem services and disservices (Hymenoptera: Formicidae). Myrmecol. News 2012, 17, 133–146. [Google Scholar]
- Gorb, S.N.; Gorb, E.V. Effects of ant species composition on seed removal in deciduous forest in eastern Europe. Oikos 1999, 84, 110–118. [Google Scholar] [CrossRef]
- Wike, L.D.; Martin, F.D.; Paller, M.H.; Nelson, E.A. Impact of forest seral stage on use of ant communities for rapid assessment of terrestrial ecosystem health. J. Insect Sci. 2010, 10, 77. [Google Scholar] [CrossRef]
- Brener, A.G.F.; Silva, J.F. Leaf-cutting ant nests and soil fertility in a well-drained savanna in western Venezuela. Biotropica 1995, 17, 250–254. [Google Scholar] [CrossRef]
- Knoepp, J.D.; Coleman, D.C.; Crossley, D., Jr.; Clark, J.S. Biological indices of soil quality: An ecosystem case study of their use. For. Ecol. Manag. 2000, 138, 357–368. [Google Scholar] [CrossRef]
- Roychoudhury, N. New record of natural enemies of sal heartwood borer, Hoplocerambyx spinicornis newman (Coleoptera: Cerambycidae). Pestology 2017, 41, 33–38. [Google Scholar]
- Lach, L. Invasive ants: Unwanted partners in ant-plant interactions? Ann. Mo. Bot. Gard. 2003, 90, 91–108. [Google Scholar] [CrossRef]
- Verdinelli, M.; Yakhlef, S.E.B.; Cossu, C.S.; Pilia, O.; Mannu, R. Variability of ant community composition in cork oak woodlands across the Mediterranean region: Implications for forest management. iForest 2017, 10, 707. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, B.D.; Luque, G.M.; Bellard, C.; Holmes, N.D.; Donlan, C.J. Improving invasive ant eradication as a conservation tool: A review. Biol. Conserv. 2016, 198, 37–49. [Google Scholar] [CrossRef]
- MoFE Nepal National REDD+ Strategy (2018–2022); Ministry of Forests and Environment, Government of Nepal, Singh Durbar: Kathmandu, Nepal, 2018.
- Price, P.W.; Denno, R.F.; Eubanks, M.D.; Finke, D.L.; Kaplan, I. Insect Ecology: Behavior, Populations and Communities; Cambridge University Press: Cambridge, UK, 2011; p. 828. [Google Scholar]
- Rossi, N.; Feldhaar, H. Carpenter ants. In Encyclopedia of Social Insects; Springer: Cham, Switzerland, 2020; pp. 973–978. [Google Scholar]
- Malla, R.; Pokharel, K.K. Forest Pests and Pathogens Problem in Different Forest Types of Nepal; Department of Forest Research and Survey: Kathmandu, Nepal, 2018.
- Finley, K.; Chhin, S. Forest health management and detection of invasive forest insects. Resources 2016, 5, 18. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | Sampling Plots | District/s | Forest Types | Latitude (N) | Longitude (E) | Altitude Range (m) |
|---|---|---|---|---|---|---|
| WT | 13 | Kailali | Sal, riverine forests | 28.48313–28.81801 | 80.67581–81.12818 | 143–208 |
| WS | 18 | Kailali, Surkhet, Dang | Eucalyptus plantation, broadleaf mixed, Sal, riverine forests | 28.09533–28.92537 | 80.56907–82.20147 | 172–243 |
| WMM | 24 | Darchula, Dadeldhura | Pine, Alnus, Quercus, deodar forests | 29.345–29.76724 | 80.40099–82.54359 | 722–2239 |
| CT | 15 | Sarlahi, Rupandehi | Eucalyptus plantation, teak plantation, Sal forest | 26.99305–27.69817 | 83.4002–85.69093 | 105–167 |
| CS | 27 | Nawalpur | Sal, Dalbergia sissoo–Acacia catechu, mixed broadleaf riverine forests | 27.55949–27.62469 | 83.87765–84.10534 | 108–204 |
| CMM | 24 | Tanahun, Lamjung | Champ plantation, Sal, Acacia, Schima–Castanopsis, Bombax ceiba, Alnus nepalensis forests | 27.59218–28.33335 | 84.16049–84.40139 | 262–1011 |
| ET | 18 | Jhapa | Sal, teak, mixed broadleaf forests | 26.4746–26.70515 | 87.84115–88.07256 | 87–135 |
| ES | 6 | Ilam | Sal forest | 26.76225–26.76538 | 88.03876–88.04182 | 276–357 |
| EMM | 24 | Ilam, Panchthar | Sal, Schima–Castanopsis, pine, Uttis, Cryptomeria forests | 26.92539–27.18556 | 87.69814–87.93221 | 613–2208 |
| EHM | 18 | Taplejung | Laurel, mixed, oak, Abies forests | 27.36369–27.42682 | 87.72441–87.76557 | 2569–3645 |
| Ant Genera | Individuals Collected | Occurrences | ||||
|---|---|---|---|---|---|---|
| Eastern | Central | Western | Eastern | Central | Western | |
| Crematogaster | 95 | 36 | 77 | 16 | 19 | 7 |
| Camponotus | 11 | 130 | 40 | 28 | 5 | 7 |
| Oecophylla | 14 | 91 | 35 | 19 | 9 | 9 |
| Lophomyrmex | 34 | 24 | 60 | 8 | 15 | 8 |
| Pheidole | 22 | 64 | 30 | 13 | 8 | 11 |
| Nylanderia | 21 | 39 | 28 | 13 | 10 | 15 |
| Technomyrmex | 23 | 18 | 7 | 7 | 11 | 2 |
| Tramp/Invasive Ant Species | Forest Types | Regions |
|---|---|---|
| Brachyponera chinensis | Alnus forest, mixed broadleaf forest, Quercus forest, Sal forest | WT, WS, WMM |
| Cardiocondyla wroughtoni | Champ plantation, Sal forest | CMM |
| Monomorium pharaonis | Champ plantation, Dalbergia sissoo–Acacia catechu forest, Eucalyptus camaldulensis plantation, Sal forest | WT, WS, CMM, ES |
| Ochetellus glaber | Pine forest | EMM |
| Paratrechina longicornis | Sal forest | WT, CS, CMM |
| Tapinoma melanocephalum | Champ plantation, riverine forest, Sal forest, Managed Sal forest, Schima–Castanopsis forest | WS, WT, CS, CMM |
| Tetramorium lanuginosum | Sal forest, Schima-Castanopsis forest | ES, EMM |
| Trichomyrmex destructor | Riverine forest, Sal forest | WT, WS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Subedi, I.P.; Budha, P.B.; Kunwar, R.M.; Charmakar, S.; Ulak, S.; Pradhan, D.K.; Pokharel, Y.P.; Velayudhan, S.T.; Sathyapala, S.; Animon, I. Diversity and Distribution of Forest Ants (Hymenoptera: Formicidae) in Nepal: Implications for Sustainable Forest Management. Insects 2021, 12, 1128. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121128
Subedi IP, Budha PB, Kunwar RM, Charmakar S, Ulak S, Pradhan DK, Pokharel YP, Velayudhan ST, Sathyapala S, Animon I. Diversity and Distribution of Forest Ants (Hymenoptera: Formicidae) in Nepal: Implications for Sustainable Forest Management. Insects. 2021; 12(12):1128. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121128
Chicago/Turabian StyleSubedi, Indra Prasad, Prem Bahadur Budha, Ripu Mardhan Kunwar, Shambhu Charmakar, Sunita Ulak, Dhirendra Kumar Pradhan, Yam Prasad Pokharel, Sajeev Thavalathadathil Velayudhan, Shiroma Sathyapala, and Illias Animon. 2021. "Diversity and Distribution of Forest Ants (Hymenoptera: Formicidae) in Nepal: Implications for Sustainable Forest Management" Insects 12, no. 12: 1128. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121128