
Functional Response and Predation Potential of Carabus elysii Adults against the Terrestrial Slug Agriolimax agrestis


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Slugs and Beetles
2.2. Functional Response
2.3. Searching Efficiency
2.4. Intraspecific Competition
2.5. Data Analysis
3. Results and Analysis
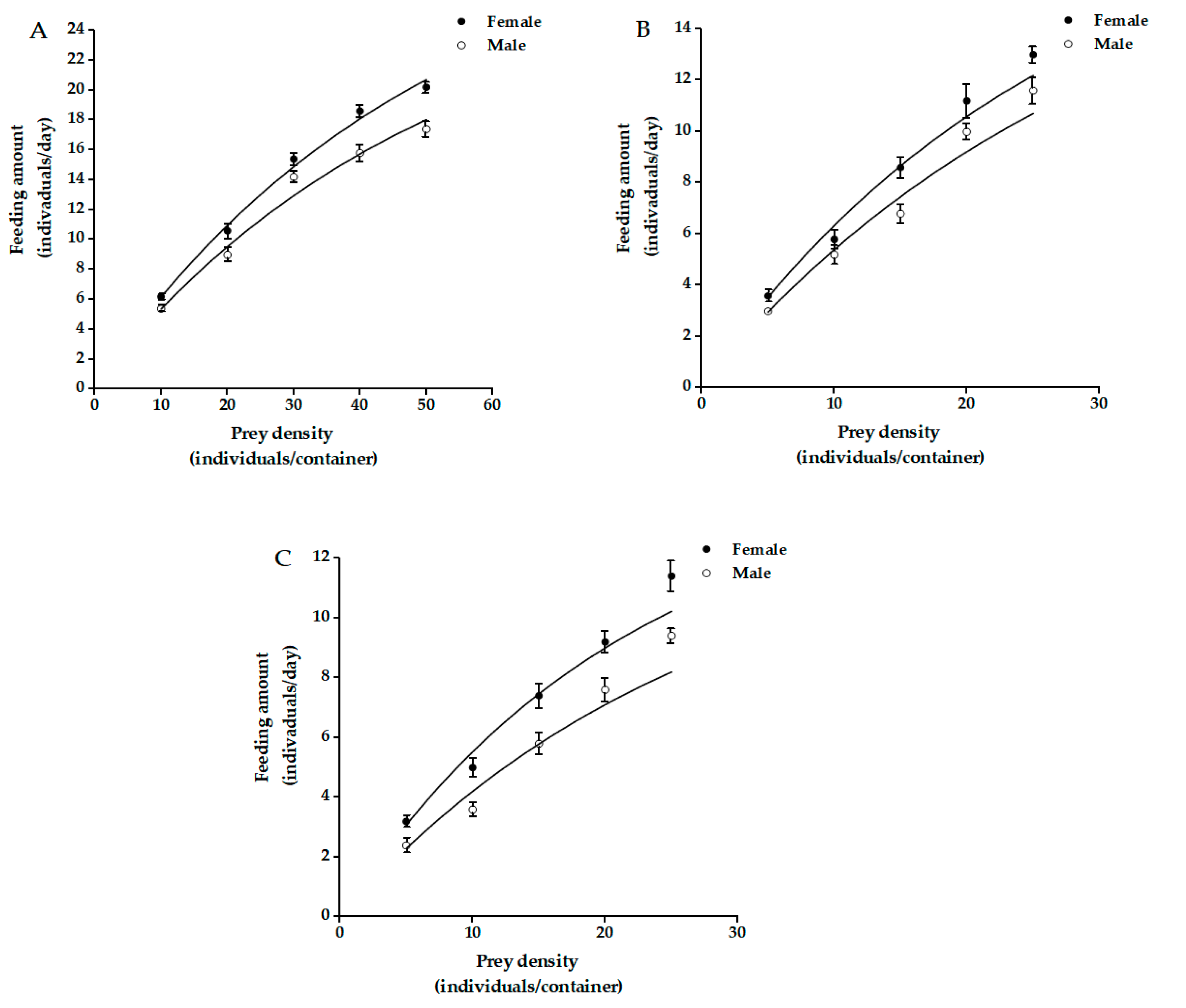
3.1. Functional Response
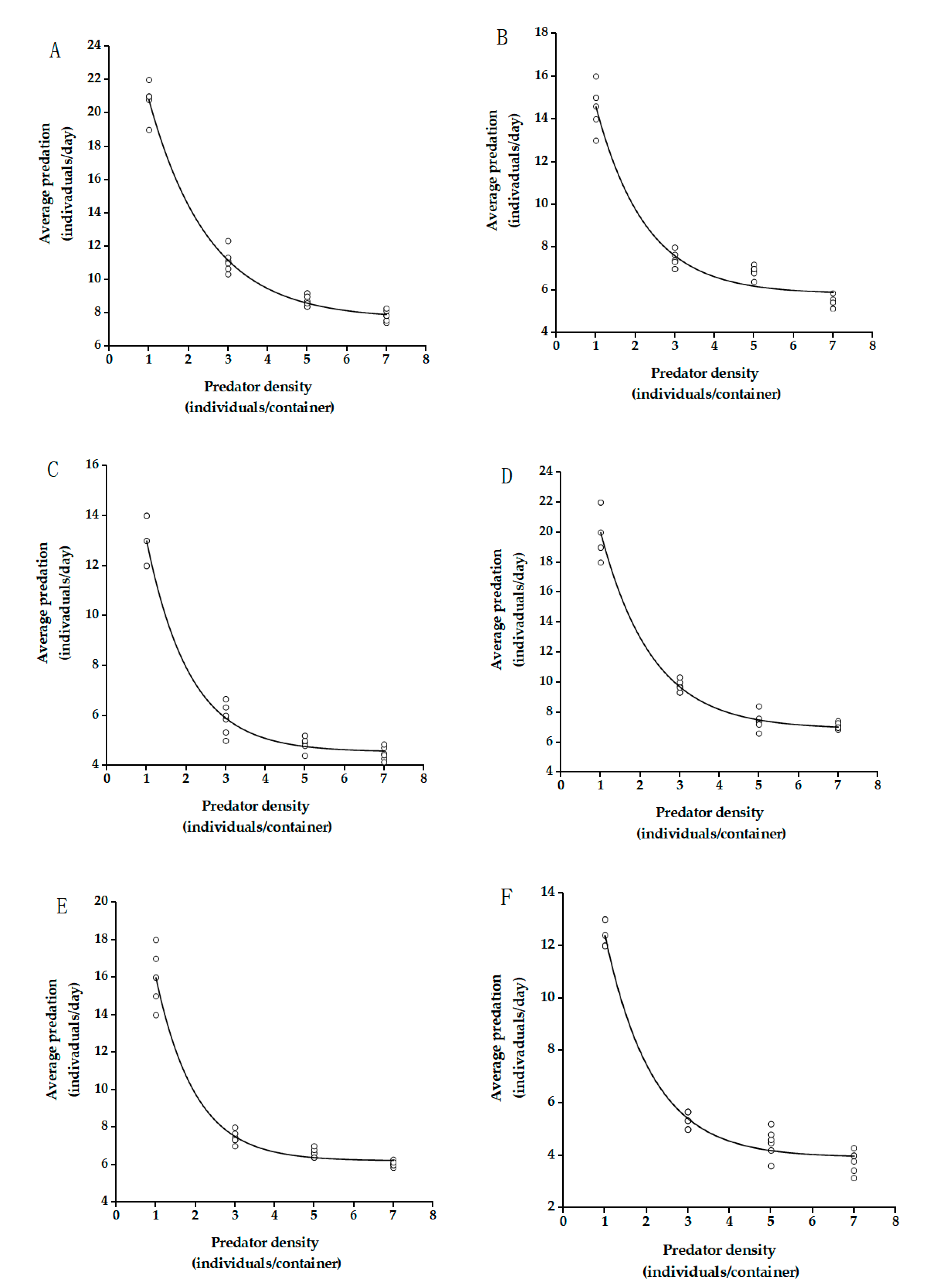
3.2. Searching Efficiency
3.3. Intraspecific Competition
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A. Photos of Slug Damage and Beetle Predation

References
- South, A. Terrestrial Slugs: Biology, Ecology and Control; Chapman and Hall: London, UK, 1992. [Google Scholar]
- Barker, G.M. Molluscs as Crop Pests; CABI Publishing: Wallingford, UK, 2002; 468p. [Google Scholar]
- Liu, Y.Y.; Chen, D.N. The morphology of slugs and their damage to agriculture. Bull. Biol. 1966, 1, 23–27. [Google Scholar]
- Chen, D.N.; Gao, J.X. Common snails and slugs that damage crops and their control. Chin. J. Zool. 1976, 3, 24–26. [Google Scholar]
- Xue, D.J.; Deng, Y.; Luo, G.W.; Xia, X.Y.; Tian, H. Regularity of outbreak and control of Agriolimax cagrestis in Guiyang region. Guizhou Agric. Sci. 1998, 2, 40–43. [Google Scholar]
- Li, Y.H. Damage characteristics and control methods of Agriolimax agrestis, a vegetable crop in wengan. China Plant Prot. 2005, 8, 21. [Google Scholar]
- Li, Z.Z. List of Wildlife in Guizhou; Guizhou Science and Technology Publishing House: Guiyang, China, 2011. [Google Scholar]
- Chen, D.N.; Gao, J.X. Molluscs from Agricultural Regions of China; Agriculture Publishing House: Beijing, China, 1984. [Google Scholar]
- Chen, D.N.; Gao, J.X.; Shi, J.C. Natural extinction of Agriolimax agrestis in relation to agrometeorological conditions. Plant Prot. 1981, 4, 39. [Google Scholar]
- Shi, D.M.; Jiao, J.H. Study on the occurring regularity and integrated control of Agriolimax agrestis in green-house. For. Pest Dis. 1998, 1, 3–4, 27. [Google Scholar]
- Ne, Q.S.; Liu, Y.Q.; Li, C.; Huang, N.Y. Occurrence and control of wild slugs on farm tobacco. Sichuan Agric. Sci. Technol. 2009, 3, 49. [Google Scholar]
- Zhou, Y.S.; Yang, M.; Zhu, P.; Zhou, Z.Q.; Su, P.; Deng, J.; Wang, T.D. Occurrence pattern of Agriolimax agrestis in tobacco fields and its integrated control. J. Southwest Agric. Univ. 2003, 2, 93–100+110. [Google Scholar]
- Yu, Y.W. The research on Agriolimax agrestis breeding habits. Liaoning Agric. Sci. 2015, 2, 66–69. [Google Scholar]
- Guan, W.G. Patterns of occurrence and chemical control of Agriolimax agrestis in greenhouses in Daqing. Chin. J. Zool. 1989, 5, 6–9. [Google Scholar]
- Liu, H.L.; Liu, X. The occurrence and control of Agriolimax agrestis in tobacco fields in Sichuan. Sichuan Agric. Sci. Technol. 2013, 9, 39. [Google Scholar]
- Zhu, F.C. Patterns of occurrence of edible mushroom slugs and comprehensive prevention and control measures. J. Chang. Veg. 2017, 23, 54–56. [Google Scholar]
- Ni, P.; Zhang, J.Q.; Li, W.; Li, N.; Liu, Y.G.; Ma, C.W. Screening and efficacy trials of molluscicide for the control of snail pests in the field. Yunnan Agric. Sci. Technol. 2018, s1, 90–92. [Google Scholar]
- DeSilva, S.M.; Chesmore, D.; Smith, J.; Port, G. Listening to Slugs: Acceptability and Consumption of Molluscicide Pellets by the Grey Field Slug, Deroceras reticulatum. Insects 2021, 12, 548. [Google Scholar] [CrossRef] [PubMed]
- Amy, C.; Neil, A.; Gordon, P. The fate of Deroceras reticulatum following metaldehyde poisoning. Insects 2021, 12, 344. [Google Scholar]
- Barratt, B.I.P.; Moran, V.C.; Bigler, F.; van Lenteren, J.C. The status of biological control and recommendations for improving uptake for the future. BioControl 2018, 63, 155–167. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.Z.; Guo, Q.; Yang, Y.Y.; Chen, W.L. Study on the predatory function of larva of Lampyrigera yunnana (Fairmaire) Coleoptera: Cantharidae. J. Mt. Agric. Biol. 2016, 35, 25–29. [Google Scholar]
- Archita, B.; Christopher, D.W.; Jenna, L.R. A literature review of biological and bio-rational control strategies for slugs: Current research and future prospects. Insects 2021, 12, 541. [Google Scholar]
- Kromp, B. Carabid beetles in sustainable agriculture: A review on pest control efficacy, cultivation impacts and enhancement. Agric. Ecosyst. Environ. 1999, 74, 187–228. [Google Scholar] [CrossRef]
- Symondson, W.O.C.; Sunderland, K.D.; Greenstone, M.H. Can generalist predators be effective biocontrol agents? Annu. Rev. Entomol. 2002, 47, 561–594. [Google Scholar] [CrossRef] [Green Version]
- Symondson, W.O.C. Coleoptera (Carabidae, Staphylinidae, Lampyridae, Drilidae and Silphidae) as predators of terrestrial gas-tropods. In Natural Enemies of Terrestrial Molluscs; Barker, G.M., Ed.; CABI Publishing: Wallingford, UK, 2004; pp. 37–84. [Google Scholar]
- Hu, D.J. Ningbo Forestry Pest Atlas; China Science and Technology Publishing House: Beijing, China, 2013. [Google Scholar]
- Shi, J. Common Forest Insects and Spiders in the Middle and Lower Reaches of the Yangtze River; Northeast Forestry University Publishing House: Harbin, China, 2008. [Google Scholar]
- Zhao, M.J.; Li, L.Z.; Tang, L. The Colourful World of Insects, An Ecological Catalogue of 600 Species of Insects in China; Shanghai Popular Science Publishing House: Shanghai, China, 2005. [Google Scholar]
- Zhu, C.Q.; Qing, D.M.; Yin, X.M. Insect Fauna of Henan: Coleoptera; Henan Science and Technology Press: Zhengzhou, China, 1999. [Google Scholar]
- Yang, X.R.; Li, W.B. Comparative morphology of the mouthparts of three species of Carabus insects. Mod. Agric. Sci. Technol. 2006, 12, 125–126. [Google Scholar]
- Qi, Q. Ultrastructure on the Cephalic Appeendages sensorial of three kinds of carabus. Master Thesis, Anhui University, Hefei, China, 2015. [Google Scholar]
- Li, W.B.; Liu, N.Y.; Xu, Q.; Sun, M.; Fang, J.; Wang, S.Y. Ultrastructure structure of antennal sensilla of carabid beetle Carabus elysii Thomson, 1856 (Coleoptera: Carabidae). Entomol. Res. 2017, 49, 81–86. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.J.; Wang, R.; Liu, B.; Huang, B.; Hou, Y.M.; Tang, B.Z. Molluscicidal activity of tea saponin to Pomacea canaliculata and its safety evaluation against three aquatic organisms. Chin. J. Pestic. Sci. 2021, 23, 139–145. [Google Scholar]
- Holling, C.S. Some characteristics of simple type of predation and parasitism. Can. Entomol. 1959, 91, 385–398. [Google Scholar] [CrossRef]
- Ding, Y.Q. Principles and Applications of Mathematical Ecology of Insect Populations; Science Press: Beijing, China, 1980; Volume 11, pp. 195–211. [Google Scholar]
- Hassell, M.P.; Varley, G.C. New inductive population model for insect parasites and its bearing on biological control. Nature 1969, 223, 1133–1137. [Google Scholar] [CrossRef] [PubMed]
- Fu, B.L.; Qiu, H.Y.; Li, Q.; Shun, Y.Y.; Zhou, S.H.; Yang, S.Y.; Li, S.G.; Tang, L.G.; Zhang, F.P.; Liu, K. Predation of Orius sauteri on Thrips hawaiiensis in the laboratory. Chin. J. Appl. Entomol. 2019, 56, 91–98. [Google Scholar]
- Oberholzer, F.; Frank, T. Predation by the Carabid Beetles Pterostichus melanarius and Poecilus cupreus on Slugs and Slug Eggs. Biocontrol. Sci. Technol. 2003, 1, 99–110. [Google Scholar] [CrossRef]
- McKemey, A.R.; Symondson, W.O.C.; Glen, D.M. Predation and prey size choice by the carabid beetle Pterostichus melanarius (Coleoptera: Carabidae): The dangers of extrapolating from laboratory to field. Bull. Entomol. Res. 2003, 93, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Pianezzola, E.; Roth, S.; Hatteland, B.A. Predation by carabid beetles on the invasive slug Arion vulgaris in an agricultural semi-field experiment. Bull. Entomol. Res. 2013, 103, 225–232. [Google Scholar] [CrossRef]
- Wendland, S.; Baudy, P.; Fusser, M.S.; Entling, M.H.; Schirmel, J. Contrasting predation pressure of different ground beetles on slugs and their eggs. BioControl 2017, 62, 749–756. [Google Scholar] [CrossRef]
- Zhou, J.Z.; Chen, C.M. Predation of wolf spider, Lycsoa Pseudoamulata on brown planthopper, Nilaparvata lugens, and its simalution model: I.functional response. Biological. Control. Bull. 1986, 2, 2–9. [Google Scholar]
- El-Danasoury, H.; Cerecedo, C.; Córdoba, M.; Iglesias-Piñeiro, J. Predation by the carabid beetle Harpalus rufipes on the pest slug Deroceras reticulatum in the laboratory. Ann. Appl. Biol. 2017, 170, 251–262. [Google Scholar] [CrossRef]
- El-Danasoury, H.; Iglesias-Piñeiro, J. Predation by polyphagous carabid beetles on eggs of a pest slug: Potential implications of climate change. J. Appl. Entomol. 2018, 142, 340–348. [Google Scholar] [CrossRef]
- Mckemey, A.R.; Symondson, W.O.C.; Glen, D.M.; Brain, P. Effects of slug size on predation by Pterostichus melanarius (Coleoptera: Carabidae). Biocontrol. Sci. 2001, 11, 83–93. [Google Scholar] [CrossRef]
- Paill, W. Slug feeding in the carabid beetle Pterostichus melanarius: Seasonality and dependence on prey size. J. Molluscan Stud. 2004, 70, 203–205. [Google Scholar] [CrossRef] [Green Version]
- Hatteland, B.A.; Grutle, K.; Mong, C.E.; Skartveit, J.; Symondson, W.O.C.; Solhøy, T. Predation by beetles (Carabidae, Staphylinidae) on eggs and juveniles of the Iberian slug Arion lusitanicus in the laboratory. Bull. Entomol. Res. 2010, 100, 559–567. [Google Scholar] [CrossRef]
- O’Hanlon, A.; Williams, C.D.; Gormally, M.J. Terrestrial slugs (Mollusca: Gastropoda) share common anti-predator defence mechanisms but their expression differs among species. J. Zool. 2008, 307, 203–214. [Google Scholar] [CrossRef] [Green Version]
- Bursztyka, P.; Lafont-Lecuelle, C.; Teruel, E.; Leclercq, J.; Brin, A.; Pageat, P. Behavioural response of xerophilous land snail and slug species to chemical cues from ground-beetle predators: The role of ecological relevance. J. Molluscan Stud. 2018, 84, 141–147. [Google Scholar] [CrossRef]
- Fontan, P. Influence of slug defence mechanisms on the prey preferences of the carabid predator Pterostichus melanarius (Coleptera: Carabidae). Eur. J. Entomol. 2004, 101, 359–364. [Google Scholar]
- Symondson, W.O.C. Biological control of slugs by carabids. Biol. Control. Slugs Carabidsr 1989, 41, 295–300. [Google Scholar]
- Pan, Y.N.; Yao, M.Y.; Fu, D.; Guo, F.; Qin, R.; Jiang, L.; Chen, W.L. Predation Effects of Eocanthecona furcellata Adults Against Spodoptera frugiperda Larvae. J. Mt. Agric. Biol. 2020, 39, 23–27. [Google Scholar]
- Zeng, G.; Zhi, J.R.; Zhang, C.R.; Zhang, T.; Ye, J.Q.; Zhou, L.; Hu, C.X.; Ye, M. Orius similis (Hemiptera: Anthocoridae): A Promising Candidate Predator of Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Econ. Entomol. 2021, 114, 582–589. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, H.M.; Yin, Y.Q.; Li, X.Y.; Zhao, X.Q.; Tang, Y.T.; Wang, M.Q.; Shen, A.D.; Chen, F.S.; Zhang, L.S. Predation of adult of Arm achinensis to larvae of spodoptera frugiperda. Plant. Prot. 2019, 45, 42–46. [Google Scholar]
- Yu, H.Z. Research Progress of Insect Antennal Sensilla. J. Anhui Agric. Sci. 2007, 35, 4238. [Google Scholar]
- Mou, Z.G. Studies on Biology and Predacious Function of Calosoma maximoviczi Morowitzi. Shandong Agric. Univ. 2005. [Google Scholar]
- Zhou, J.H.; Li, P.L.; Naiwuzhati, Z.N.; Zheng, H.N.; Huang, J.; Wang, Z.H. Functional response and predation preference of ladybeetle Propylea japonica to Asian citrus psyllid Diaphorina citri. J. Plant Prot. 2020, 47, 1062–1070. [Google Scholar]
- Li, P.; Zhu, J.X.; Mo, X.N.; Lin, X.M.; Liu, T.Z.; Li, J.H.; Cai, D.C. The functional response of Menochilus sexmaculata on Megal. Usitatus. Chin. J. Biol. Control. 2021, 1–13. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Prey Ages | Predator Sexes | R2 | a’ | Th | Maximum Daily Prey Consumption |
|---|---|---|---|---|---|
| Juvenile | Female | 0.9871 | 0.7027 ± 0.0313 ab | 0.0198 ± 0.0014 b | 35.4895 |
| Male | 0.9762 | 0.6107 ± 0.0364 bc | 0.0227 ± 0.0030 ab | 26.9042 | |
| Immature | Female | 0.9642 | 0.7952 ± 0.0800 a | 0.0317 ± 0.0090 ab | 25.0841 |
| Male | 0.9691 | 0.6550 ± 0.0073 abc | 0.0323 ± 0.0065 ab | 20.2789 | |
| Adult | Female | 0.9717 | 0.7206 ± 0.0612 ab | 0.0421 ± 0.0054 a | 17.1168 |
| Male | 0.9148 | 0.5175 ± 0.0664 c | 0.0445 ± 0.0115 a | 11.6290 |
| Prey Ages | Predator Ages | R2 | Q | m |
|---|---|---|---|---|
| Juvenile | Female | 0.9983 | 0.5008 ± 0.0036 a | 1.5112 ± 0.0111 ab |
| Male | 0.9956 | 0.4886 ± 0.0086 a | 1.5561 ± 0.0307 ab | |
| Immature | Female | 0.9958 | 0.4273 ± 0.0054 b | 1.4913 ± 0.0230 c |
| Male | 0.9931 | 0.4392 ± 0.0066 b | 1.5084 ± 0.0238 bc | |
| Adult | Female | 0.9918 | 0.4023 ± 0.0038 c | 1.5598 ± 0.0177 bc |
| Male | 0.9915 | 0.3953 ± 0.0040 c | 1.6134 ± 0.0182 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, L.; Zhao, R.; Tian, H.; Wu, X.; Guo, F.; Chen, W. Functional Response and Predation Potential of Carabus elysii Adults against the Terrestrial Slug Agriolimax agrestis. Insects 2021, 12, 1135. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121135
Jiang L, Zhao R, Tian H, Wu X, Guo F, Chen W. Functional Response and Predation Potential of Carabus elysii Adults against the Terrestrial Slug Agriolimax agrestis. Insects. 2021; 12(12):1135. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121135
Chicago/Turabian StyleJiang, Lin, Runa Zhao, Hui Tian, Xuesan Wu, Feng Guo, and Wenlong Chen. 2021. "Functional Response and Predation Potential of Carabus elysii Adults against the Terrestrial Slug Agriolimax agrestis" Insects 12, no. 12: 1135. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121135