
A Comprehensive Review of the Coffee Leaf Miner Leucoptera coffeella (Lepidoptera: Lyonetiidae)—A Major Pest for the Coffee Crop in Brazil and Others Neotropical Countries

 , , ,
, , ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
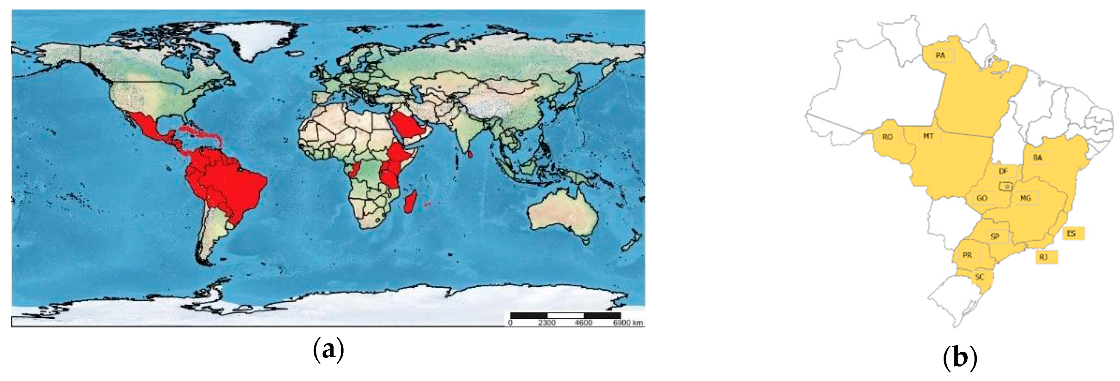
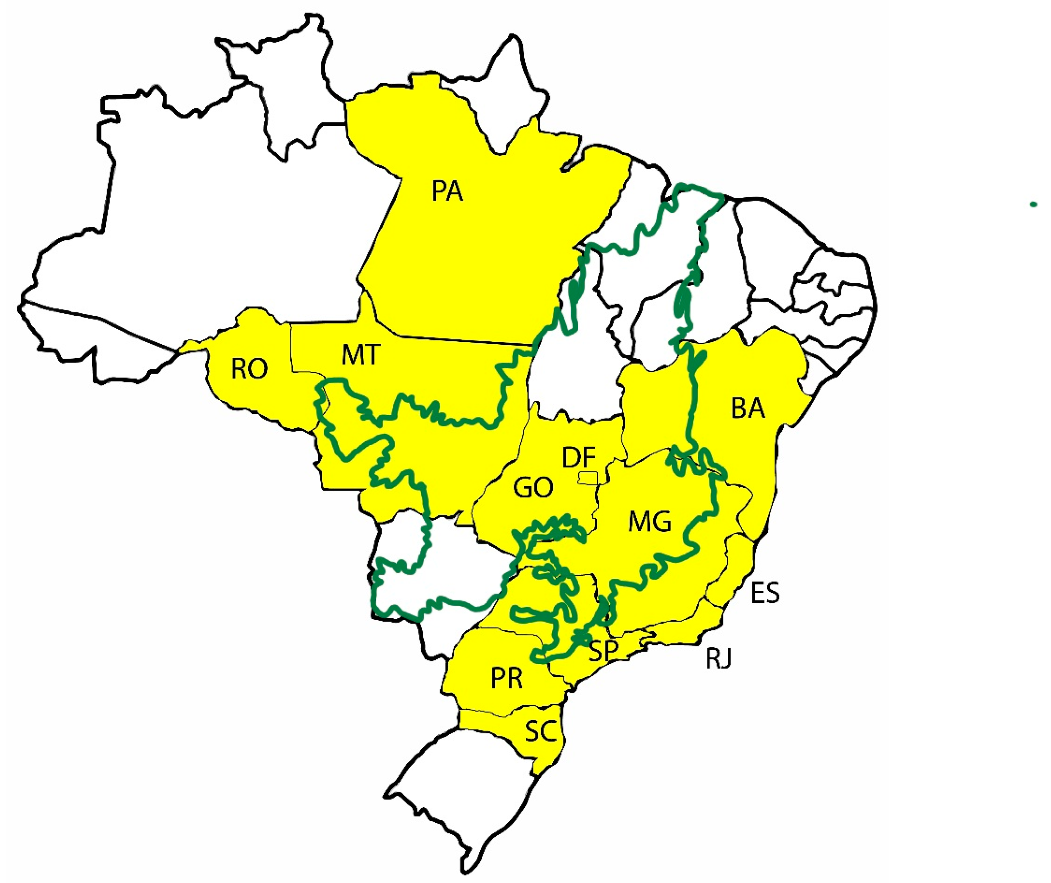
2. History, Origin, and Distribution
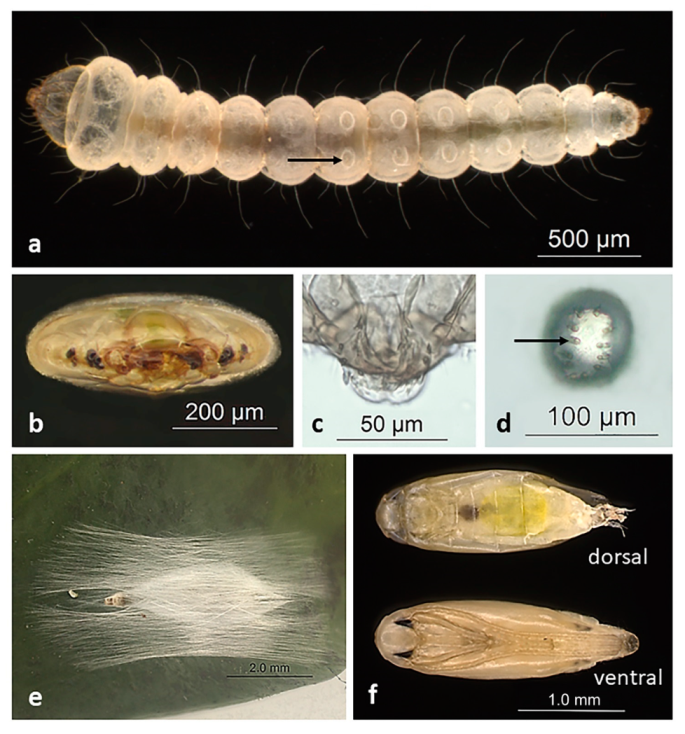
3. Biology
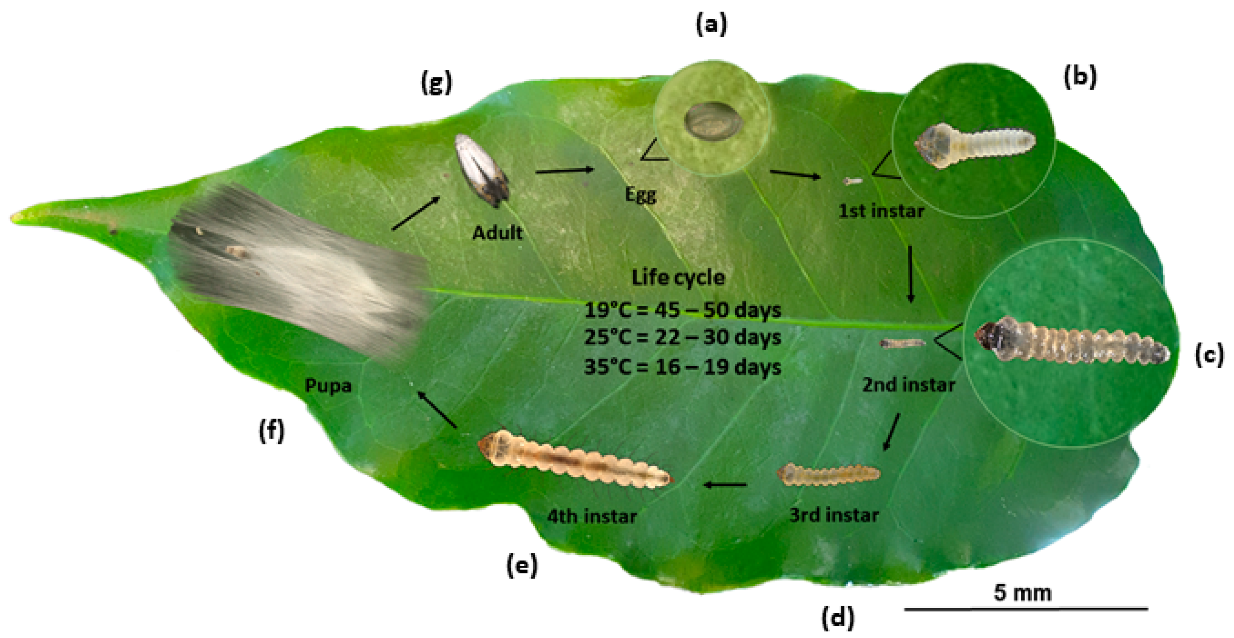
3.1. Life Cycle
3.2. Larval Feeding Behavior
3.3. Adult Behavior
4. Losses Caused by the CLM
5. Control and Management of the CLM
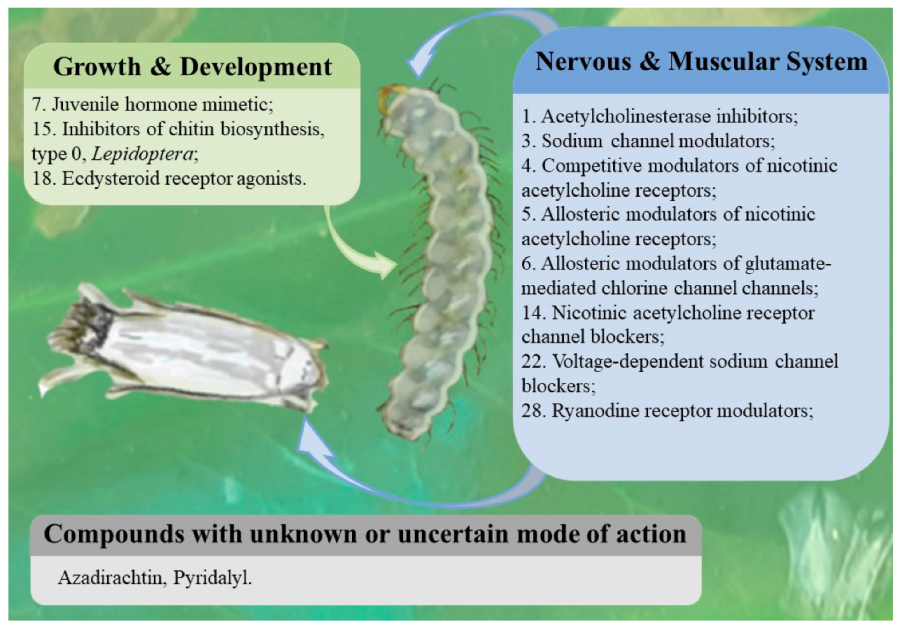
5.1. Chemical Control
5.2. Genetic Resistance
5.3. Biological Control
5.4. Semiochemicals
5.5. Biopesticides
5.5.1. Botanical Pesticides
5.5.2. Nano-Biopesticides
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Species | Country (a) % Parasitism (b) | Reference |
|---|---|---|---|
| Braconidae | Apanteles bordagei | (a) Tanzania (b) 20–75% | [94] |
| Encyrtidae | Ageniaspis sp. | ||
| Eulophidae | Cirrospilus variegatus Closterocerus ritchiei (sin. Achrysocharis ritchiei) Elasmus leucopterae Pediobius coffeicola | ||
| Braconidae Encyrtidae | Apanteles bordagei Parahormius sp. Ageniaspis sp. | (a) Kenia (b) 17–32% | [99] |
| Eulophidae | Closterocerus ritchiei (sin. Achrysocharis ritchiei) Zagrammosoma variegatum Pediobius coffeicola Elasmus sp. Chrysonotomyia sp. | ||
| Braconidae | Mirax insulatris | (a) Puerto Rico (b) 19.5–23.5% | [95] |
| Eulophidae | Achrysocharoides sp. Zagrammosoma spp. Cirrospiloideus sp. Horismenus spp. Chrysonotomyia sp. | ||
| Braconidae | Centistidea striata Orgilus niger Stiropius reticulatus Mirax sp. | (a) Brazil (b) 8–44% | [28,92,97] |
| Eulophidae | Closterocerus coffeellae Cirrospilus sp. Horismenus sp. Neochrysocharis coffeae Proacrias coffeae Tetrastichus sp. | ||
| Braconidae | Allobracon sp. Stiropius letifer | (a) Mexico (b) ≤10% | [96] |
| Eulophidae | Cirrospilus spp. Closterocerus spp. Elachertus spp. Horismenus spp. Miotropis sp. Neochrysocharis spp. Pnigalio spp. Zagrammosoma spp. | ||
| Eulophidae | Zagrammosoma multilineatum Pnigalio sarasolai Closterocerus spp. Horismenus sp. Apleurotropis sp. | (a) Colombia (b) 58–89% | [10] |
References
- USDA. Coffee: World Markets and Trade; United States Department of Agriculture: Washington, DC, USA, 2021.
- Souza, B.; Vázquez, L.L.; Marucci, R.C. Natural Enemies of Insect Pests in Neotropical Agroecosystems: Biological Control and Functional Biodiversity; Springer: Berlin/Heidelberg, Germany, 2019; ISBN 978-3-030-24733-1. [Google Scholar]
- CONAB. Acompanhamento Da Safra Brasileira de Café. Prim. Levant. 2021, 8, 72. [Google Scholar]
- Rena, A.B.; Malavolta, E.; Rocha, M.; Yamada, T. Cultura Do Cafeeiro—Fatores Que Afetam a Produtividade; Associacao Brasileira para Pesquisa da Potassa e do Fosfato: Piracicaba, Brazil, 1986; p. 447. [Google Scholar]
- Parra, J.R.P.; Reis, P.R. Manejo integrado para as principais pragas da cafeicultura, no Brasil. Visão Agríc. 2013, 8, 47–50. [Google Scholar]
- Neves, M.F. Análise Dos Benefícios Econômicos e Sociais Da Utilização Do Carbofurano No Controle de Nematoides, Bicho Mineiro (Leucoptera coffeella) e Cigarra Do Cafeeiro (Quesada Gigas e Fidicina Pronoe) Na Cultura Do Café. Univ. Sao Paulo 2016, 29, 1–29. [Google Scholar]
- de Souza, J.C.; Reis, P.R.; de Oliveira, R.L.; Ciociola Júnior, A.I. Eficiência de Thiamethoxam No Controle do Bicho-Mineiro do Cafeeiro. II—Influência na Época de Aplicação via Irrigação por Gotejamento; SBICafé: Viçosa, Brazil, 2006. [Google Scholar]
- Ramiro Alves, D.; Filho-Guerreiro, O.; Voltan-Queiroz, R.B.; Matthiesen Chebabi, S. Caracterização anatômica de folhas de cafeeiros resistentes e suscetíveis ao bicho-mineiro. Bragantia 2004, 63, 363–372. [Google Scholar] [CrossRef]
- Constantino, L.M.; Flórez, J.C.; Benavides, P.; Bacca, R. Minador de las Hojas del Cafeto: Una Plaga Potencial por Efectos del Cambio Climático; Centro Nacional de Investigaciones de Café (Cenicafé): Caldas, Colômbia, 2013. [Google Scholar]
- David-Rueda, G.; Constantino, L.M.; Montoya, E.C.; Ortega, M.O.E.; Gil, Z.N.; Benavides-Machado, P. Diagnóstico de Leucoptera coffeella (Lepidoptera: Lyonetiidae) y Sus Parasitoides En El Departamento de Antioquia, Colombia. Rev. Colomb. Entomol. 2016, 42, 4–11. [Google Scholar] [CrossRef]
- Leite, S.A.; dos Santos, M.P.; da Costa, D.R.; Moreira, A.A.; Guedes, R.N.C.; Castellani, M.A. Time-Concentration Interplay in Insecticide Resistance among Populations of the Neotropical Coffee Leaf Miner, Leucoptera coffeella. Agric. For. Entomol. 2021, 23, 232–241. [Google Scholar] [CrossRef]
- Giraldo, J.M.; Postali, P.J.R. Biological Aspects of Leucoptera coffeella Guerin Meneville 1842 Lepidoptera: Lyonetiidae in Coffea Arabica under Laboratory Conditions; Cenicafé: Chinchiná, Colombia, 2017; pp. 20–27. [Google Scholar]
- Vega, F.; Posada-Florez, F.; Infante, F. Coffee Insects: Ecology and Control. In Encyclopedia of Pest Managment; CRC Press Taylor & Francis Group: London, UK, 2006; pp. 1–4. [Google Scholar]
- Souto, L.A. Ocorrência de Leucoptera coffeella e Detecção Da Presença de Minas Comparando Amostragem Convencional e Amostragem Por Fotogrametria Terrestre; Repositório Institucional, Universidade Federal de Uberlândia—Instituto de Ciências Agrárias (ICIAG): Uberlândia, Brazil, 2019; p. 27. [Google Scholar]
- Vasconcelos, G.C. Identificação da praga bicho-mineiro em plantações de café usando imagens aéreas e deep learning. Univ. Uberlandia 2019, 30, 1–30. [Google Scholar]
- Leite, S.A.; Guedes, R.N.C.; dos Santos, M.P.; da Costa, D.R.; Moreira, A.A.; Matsumoto, S.N.; Lemos, O.L.; Castellani, M.A. Profile of Coffee Crops and Management of the Neotropical Coffee Leaf Miner, Leucoptera coffeella. Sustainability 2020, 12, 8011. [Google Scholar] [CrossRef]
- da Consolação Rosado, M.; de Araújo, G.J.; Pallini, A.; Venzon, M. Cover Crop Intercropping Increases Biological Control in Coffee Crops. Biol. Control 2021, 160, 104675. [Google Scholar] [CrossRef]
- Valerio, D.R.; Jiménez, F.G. Manejo del minador de la hoja (Leucoptera coffeella) en el cultivo de café en Costa Rica. Agron. Costarric. Rev. Cienc. Agríc. 2021, 45, 143–153. [Google Scholar]
- Guastella, D.; Massimino Cocuzza, G.E.; Rapisarda, C. Integrated Pest Management. In Integrated Pest Management in Tropical Regions; CAB International: Oxfordshire, UK, 2017; pp. 285–312. [Google Scholar]
- Świtek, S.; Sawinska, Z. Farmer Rationality and the Adoption of Greening Practices in Poland. Sci. Agric. 2017, 74, 275–284. [Google Scholar] [CrossRef]
- Sawinska, Z.; Świtek, S.; Głowicka-Wołoszyn, R.; Kowalczewski, P.Ł. Agricultural Practice in Poland Before and After Mandatory IPM Implementation by the European Union. Sustainability 2020, 12, 1107. [Google Scholar] [CrossRef] [Green Version]
- Guérin-Méneville, F.-É.; Perrottet, S. Mémoire sur un Insecte et un Champignon qui Ravagent les Cafiers aux Antilles; Bouchard-Huzard: Paris, France, 1842; pp. 6–30. [Google Scholar]
- Box, H.E. The Bionomics of the White Coffee-Leaf Miner, Leucoptera coffeella, Guér., in Kenya Colony. (Lepidoptera, Lyonetidae.). Bull. Entomol. Res. 1923, 14, 133–145. [Google Scholar] [CrossRef]
- Gallo, D.O.; Nakano, S.S.; Neto, R.P.L.; Carvalho, G.C.; Batista, E.B.; Filho, J.R.P.; Parra, R.A.; Zucchi, S.B.; Alves, J.D. Vendramim Manual de Entomologia AgrÌcola; Agronômica Ceres: Ouro Fino, Brazil, 1988; p. 649. [Google Scholar]
- Silvestri, F. Compendio di Entomologica Applicata (Agraria, Forestale, Medica, Veterinaria); Bellavista: Portici, Italy, 1943; p. 512. [Google Scholar]
- Green, G. A Proposed Origin of the Coffee Leaf-Miner, Leucoptera coffeella (Guerin-Meneville) (Lepidoptera:Lyonetiidae). Bull. Entomol. Soc. Am. 1984, 30, 30–31. [Google Scholar]
- Mey, W. Taxonomische Bearbeitung Der Westpaläarktischen Arten Der Gattung Leucoptera Hübner, ‘1825′, s. 1. (Lepidoptera, Lyonetiidae) ‘Taxonomic Revision of the Westpalaearctic Species of the Genus Leucoptera Hübner, “1825”, s. I. (Lepidoptera, Lyonetiidae)’. Dtsch. Entomol. Z. 1994, 41, 173–234. [Google Scholar] [CrossRef]
- Pereira, E.J.G.; Picanço, M.C.; Bacci, L.; Lucia, T.M.C.D.; Silva, É.M.; Fernandes, F.L. Natural Mortality Factors of Leucoptera coffeella (Lepidoptera: Lyonetiidae) on Coffea Arabica. Biocontrol. Sci. Technol. 2007, 17, 441–455. [Google Scholar] [CrossRef]
- Medina Filho, H.P.; Carvalho, A.; Mônaco, L.C. Melhoramento do cafeeiro: XXXVII—Observações sobre a resistência do cafeeiro ao bicho-mineiro. Bragantia 1977, 36, 131–137. [Google Scholar] [CrossRef] [Green Version]
- Ghini, R.; Hamada, E.; Pedro Júnior, M.J.; Marengo, J.A.; Gonçalves, R.R.D.V. Risk Analysis of Climate Change on Coffee Nematodes and Leaf Miner in Brazil. Pesqui. Agropecuária Bras. 2008, 43, 187–194. [Google Scholar] [CrossRef] [Green Version]
- Pantoja-Gomez, L.M.; Corrêa, A.S.; de Oliveira, L.O.; Guedes, R.N.C. Common Origin of Brazilian and Colombian Populations of the Neotropical Coffee Leaf Miner, Leucoptera coffeella (Lepidoptera: Lyonetiidae). J. Econ. Entomol. 2019, 112, 924–931. [Google Scholar] [CrossRef]
- Parra, J.R.P.; Gonçalves, W.; Precetti, A.A.C.M. Flutuação Populacional de Parasitos e Predadores de Perileucoptera coffeella (Guérin-Méneville, 1842) Em Três Localidades Do Estado de São Paulo. Turrialba 1981, 31, 357–364. [Google Scholar]
- de Souza, J.C.; Reis, P.R.; Rigitano, R.D.O. Bicho-Mineiro Do Cafeeiro: Biologia, Danos e Manejo Integrado; EPAMIG: Belo Horizonte, Brazil, 1998; pp. 48–54. [Google Scholar]
- Motta, I.O.; Dantas, J.; Vidal, L.; Bílio, J.; Pujol-Luz, J.R.; Albuquerque, É.V.S. The Coffee Leaf Miner, Leucoptera coffeella (Lepidoptera: Lyonetiidae): Identification of the Larval Instars and Description of Male and Female Genitalia. Rev. Bras. Entomol. 2021, 65, e20200122. [Google Scholar] [CrossRef]
- Katiyar, K.P.; Ferrer, F. Rearing Technique, Biology and Sterilization of the Coffee Leaf Miner, Leucoptera coffeella Guer. (Lepidoptera: Lyonetiidae); International Atomic Energy Agency: Vienna, Austria, 1968; pp. 165–175. [Google Scholar]
- Costa, J.N.M.; Teixeira, C.A.D.; Júnior, J.R.V.; Rocha, R.B.; de Freitas Fernandes, C. Informações para facilitar a identificação das diferentes fases do bicho-mineiro (Leucoptera coffeella) em campo. Embrapa Rondônia-Comun. Técnico INFOTECA-E 2012, 4, 1–4. [Google Scholar]
- Wolcott, G.N. A Quintessence of Sensitivity: The Coffee Leaf-Miner. J. Agric. Univ. Puerto Rico 1947, 31, 215–219. [Google Scholar] [CrossRef]
- Guerreiro Filho, O. Coffee Leaf Miner Resistance. Braz. J. Plant Physiol. 2006, 18, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Notley, F.B. The Leucoptera Leaf Miners of Coffee on Kilimanjaro; Leucoptera coffeella, Guér. Bull. Entomol. Res. 1948, 39, 399–416. [Google Scholar] [CrossRef]
- Nielsen, C.D. The Insects of Australia: A Textbook for Students and Research Workers; Melbourne University Press: Carlton, VIC, Australia, 1991; ISBN 978-0-522-84454-2. [Google Scholar]
- de Sabino, P.H.; Junior, F.A.R.; Carvalho, G.A.; Mantovani, J.R. Nitrogen Fertilizers and Occurence of Leucoptera coffeella (Guérin-Mèneville & Perrottet) in Transplanted Coffee Seedlings. Coffee Sci. Braz. 2018, 13, 410–414. [Google Scholar] [CrossRef]
- De Almeida, V.C.; Guimarães, R.J.; Mendes, A.N.G. Infestação por bicho-mineiro e teores foliares de açúcares solúveis totais e proteína em cafeeiros orgânicos. Coffee Sci. 2014, 9, 300–311. [Google Scholar]
- Conceição, C.H.C.; Guerreiro-Filho, O.; Gonçalves, W. Flutuação populacional do bicho-mineiro em cultivares de café arábica resistentes à ferrugem. Bragantia 2005, 64, 625–631. [Google Scholar] [CrossRef] [Green Version]
- Michereff, M.F.F.; Michereff, M.; Vilela, E.F. Mating Behavior of the Coffee Leaf-Miner Leucoptera coffeella (Guerin-Meneville) (Lepidoptera: Lyonetiidae). Neotrop. Entomol. 2007, 36, 376–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Concafe Produção Dos Cafés Do Brasil Atinge 61,62 Milhões de Sacas de 60kg Em 2020, Volume 25% Maior Que 2019. Available online: http://www.consorciopesquisacafe.com.br/index.php/imprensa/noticias/1023-2020-09-29-15-22-05 (accessed on 21 October 2020).
- Shahbandeh, M. Top Coffee Producing Countries. 2018. Available online: https://0-www-statista-com.brum.beds.ac.uk/statistics/277137/world-coffee-production-by-leading-countries/ (accessed on 21 October 2020).
- Cecafé. Relatório Mensal de Exportações—Setembro 2020; Conselho dos Exportadores de Café do Brasil: São Paulo, Brazil, 2020. [Google Scholar]
- Lomelí-Flores, J.R.; Barrera, J.F.; Bernal, J.S. Impacts of Weather, Shade Cover and Elevation on Coffee Leafminer Leucoptera coffeella (Lepidoptera: Lyonetiidae) Population Dynamics and Natural Enemies. Crop Prot. 2010, 29, 1039–1048. [Google Scholar] [CrossRef]
- Bustillo, P.A.E. El minador de la hoja del cafeto, Leucoptera coffeella (Guérin-Méneville) (Lepidoptera: Lyonetiidae). In Los Insectos y su Manejo in Colombian Caficulture; FNC, Cenicafé: Chinchiná, Colombia, 2008; pp. 360–363. [Google Scholar]
- IBGE|Brasil Em Síntese|Território. Available online: https://brasilemsintese.ibge.gov.br/territorio.html (accessed on 4 August 2021).
- Nimer, E.; de Paiva Macedo, A.M. Balanço Hídrico e Clima Da Região Dos Cerrados; IBGE, Departamento de Recursos Naturais e Estudos Ambientais: Rio de Janeiro, Brazil, 1989; ISBN 85-240-0164-X.
- de Oliveira Aparecido, L.E.; de Souza Rolim, G. Models for Simulating the Frequency of Pests and Diseases of Coffea arabica L. Int. J. Biometeorol. 2020, 64, 1063–1084. [Google Scholar] [CrossRef]
- Reis, P.R.; de Souza, J.C.; Venzon, M. Manejo Ecológico Das Principais Pragas Do Cafeeiro. Inf. Agropecuário 2002, 23, 201–215. [Google Scholar]
- Souza, J.C. de Café: Controle do Bicho Mineiro Precisa ser Feito a Partir de Agora para Garantir Grande Safra em. 2018. Available online: /noticias/cafe/187850-cafe-controle-do-bicho-mineiro-precisa-ser-feito-a-partir-de-agora-para-garantir-grande-safra-em-2018.html (accessed on 12 June 2018).
- Jaramillo, J.; Muchugu, E.; Vega, F.E.; Davis, A.; Borgemeister, C.; Chabi-Olaye, A. Some like It Hot: The Influence and Implications of Climate Change on Coffee Berry Borer (Hypothenemus hampei) and Coffee Production in East Africa. PLoS ONE 2011, 6, e24528. [Google Scholar] [CrossRef] [Green Version]
- Weldemichael, G.; Teferi, D. The Impact of Climate Change on Coffee (Coffea Arabica L.) Production and Genetic Resources. Int. J. Res. Stud. Agric. Sci. IJRSAS 2020, 5, 26–34. [Google Scholar] [CrossRef]
- Guedes, R.N.C.; Smagghe, G.; Stark, J.D.; Desneux, N. Pesticide-Induced Stress in Arthropod Pests for Optimized Integrated Pest Management Programs. Annu. Rev. Entomol. 2016, 61, 43–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guedes, R.N.C.; Walse, S.S.; Throne, J.E. Sublethal Exposure, Insecticide Resistance, and Community Stress. Curr. Opin. Insect Sci. 2017, 21, 47–53. [Google Scholar] [CrossRef] [Green Version]
- LORINI, I. Pragas de Graos de Cereais Armazenados; Embrapa Trigo: Passo Fundo, Brazil, 1999. [Google Scholar]
- Vencill, W.K.; Nichols, R.L.; Webster, T.M.; Soteres, J.K.; Mallory-Smith, C.; Burgos, N.R.; Johnson, W.G.; McClelland, M.R. Herbicide Resistance: Toward an Understanding of Resistance Development and the Impact of Herbicide-Resistant Crops. Weed Sci. 2012, 60, 2–30. [Google Scholar] [CrossRef] [Green Version]
- Omoto, C.; Bernardi, O.; Guedes, R.N.C. Classificação do Modo de Ação de Inseticidas. Comitê de Ação à Resistência a Inseticidas. Available online: https://92813ac4-b3b4-47f4-a8b3-43c4292d561c.filesusr.com/ugd/2bed6c_d12a93a1ae8b4c83b17b872407dbfb9a.pdf (accessed on 4 August 2021).
- Omoto, C.; Bernardi, O.; Guedes, R.N.C. Manejo da Resistência do Bicho-Mineiro do Café a Inseticidas. Comitê de Ação à resistência a Inseticidas. Available online: https://92813ac4-b3b4-47f4-a8b3-43c4292d561c.filesusr.com/ugd/2bed6c_0e75e5fd039d4ff1b8d11f9724ea3e7c.pdf (accessed on 4 August 2021).
- Trdan, S. Insecticides Resistance; InTech: Rijeka, Croatia, 2016; pp. 4–13. [Google Scholar]
- Fragoso, D.B.; Guedes, R.N.C.; Guedes, R.N.C.; Picanço, M.C.; Zambolim, L. Insecticide Use and Organophosphate Resistance in the Coffee Leaf Miner Leucoptera coffeella (Lepidoptera: Lyonetiidae). Bull. Entomol. Res. 2002, 92, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Leite, S.A.; Dos Santos, M.P.; Resende-Silva, G.A.; da Costa, D.R.; Moreira, A.A.; Lemos, O.L.; Guedes, R.N.C.; Castellani, M.A. Area-Wide Survey of Chlorantraniliprole Resistance and Control Failure Likelihood of the Neotropical Coffee Leaf Miner Leucoptera coffeella (Lepidoptera: Lyonetiidae). J. Econ. Entomol. 2020, 113, 1399–1410. [Google Scholar] [CrossRef] [PubMed]
- Melo, T.L.; Raetano, C.G.; UNESP; Nery, M.S.; Cardoso, A.D.; Moreira, A.A.; Leite, S.A.; de Jesus, T.F.; Silva, W.G.O.; Castellani, M.A. Management of Coffee Leaf Miner: Spray Volume, Efficacy of Cartap Hydrochloride and Impact on Parasitism. Coffee Sci. 2019, 10, 250–260. [Google Scholar] [CrossRef]
- Diez-Rodríguez, G.I.; de Baptista, G.C.; Trevizan, L.R.P.; Haddad, M.L.; Nava, D.E. Resíduos de tiametoxam, aldicarbe e de seus metabólitos em folhas de cafeeiro e efeito no controle de Leucoptera coffeella (Guérin-Mèneville) (Lepidoptera: Lyonetiidae). Neotrop. Entomol. 2006, 35, 257–263. [Google Scholar] [CrossRef] [Green Version]
- Alves, P.M.P.; de Lima, J.O.G.; de Oliveira, L.M. Monitoramento Da Resistência Do Bicho-Mineiro-Do-Cafeeiro, Perileucoptera coffeella (Guérin-Méneville, 1842) (Lepidoptera: Lyonetiidae), a Inseticidas, Em Minas Gerais. An. Soc. Entomol. Bras. 1992, 21, 77–91. [Google Scholar] [CrossRef]
- Ribeiro, B.M.; de Magalhaes, L.C.; Guedes, R.N.C. Niveis de Resistencia a Inseticidas Organo-Fosforados Em Populacoes de Leucoptera coffeella (Guer-Menev) (Lepidoptera: Lyonetiidae) de Minas Gerais. Biosci. J. 2006, 19, 1–5. [Google Scholar]
- Troczka, B.; Zimmer, C.T.; Elias, J.; Schorn, C.; Bass, C.; Davies, T.G.E.; Field, L.M.; Williamson, M.S.; Slater, R.; Nauen, R. Resistance to Diamide Insecticides in Diamondback Moth, Plutella Xylostella (Lepidoptera: Plutellidae) Is Associated with a Mutation in the Membrane-Spanning Domain of the Ryanodine Receptor. Insect Biochem. Mol. Biol. 2012, 42, 873–880. [Google Scholar] [CrossRef]
- Wang, X.; Wu, Y. High Levels of Resistance to Chlorantraniliprole Evolved in Field Populations of Plutella Xylostella. J. Econ. Entomol. 2012, 105, 1019–1023. [Google Scholar] [CrossRef]
- Roditakis, E.; Vasakis, E.; Grispou, M.; Stavrakaki, M.; Nauen, R.; Gravouil, M.; Bassi, A. First Report of Tuta Absoluta Resistance to Diamide Insecticides. J. Pest Sci. 2015, 88, 9–16. [Google Scholar] [CrossRef]
- Grant, C.; Jacobson, R.; Ilias, A.; Berger, M.; Vasakis, E.; Bielza, P.; Zimmer, C.T.; Williamson, M.S.; ffrench-Constant, R.H.; Vontas, J.; et al. The Evolution of Multiple-Insecticide Resistance in UK Populations of Tomato Leafminer, Tuta Absoluta. Pest Manag. Sci. 2019, 75, 2079–2085. [Google Scholar] [CrossRef]
- Boaventura, D.; Bolzan, A.; Padovez, F.E.; Okuma, D.M.; Omoto, C.; Nauen, R. Detection of a Ryanodine Receptor Target-Site Mutation in Diamide Insecticide Resistant Fall Armyworm, Spodoptera Frugiperda. Pest Manag. Sci. 2020, 76, 47–54. [Google Scholar] [CrossRef]
- Bolzan, A.; Padovez, F.E.; Nascimento, A.R.; Kaiser, I.S.; Lira, E.C.; Amaral, F.S.; Kanno, R.H.; Malaquias, J.B.; Omoto, C. Selection and Characterization of the Inheritance of Resistance of Spodoptera Frugiperda (Lepidoptera: Noctuidae) to Chlorantraniliprole and Cross-Resistance to Other Diamide Insecticides. Pest Manag. Sci. 2019, 75, 2682–2689. [Google Scholar] [CrossRef]
- de Carvalho, C.H.S. Cultivares de Café; Carlos Henrique S. Carvalho: Brasília, Brazil, 2007. [Google Scholar]
- Carvalho, C.H.S. Cultivares de Café: Origem, Características e Recomendações; Embrapa Café: Brasília, Brazil, 2008; p. 247. [Google Scholar]
- Matiello, J.B.; Almeida, S.R.; da Silva, M.B.; Ferreira, I.B.; Carvalho, C.H.S. Siriema VC4, Cultivar Clonal de Cafeeiros com Resistência Múltipla, à Ferrugem e ao Bicho Mineiro; Embrapa Café: Brasília, Brazil, 2015; p. 1. [Google Scholar]
- Matiello, J.B.; de Almeida, S.R.; da Silva, M.B.; Ferreira, I.B.; de Carvalho, C.H.S. Siriema AS1, Cultivar de Cafeeiro com Resistência à Ferrugem e ao Bicho Mineiro; Embrapa Café: Brasília, Brazil, 2015; p. 1. [Google Scholar]
- Guerreiro Filho, O.; Medina Filho, H.P.; Carvalho, A. Fontes de resistência ao bicho-mineiro, Perileucoptera coffeella, em Coffea spp. Bragantia 1991, 50, 45–55. [Google Scholar] [CrossRef]
- Medina, D.M. Microsporogênese em um híbrido tri-plóide de Coffea racemosa lour. x C. arábica L. Bragantia 1963, 22, 299–318. [Google Scholar] [CrossRef]
- Guerreiro Filho, O.; Mazzafera, P. Caffeine Does Not Protect Coffee Against the Leaf Miner Perileucoptera coffeella. J. Chem. Ecol. 2000, 26, 1447–1464. [Google Scholar] [CrossRef]
- Mishra, M.K.; Slater, A. Recent Advances in the Genetic Transformation of Coffee. Biotechnol. Res. Int. 2012, 2012, 580857. [Google Scholar] [CrossRef] [Green Version]
- Villalta-Villalobos, J.; Gatica-Arias, A. A Look Back in Time: Genetic Improvement of Coffee through the Application of Biotechnology. Agron. Mesoam. 2019, 30, 577–599. [Google Scholar] [CrossRef]
- Fernández, R.; De Guglielmo, Z.; Menéndez, A. Cultivo de Tejidos y Transformación Genética de Café Cultivo de Tejidos y Transformación Genética de Café. Rev. Investig. 2010, 34, 57–84. [Google Scholar]
- Leroy, T.; Henry, A.-M.; Royer, M.; Altosaar, I.; Frutos, R.; Duris, D.; Philippe, R. Genetically Modified Coffee Plants Expressing the Bacillus Thuringiensis Cry1Ac Gene for Resistance to Leaf Miner. Plant Cell Rep. 2000, 19, 382–389. [Google Scholar] [CrossRef]
- Barbosa, C.; de Rezende, J.C.; Carvalho, G.R.; de Carvalho, A.M.; Andrade, V.T.; Botelho, C.E. Adaptabilidade e Estabilidade Fenotípica de Cultivares de Café Arábica Em Minas Gerais. Pesqui. Agropecuária Bras. 2010, 45, 1404–1411. [Google Scholar] [CrossRef] [Green Version]
- Perthuis, B.; Pradon, J.; Montagnon, C.; Dufour, M.; Leroy, T. Stable Resistance against the Leaf Miner Leucoptera coffeella Expressed by Genetically Transformed Coffea Canephora in a Pluriannual Field Experiment in French Guiana. Euphytica 2005, 144, 321–329. [Google Scholar] [CrossRef]
- Perthuis, B.; Vassal, J.-M.; Fenouillet, C.; Leroy, T. Cry1Ac Insecticidal Protein Levels in Genetically Modified Coffea Canephora Pierre Coffee Plants Were Negatively Correlated with the Growth Speed in a Field Experiment. Euphytica 2015, 202, 373–383. [Google Scholar] [CrossRef]
- Ran, F.A.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome Engineering Using the CRISPR-Cas9 System. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breitler, J.C.; Dechamp, E.; Campa, C.; Rodrigues, L.A.Z.; Guyot, R.; Marraccini, P.; Etienne, H. CRISPR/Cas9-Mediated Efficient Targeted Mutagenesis Has the Potential to Accelerate the Domestication of #Coffea Canephora#. Plant Cell Tissue Organ Cult. 2018, 134, 383–394. [Google Scholar]
- Amaral, D.S.; Venzon, M.; Pallini, A.; Lima, P.C.; DeSouza, O. Does Vegetational Diversification Reduce Coffee Leaf Miner Leucoptera coffeella (Guérin-Mèneville) (Lepidoptera: Lyonetiidae) Attack? Neotrop. Entomol. 2010, 39, 543–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, F.L.; Mantovani, E.C.; Bonfim Neto, H.; Nunes, V.D.V. Efeitos de Variáveis Ambientais, Irrigação e Vespas Predadoras Sobre Leucoptera coffeella (Guérin-Méneville) (Lepidoptera: Lyonetiidae) No Cafeeiro. Neotrop. Entomol. 2009, 38, 410–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bigger, M. An Investigation by Fourier Analysis into the Interaction between Coffee Leaf-Miners and Their Larval Parasites. J. Anim. Ecol. 1973, 42, 417–434. [Google Scholar] [CrossRef]
- Gallardo-Covas, F. Faunal Survey of the Coffee Leaf Miner, Leucoptera coffeella, Parasitoids in Puerto Rico. J. Agric. Univ. Puerto Rico 1988, 72, 255–263. [Google Scholar] [CrossRef]
- Lomeli-Flores, J.R.; Barrera, J.F.; Bernal, J.S. Impact of Natural Enemies on Coffee Leafminer Leucoptera coffeella (Lepidoptera: Lyonetiidae) Population Dynamics in Chiapas, Mexico. Biol. Control 2009, 51, 51–60. [Google Scholar] [CrossRef]
- Parra, J.R.P.; Gonçalves, W.; Gravena, S.; Marconato, A.R. Parasitos e Predadores Do Bicho-Mineiro Do Cafeeiro Perileucoptera coffeella (Guérin-Méneville, 1842) Em São Paulo. An. Soc. Entomológica Bras. 1977, 6, 138–143. [Google Scholar] [CrossRef]
- Picanço, M.; Oliveira, I.; Fernandes, F.; Prieto, M.; Bacci, L.; Silva, É. Ecology of Vespidae (Hymenoptera)Predators in Coffea Arabica Plantations. Sociobiology 2012, 59, 1269–1280. [Google Scholar]
- Wanjala, F.M.E. Relative Abundance and within Canopy Distribution of the Parasites of the Coffee Leafminer, Leucoptera Meyricki [Lep.: Lyonetiidae] in Kenya. Entomophaga 1978, 23, 57–62. [Google Scholar] [CrossRef]
- Gallardo-Covas, F. Augmentation of Mirax Insularism Muesebeck: Alternative for Population Control of the Coffee Leaf Miner, Leucoptera coffeella Guérin-Ménéville, in Puerto Rico. J. Agric. Univ. Puerto Rico 1992, 76, 43–54. [Google Scholar] [CrossRef]
- Villacorta, A. (Instituto A. do P. Ovicidal Activity of Metarhizium anisopliae Isolate, CM-14 on the Coffee Leaf Miner. Perileucoptera coffeella (Lep.: Lyonetiidae). Entomophaga Fr. 1983, 28, 179–184. [Google Scholar] [CrossRef]
- Draganova, S.A. Tomov Virulence of a Strain Beauveria Bassiana (Bals.) Vuill. (Hyphomycetes, Deuteromycotina) against Larvae of Leucoptera Malifoliella Costa (Lepidoptera, Lyonetiidae). Bulg. J. Agric. Sci. 1998, 4, 435–439. [Google Scholar]
- Iverson, A.; Gonthier, D.; Pak, D.; Ennis, K.; Burnham, R.; Perfecto, I.; Rodriguez, M.; Vandermeer, J. A Multifunctional Approach for Achieving Simultaneous Biodiversity Conservation and Farmer Livelihood in Coffee Agroecosystems. Biol. Conserv. 2019, 238, 108–179. [Google Scholar] [CrossRef]
- Medeiros, H.R.; Grandinete, Y.C.; Manning, P.; Harper, K.A.; Cutler, G.C.; Tyedmers, P.; Righi, C.A.; Ribeiro, M.C. Forest Cover Enhances Natural Enemy Diversity and Biological Control Services in Brazilian Sun Coffee Plantations. Agron. Sustain. Dev. 2019, 39, 50. [Google Scholar] [CrossRef] [Green Version]
- Law, J.H.; Regnier, F.E. Pheromones. Annu. Rev. Biochem. 1971, 40, 533–548. [Google Scholar] [CrossRef] [PubMed]
- Nordlund, D.A.; Lewis, W.J. Terminology of Chemical Releasing Stimuli in Intraspecific and Interspecific Interactions. J. Chem. Ecol. 1976, 2, 211–220. [Google Scholar] [CrossRef]
- Ambrogi, B.; Lima, E.; Sousa-Souto, L. Efficacy of Mating Disruption for Control of the Coffee Leaf Miner. BioAssay 2006, 1, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Francke, W.; Toth, M.; Szöcs, G.; Krieg, W.; Ernst, H.; Buschmann, E. Identifizierung Und Synthese von Dimethylalkanen Als Sexuallockstoffe Weiblicher Miniermotten (Lyonetiidae) / Identification and Synthesis of Dimethylalkanes as Sex Attractants of Female Leaf Miner Moths (Lyonetiidae). Z. Für Nat. C 1988, 43, 787–789. [Google Scholar] [CrossRef]
- Malo, E.A.; Rojas, J.C.; Lopez-Guillen, G.; Barrera, J.F. Chemical Analysis of Female Volatiles and Field Response of the Coffee Leafminer Moth (Lepidoptera: Lyonetiidae) to Stereoisomers of Its Major Sex Pheromone Component. Fla. Entomol. 2009, 92, 548–553. [Google Scholar] [CrossRef]
- Lima, E.R.; Vilela, E.F.; Della Lucia, T.M.C.; Ataíde, L.M.S. Age and Time Related Pheromone Production in Coffee Leafminer Leucoptera coffeella Guérin-Méneville (Lepidoptera: Lyonetiidae). J. Braz. Chem. Soc. 2008, 19, 1659–1662. [Google Scholar] [CrossRef] [Green Version]
- das Graças, M.; Araújo, A.P.A.; de Resende, T.T.; Lima, E. Influence of host plant on reproductive behavior of Leucaoptera coffeella (Guerin-Meneville) (Lepidoptera: Lyonetiidae). Coffee Sci. Braz. 2013, 8, 1–8. [Google Scholar]
- Welter, S.; Pickel, C.; Millar, J.; Cave, F.; Van Steenwyk, R.; Dunley, J. Pheromone Mating Disruption Offers Selective Management Options for Key Pests. Hilgardia 2005, 59, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Lima, E.R. Feromônio Sexual do Bicho-Mineiro do Café, Leucoptera coffeella: Avaliação para uso em Programas de Manejo Integrado; Locus UFV-Repositório Universidade Federal de Viçosa: Viçosa, Brazil, 2001; pp. 20–71. [Google Scholar]
- Cardé, R.T.; Minks, A.K. Control of Moth Pests by Mating Disruption: Successes and Constraints. Annu. Rev. Entomol. 1995, 40, 559–585. [Google Scholar] [CrossRef]
- Ambrogi, B.G. Viabilidade da Técnica de Confusão Sexual de Machos para o Controle do Bicho-Mineiro do Café Leucoptera coffeella (Guérin-Méneville) (Lepidoptera: Lyonetiidae); Locus UFV-Repositório Universidade Federal de Viçosa: Viçosa, Brazil, 2004; pp. 1–29. [Google Scholar]
- Benelli, G.; Lucchi, A.; Thomson, D.; Ioriatti, C. Sex Pheromone Aerosol Devices for Mating Disruption: Challenges for a Brighter Future. Insects 2019, 10, 308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Régnière, J.; Delisle, J.; Dupont, A.; Trudel, R. The Impact of Moth Migration on Apparent Fecundity Overwhelms Mating Disruption as a Method to Manage Spruce Budworm Populations. Forests 2019, 10, 775. [Google Scholar] [CrossRef] [Green Version]
- Isman, M.B. Botanical Insecticides in the Twenty-First Century—Fulfilling Their Promise? Annu. Rev. Entomol. 2020, 65, 233–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, K.W.; Freed, S.; Shoukat, R.F. Efficacy of Entomopathogenic Fungi and Botanicals on Development of Musca Domestica. J. Entomol. Zool. Stud. 2017, 7, 593–599. [Google Scholar]
- FAO. Biopesticides for Locust Control|FAO Stories|Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/fao-stories/article/en/c/1267098/ (accessed on 21 October 2020).
- Gakuubi, M.M.; Wagacha, J.M.; Dossaji, S.F.; Wanzala, W. Chemical Composition and Antibacterial Activity of Essential Oils of Tagetes Minuta (Asteraceae) against Selected Plant Pathogenic Bacteria. Available online: https://www.hindawi.com/journals/ijmicro/2016/7352509/ (accessed on 21 October 2020).
- Isman, M. Commercial Development of Plant Essential Oils and Their Constituents as Active Ingredients in Bioinsecticides. Phytochem. Rev. 2020, 19, 235–241. [Google Scholar] [CrossRef]
- Vidyasagar, G.M.; Tabassum, N. Antifungal Investigations on Plant Essential Oils. A Review. Int. J. Pharm. Pharm. Sci. 2013, 5, 19–28. [Google Scholar]
- Mizubuti, I.; Pereira Pinto, A.; Ribeiro, E.; Fey, R.; Palumbo, G.; Alves, T. Avaliação Da Silagem de Bagaço de Laranja e Silagem de Milho Em Diferentes Períodos de Armazenamento—doi:10.4025/Actascianimsci.V29i4.995. Acta Sci. Anim. Sci. 2007, 29, 371–377. [Google Scholar] [CrossRef]
- Dubey, N.; Shukla, R.; Kumar, A.; Singh, P.; Prakash, B. Prospects of Botanical Pesticides in Sustainable Agriculture. Curr. Sci. 2010, 98, 479–480. [Google Scholar]
- Fanela, T.L.M.; Bentivenha, J.P.F.; Baldin, E.L.L.; Santana, A.S. Assessing Potential Plants Extracts to Reduce Leucoptera coffeella (Lepidoptera: Lyonetiidae) Attack in Coffee. Coffee Sci. 2020, 15, e151653. [Google Scholar] [CrossRef]
- Alves, D.S.; Oliveira, D.F.; Carvalho, G.A.; Carvalho, D.A.; Souza, L.P.; Lasmar, O. Selection of Active Plant Extracts against the Coffee Leaf Miner Leucoptera coffeella (Lepidoptera: Lyonetiidae). Rev. Bras. Plantas Med. 2013, 15, 352–362. [Google Scholar] [CrossRef] [Green Version]
- Alves, D.S.; Oliveira, D.F.; Carvalho, G.A.; dos Santos, H.M.; Carvalho, D.A.; Santos, M.A.I.; de Carvalho, H.W.P. Plant Extracts as an Alternative to Control Leucoptera coffeella (Guerin-Meneville) (Lepidoptera: Lyonetiidae). Neotrop. Entomol. 2011, 40, 123–128. [Google Scholar] [CrossRef] [Green Version]
- Koul, O. Nano-Biopesticides Today and Future Perspectives; Academic Press: Cambridge, MA, USA, 2019; ISBN 978-0-12-815830-2. [Google Scholar]
- Duhan, J.S.; Kumar, R.; Kumar, N.; Kaur, P.; Nehra, K.; Duhan, S. Nanotechnology: The New Perspective in Precision Agriculture. Biotechnol. Rep. Amst. Neth. 2017, 15, 11–23. [Google Scholar] [CrossRef]
- Saranya, K.; Sundaramanickam, A.; Srinath, R.; Thangavelu, R.; Sathishkumar, R.S.; Moorthy, M.; Parthasarathy, S. Green Synthesis of Silver Nanoparticles by Bloom Forming Marine Microalgae Trichodesmium Erythraeum and Its Applications in Antioxidant, Drug-Resistant Bacteria, and Cytotoxicity Activity. J. Saudi Chem. Soc. 2019, 23, 1180–1191. [Google Scholar]
- Krishnamurthy, S.; Veerasamy, M.; Karruppaya, G. A Review on Plant Sources for Nano Biopesticide Production. Lett. Appl. NanoBioSci. 2020, 9, 1348–1358. [Google Scholar] [CrossRef]
- Prasad, R.; Bhattacharyya, A.; Nguyen, Q.D. Nanotechnology in Sustainable Agriculture: Recent Developments, Challenges, and Perspectives. Front. Microbiol. 2017, 8, 124–127. [Google Scholar] [CrossRef] [Green Version]
- Malathi, P.; Nishanthi, P. Green Synthesis and Characterization of Bioinspired Silver, Gold and Platinum Nanoparticles and Evaluation of Their Synergistic Antibacterial Activity after Combining with Different Classes of Antibiotics. Mater. Sci. Eng. C 2019, 96, 693–707. [Google Scholar] [CrossRef]
- Hashem, A.S.; Awadalla, S.S.; Zayed, G.M.; Maggi, F.; Benelli, G. Pimpinella Anisum Essential Oil Nanoemulsions against Tribolium Castaneum-Insecticidal Activity and Mode of Action. Environ. Sci. Pollut. Res. Int. 2018, 25, 18802–18812. [Google Scholar] [CrossRef]
- Adel, M.M.; Salem, N.Y.; Abdel-Aziz, N.F.; Ibrahim, S.S. Application of New Nano Pesticide Geranium Oil Loaded- Solid Lipid Nanoparticles for Control the Black Cutworm Agrotis ipsilon (Hub.) (Lepi., Noctuidae). Eurasian J. Biosci. 2019, 9, 1453–1461. [Google Scholar]
- Wang, Y.; Liu, Y.; Gong, X.; Xie, Z.; Dong, Q.; Dong, M.; Liu, H.; Shao, Q.; Lu, N.; Murugadoss, V.; et al. Amino Graphene Oxide/Dopamine Modified Aramid Fibers: Preparation, Epoxy Nanocomposites and Property Analysis. Polymer 2019, 168, 131–137. [Google Scholar] [CrossRef]
- Castillo, G. O bicho mineiro e os métodos de controle dessa praga tão devastadora. 3rlab 2016. Available online: https://www.3rlab.com.br/2016/06/01/o-bicho-mineiro-e-os-metodos-de-controle-dessa-praga-tao-devastadora/ (accessed on 4 August 2021).
- Barrera, J.F. Beyond IPM: Introduction to the Theory of Holistic Pest Management; Springer: Berlin/Heidelberg, Germany, 2020; ISBN 978-3-030-43370-3. [Google Scholar]
- Andreazi, E.; Sera, G.H.; de Faria, R.T.; Sera, T.; Shigueoka, L.H.; Carvalho, F.G.; Carducci, F.C.; Chamlet, D. Desempenho de Híbridos F1 de café Arábica com Resistência Simultânea a Ferrugem, Mancha Aureolada e Bicho Mineiro; Coffee Science: Lavras, Brazil, 2015; pp. 375–382. [Google Scholar]
- Dort, E.N.; Tanguay, P.; Hamelin, R.C. CRISPR/Cas9 Gene Editing: An Unexplored Frontier for Forest Pathology. Front. Plant Sci. 2020, 11, 1126. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dantas, J.; Motta, I.O.; Vidal, L.A.; Nascimento, E.F.M.B.; Bilio, J.; Pupe, J.M.; Veiga, A.; Carvalho, C.; Lopes, R.B.; Rocha, T.L.; et al. A Comprehensive Review of the Coffee Leaf Miner Leucoptera coffeella (Lepidoptera: Lyonetiidae)—A Major Pest for the Coffee Crop in Brazil and Others Neotropical Countries. Insects 2021, 12, 1130. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121130
Dantas J, Motta IO, Vidal LA, Nascimento EFMB, Bilio J, Pupe JM, Veiga A, Carvalho C, Lopes RB, Rocha TL, et al. A Comprehensive Review of the Coffee Leaf Miner Leucoptera coffeella (Lepidoptera: Lyonetiidae)—A Major Pest for the Coffee Crop in Brazil and Others Neotropical Countries. Insects. 2021; 12(12):1130. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121130
Chicago/Turabian StyleDantas, Juliana, Isabela O. Motta, Leonardo A. Vidal, Eliza F. M. B. Nascimento, João Bilio, Júlia M. Pupe, Adriano Veiga, Carlos Carvalho, Rogerio B. Lopes, Thales L. Rocha, and et al. 2021. "A Comprehensive Review of the Coffee Leaf Miner Leucoptera coffeella (Lepidoptera: Lyonetiidae)—A Major Pest for the Coffee Crop in Brazil and Others Neotropical Countries" Insects 12, no. 12: 1130. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121130