
Functional Response and Control Potential of Orius sauteri (Hemiptera: Anthocoridae) on Tea Thrips (Dendrothrips minowai Priesner)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Collection
2.2. Method
2.2.1. O. sauteri Nymphs and Adults Preying on the Tea Thrips
2.2.2. O. sauteri Adults Preying on Tea Thrips Adults at Different Temperatures
2.2.3. Intra-Specific Interference Response of Predators
2.2.4. Response of O. sauteri to Population Density of Tea Thrips
2.3. Data Analysis
2.3.1. Predatory Function
2.3.2. O. sauteri Disturbance Response
3. Results
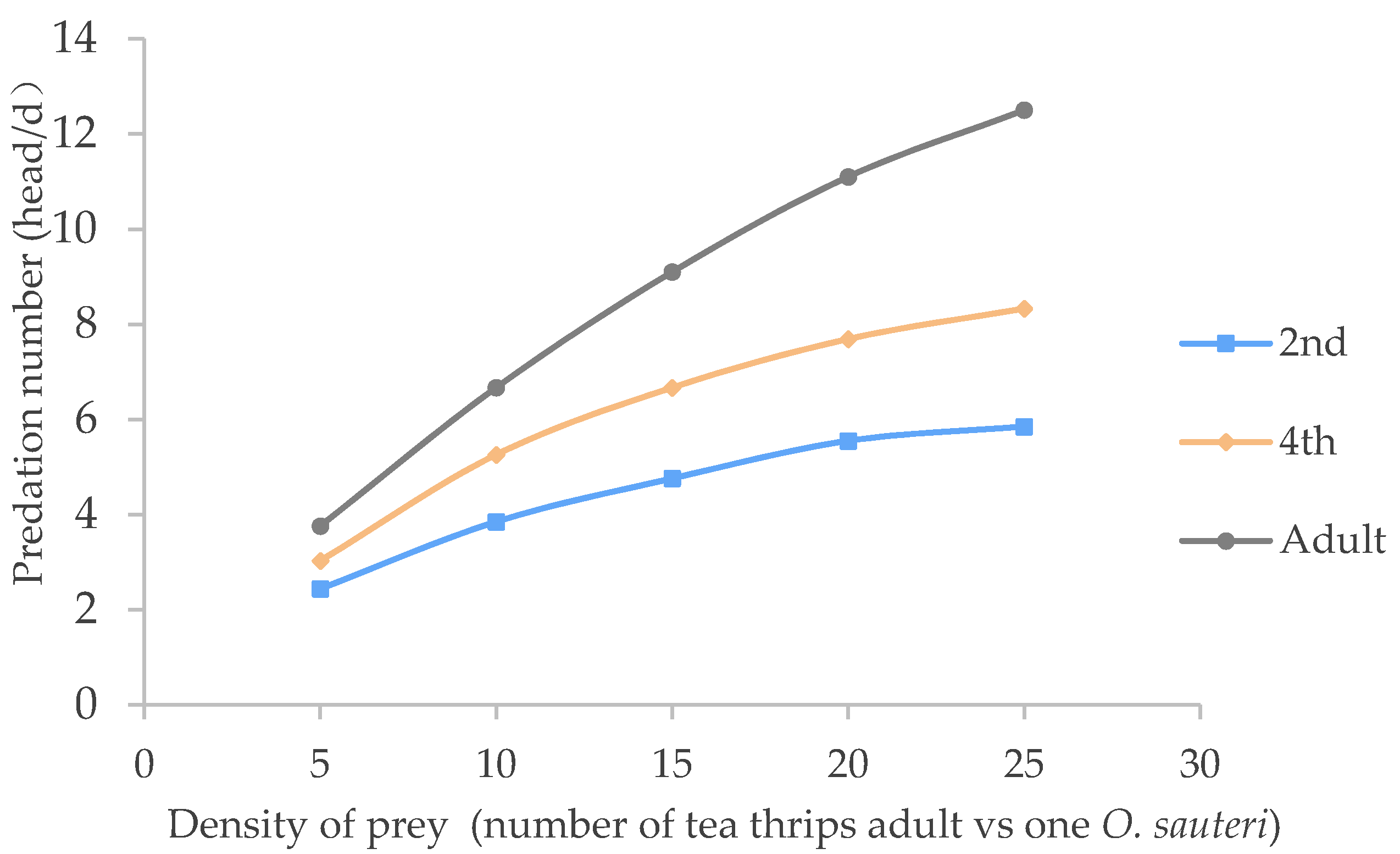
3.1. Predatory Effects of O. sauteri on the Adult Tea Thrips
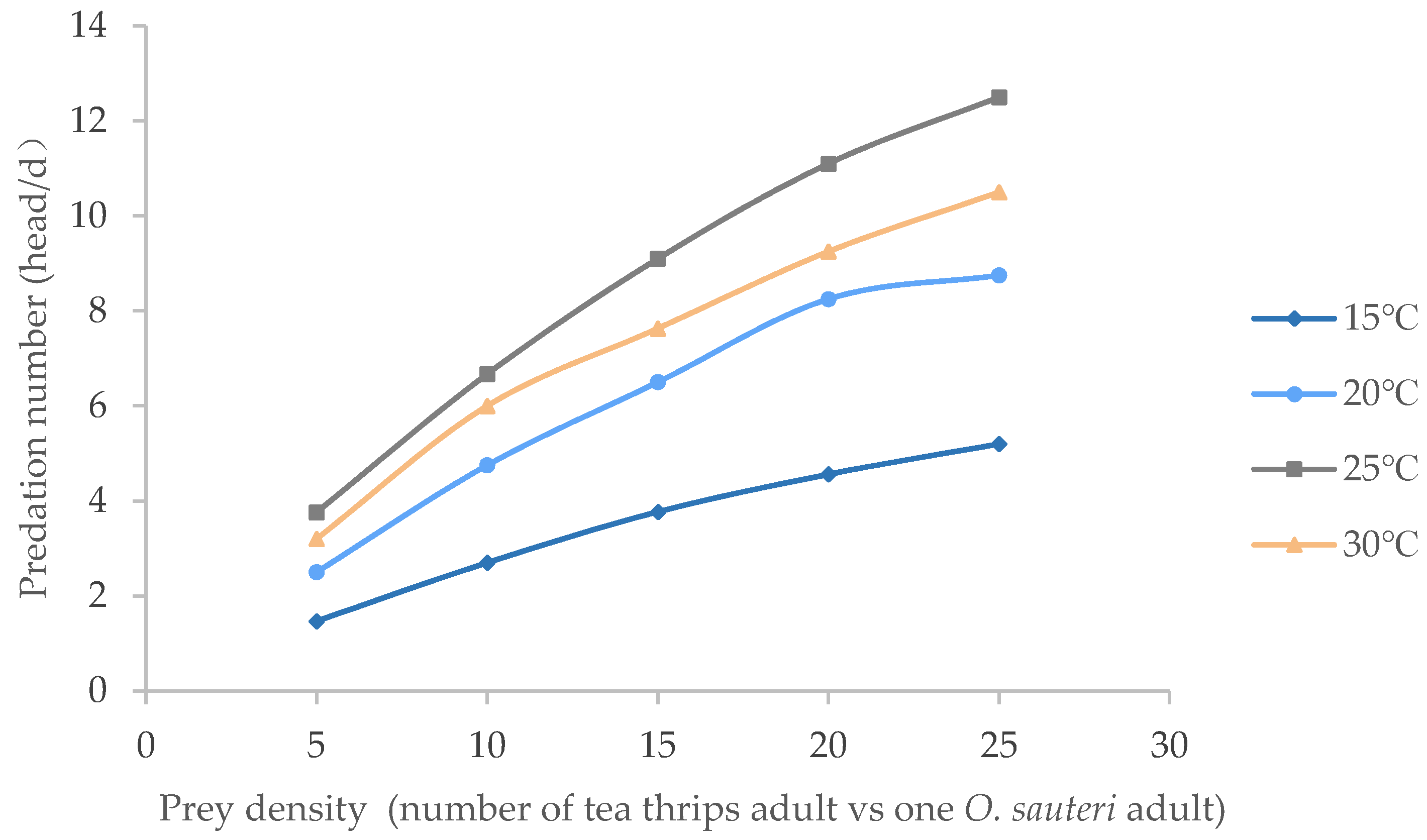
3.2. Temperature Affected Predation Ability of O. sauteri Adults
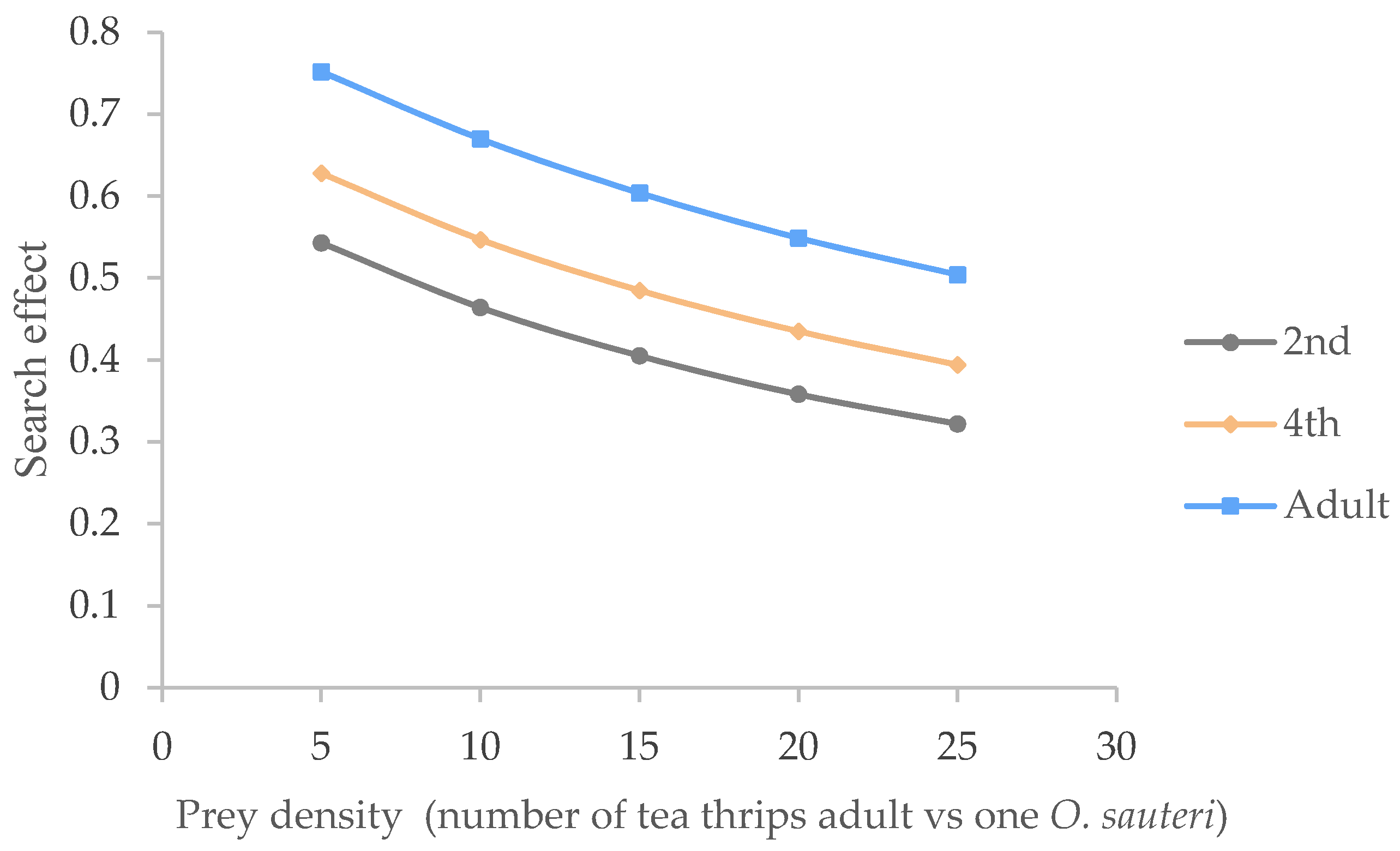
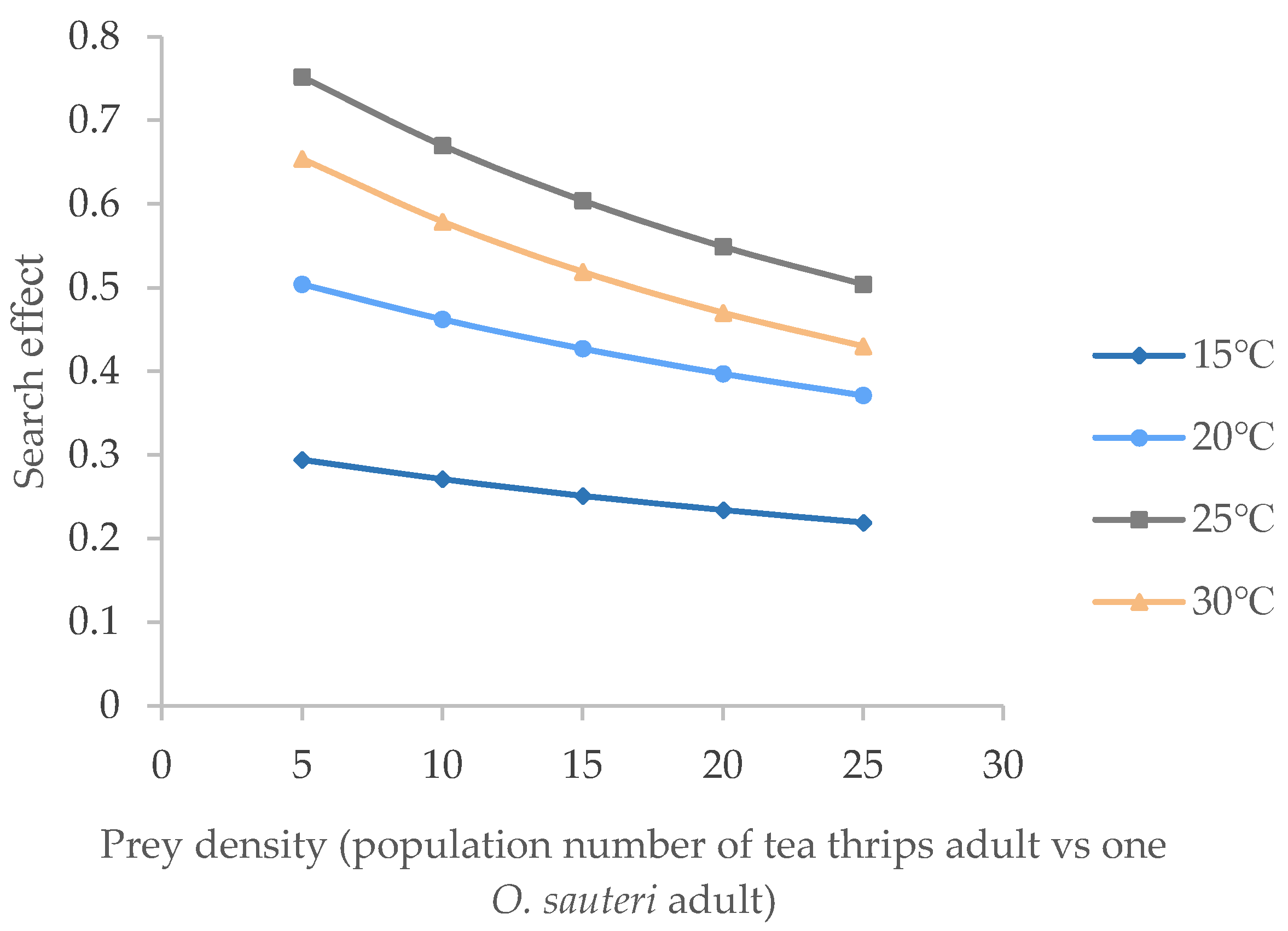
3.3. Searching Effects of O. sauteri for the of Adult Tea Thrips
3.4. Intra-Specific Interference Response of O. sauteri
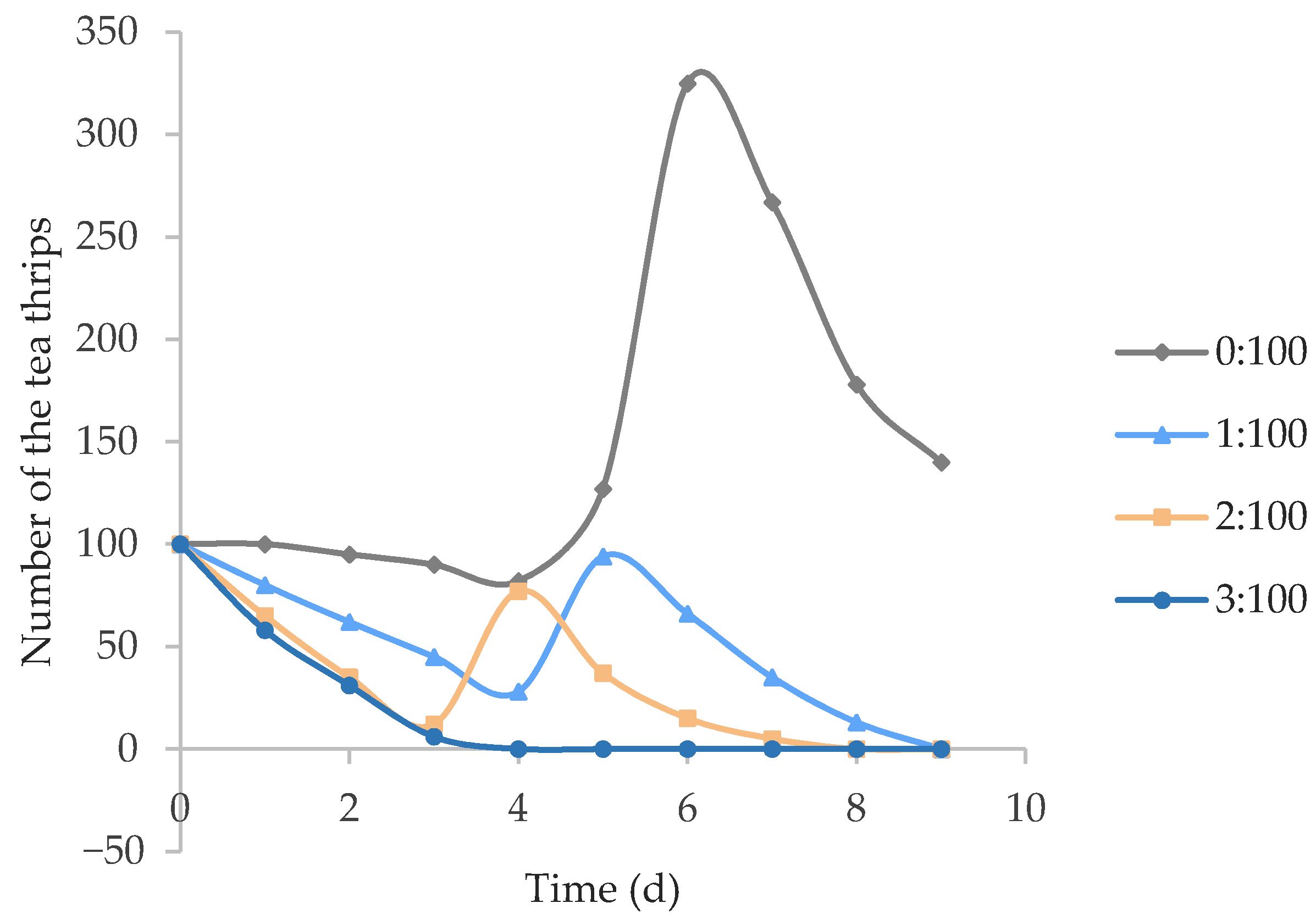
3.5. The Effects of O. sauteri on Population Density of Tea Thrips
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Singh, H.R.; Hazarika, P. Biotechnological approaches for tea improvement. In Biotechnological Progress and Beverage Consumption; Academic Press: Cambridge, MA, USA, 2020; pp. 111–148. [Google Scholar]
- Jones, D.R. Plant viruses transmitted by thrips. Eur. J. Plant Pathol. 2005, 113, 119–157. [Google Scholar] [CrossRef]
- Settele, J.; Biesmeijer, J.; Bommarco, R. Switch to ecological engineering would aid independence. Nature 2008, 456, 570. [Google Scholar] [CrossRef] [Green Version]
- Gurr, G.M.; Wratten, S.D.; Snyder, W.E.; Read, D.M.Y. Biodiversity and Insect Pests: Key Issues for Sustainable Management; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Chailleux, A.; Mohl, E.K.; Alves, M.T.; Messelink, G.J.; Desneux, N. Natural enemy-mediated indirect interactions among prey species: Potential for enhancing biocontrol services in agroecosystems. Pest Manag. Sci. 2014, 70, 1769–1779. [Google Scholar] [CrossRef]
- Li, H.L.; Li, P.; Zhang, H.; Wang, D.F.; Li, L.D.; Zeng, M.S.; Wu, G.Y.; Wang, Q.S. Predation of Orius sauterion pest insects of pea pushes. Acta Teas Sin. 2019, 60, 38–40. [Google Scholar]
- Nagai, K.; Yano, E. Predation by Orius sauteri (Poppius) (Heteroptera: Anthocoridae) on Thrips palmi Karny (Thysanoptera: Thripidae). Functional response and selective predation. Appl. Èntomol. Zool. 2000, 35, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Yin, J.; Gao, X.G.; Wu, Y.Q. Thrips control on the greenhouse eggplant by releasing Orius sauteri (Heteroptera: Anthocoridae). Chin. J. Biol. Control 2013, 29, 459–462. [Google Scholar]
- Guo, J.Y.; Wan, F.H. Use Kalanchoe bolssfeldiana as oviposition plant for mass rearing Orius sauteri (Hemiptera: Anthocoridae). Chin. J. Biol. Control 2001, 17, 53–56. [Google Scholar]
- Zhang, W. A brief description of the species and biology of common Orius insidiosus (Hemiptera: Anthocoridae). Nat. Enemies Insects 1980, 2, 20–27. [Google Scholar]
- Wenjun, B.; Leyi, Z. Chinese Animal Records: Insects; Science Press: Beijing, China, 2011; pp. 185–187. [Google Scholar]
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. Habitat management to conserve natural enemies of arthropod pests in agriculture. Annul. Rev. Entomol. 2000, 45, 175–201. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Ji, D.; Zhang, Q.; Jin, L. Evaluation of eleven plant species as potential banker plants to support predatory Orius sauteri in tea plant systems. Insects 2021, 12, 162. [Google Scholar] [CrossRef] [PubMed]
- Ha, Q.H. Study on morphological, biological, ecological characteristics of predacious bug Orius Poppius (Hemiptera: Anthocoridae) reared on Thrips palmi Karny and eggs of Corcyra cephalonica Staintou. Proc. Vietnam. Natl. Conf. Entomol. 2002, 4, 210–214. [Google Scholar]
- Nagai, K.; Yano, E. Effects of temperature on the development and reproduction of Orius sauteri (Poppius) (Heteroptera: Anthocoridae), a predator of Thrips palmi Karny (Thysanoptera: Thripidae). Appl. Entomol. Zool. 1999, 34, 223–229. [Google Scholar] [CrossRef] [Green Version]
- Holling, C.S. Some characteristics of simple types of predation and parasitism. Can. Ent. 1959, 91, 385–398. [Google Scholar] [CrossRef]
- Wu, K.J.; Sheng, C.F.; Gong, P.Y. Equation of predator functional response and estimation of the parameters in it. Entomol. Knowl. 2004, 41, 267–269. [Google Scholar]
- Ding, Y.Q. Mathematical Ecology of Insects; Science Press: Beijing, China, 1994; pp. 303–304. [Google Scholar]
- Hassell, M.P.; Verley, G.C. New inductive population model for insect parasite and its bearing on biological control. Nature 1969, 223, 1113–1117. [Google Scholar] [CrossRef]
- Schenk, D.; Bacher, S. Functional response of a generalist insect predator to one of its prey species in the field. J. Anim. Ecol. 2010, 71, 524–531. [Google Scholar] [CrossRef]
- Bing, L.; Meng, S.; Yifan, Z.; Hao, C.; Yi, Y.; Li, Z. Evaluation of the biocontrol capacity of predatory bug Orius sauteri, reared on Sitotroga cerealella eggs, on Thrips palmi based on predatory functional response. J. Plant Prot. 2017, 44, 875–876. [Google Scholar]
- Bing, L.V.; Sun, M.; Zhai, Y.F.; Chen, H.; Yi, Y.U.; Zheng, L. Effection of short adaptive pre-feeding on the predatory functional response to Orius sauteri reared on Sitotroga cerealella eggs. J. Environ. Entomol. 2018, 40, 64–69. [Google Scholar]
- Fu, B.L.; Qiu, H.Y.; Li, Q.; Sun, Y.T.; Zhou, S.H.; Yang, S.Y.; Shan-guang, L.; Tang, L.D.; Zhang, F.P.; Liu, K. Predation of Orius sauteri on Thrips hawaiiensis in the laboratory. Chin. J. Appl. Entomol. 2019, 56, 91–98. [Google Scholar]
- Han, L.L.; Dong, T.Y.; Zhao, K.J.; Zhu, M.H.; Sun, W.P.; Xu, Z.X.; Shi, L. Predation of Aphis glycines by Orius sauteri nymphs. China J. Biol. Control. 2015, 31, 323–326. [Google Scholar]
- Liang, Z.; Ge, Z.T.; Gong, Y.J.; Shi, B.C.; Su, W.; Wei, S.J. Effects of temperature on predation of the thrips Echinothrips americanus (Thysanoptera: Thripidae) by the predatory bug Orius sauteri (Heteroptera: Anthocoridae). J. Plant Prot. 2015, 42, 229–236. [Google Scholar]
- Zhi, J.R. The predation of Orius similis to Frankliniella occidentalis and Aphis craccivora. Chin. J. Appl. Entomol. 2011, 48, 573–578. [Google Scholar]
- Shang, S.Q.; Liu, P.; Chen, Y.N.; Zhang, X.H. Functional response and control potential of Neoseiulus barkeri to Tetranychus urticae. Plant Prot. 2017, 43, 118–121. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| O. sauteri | Functional Response Equation | R | Handling Time (Th)/Day | Instantaneous Attack Rate (a) | Predatory Capacity (a/Th) | Predation Upper Limit (No./Day) |
|---|---|---|---|---|---|---|
| 2nd nymph | 1/Na = 1.5262/N + 0.1051 | 0.9429 | 0.1051 | 0.6552 | 6.420 | 9.500 |
| 4th nymph | 1/Na = 1.356/N + 0.0588 | 0.9757 | 0.05882 | 0.7373 | 12.53 | 17.00 |
| Adult | 1/Na = 1.1664/N + 0.0327 | 0.9982 | 0.03270 | 0.8573 | 26.21 | 30.50 |
| Temp. (°C) | Functional Response Equation | R | Th (d) | a | a/Th | Predation Upper Limit (No./d) |
|---|---|---|---|---|---|---|
| 15 | 1/Na = 3.0948/N + 0.059 | 0.9968 | 0.0590 | 0.3231 | 5.480 | 17.00 |
| 20 | 1/Na = 1.809/N + 0.0354 | 0.9814 | 0.03540 | 0.5528 | 15.61 | 28.00 |
| 25 | 1/Na = 1.1664/N + 0.0327 | 0.9982 | 0.03210 | 0.8573 | 26.21 | 30.50 |
| 30 | 1/Na = 1.3603/N + 0.04 | 0.9833 | 0.04000 | 0.7531 | 18.82 | 25.00 |
| Insect Stage | No. of O. sauteri VS 30 Tea Thrips (X) | No. of Total Predation (A) | Predation Rate per O. sauteri | Intra-Specific Interference Response Equation | R2 |
|---|---|---|---|---|---|
| 2nd nymph | 1 | 7.3 | 24.3% | A = 7.9026X−0.933 | 0.9335 |
| 2 | 9 | 15% | |||
| 3 | 9.9 | 11% | |||
| 4 | 7.2 | 6% | |||
| 4th nymph | 1 | 15.3 | 51% | A = 15.953X−0.629 | 0.9783 |
| 2 | 22.4 | 37.35 | |||
| 3 | 24 | 26.7% | |||
| 4 | 25.6 | 21.3% | |||
| Adult | 1 | 18.7 | 62.3% | A = 19.257X−0.523 | 0.9887 |
| 2 | 25 | 41.7% | |||
| 3 | 25.8 | 28.7% | |||
| 4 | 27.6 | 23% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Q.; Zhang, R.; Zhang, Q.; Ji, D.; Zhou, X.; Jin, L. Functional Response and Control Potential of Orius sauteri (Hemiptera: Anthocoridae) on Tea Thrips (Dendrothrips minowai Priesner). Insects 2021, 12, 1132. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121132
Zhang Q, Zhang R, Zhang Q, Ji D, Zhou X, Jin L. Functional Response and Control Potential of Orius sauteri (Hemiptera: Anthocoridae) on Tea Thrips (Dendrothrips minowai Priesner). Insects. 2021; 12(12):1132. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121132
Chicago/Turabian StyleZhang, Qiuping, Ruifang Zhang, Qiuqiu Zhang, Dezhong Ji, Xia Zhou, and Linhong Jin. 2021. "Functional Response and Control Potential of Orius sauteri (Hemiptera: Anthocoridae) on Tea Thrips (Dendrothrips minowai Priesner)" Insects 12, no. 12: 1132. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121132