Effects of Imunit Insecticide on Biological Characteristics and Life Table Parameters of Spodoptera cilium (Lepidoptera: Noctuidae)


 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Rearing of Spodoptera Cilium
2.2. Insecticide Tested
2.3. Dose-Mortality Response Bioassay
2.4. Imunit Effects on Parental Generation
2.5. Imunit Effects on Offspring Generation
2.6. Data Analysis
3. Results
3.1. Estimation of Concentrations
3.2. Imunit Effects on Parental Generation
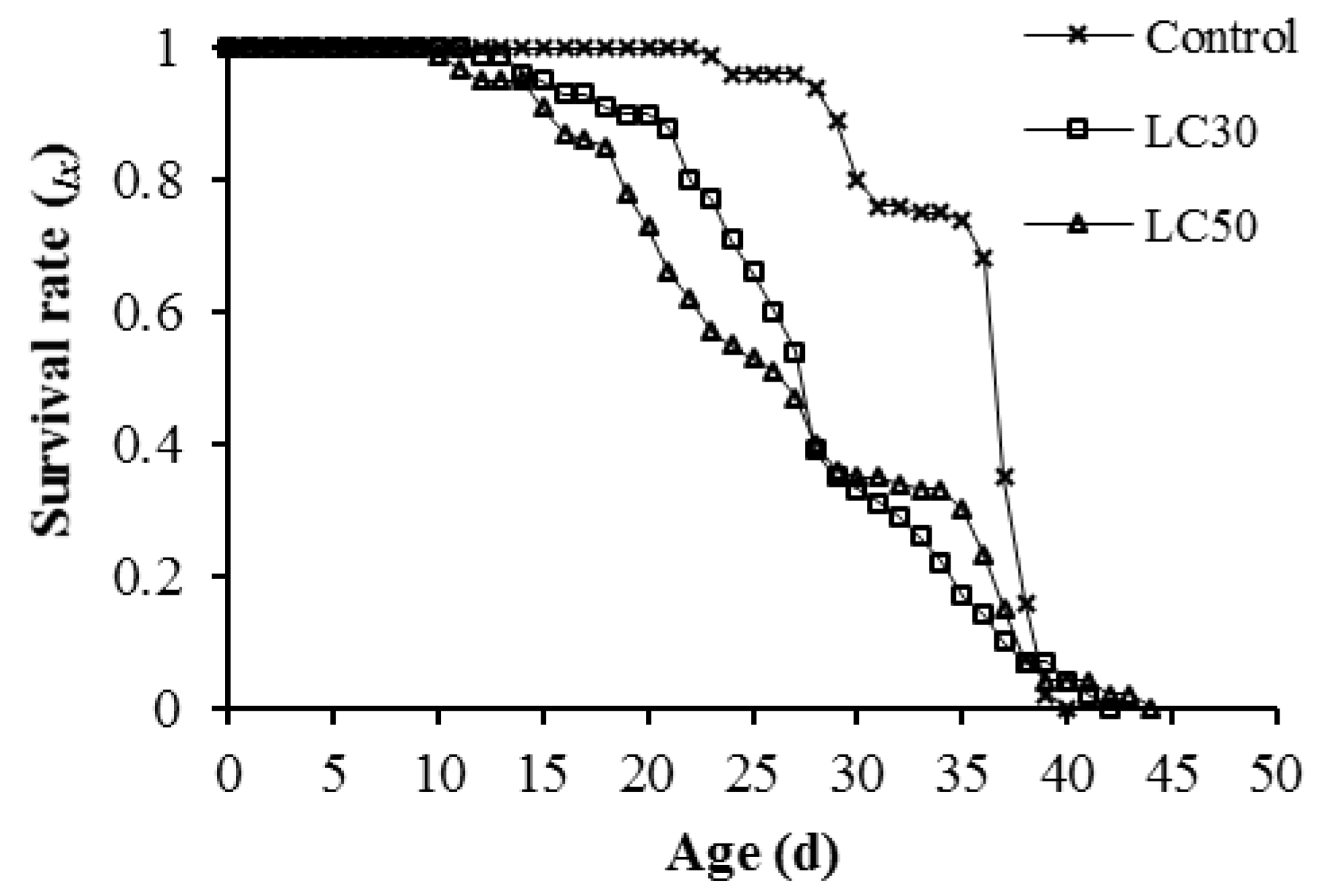
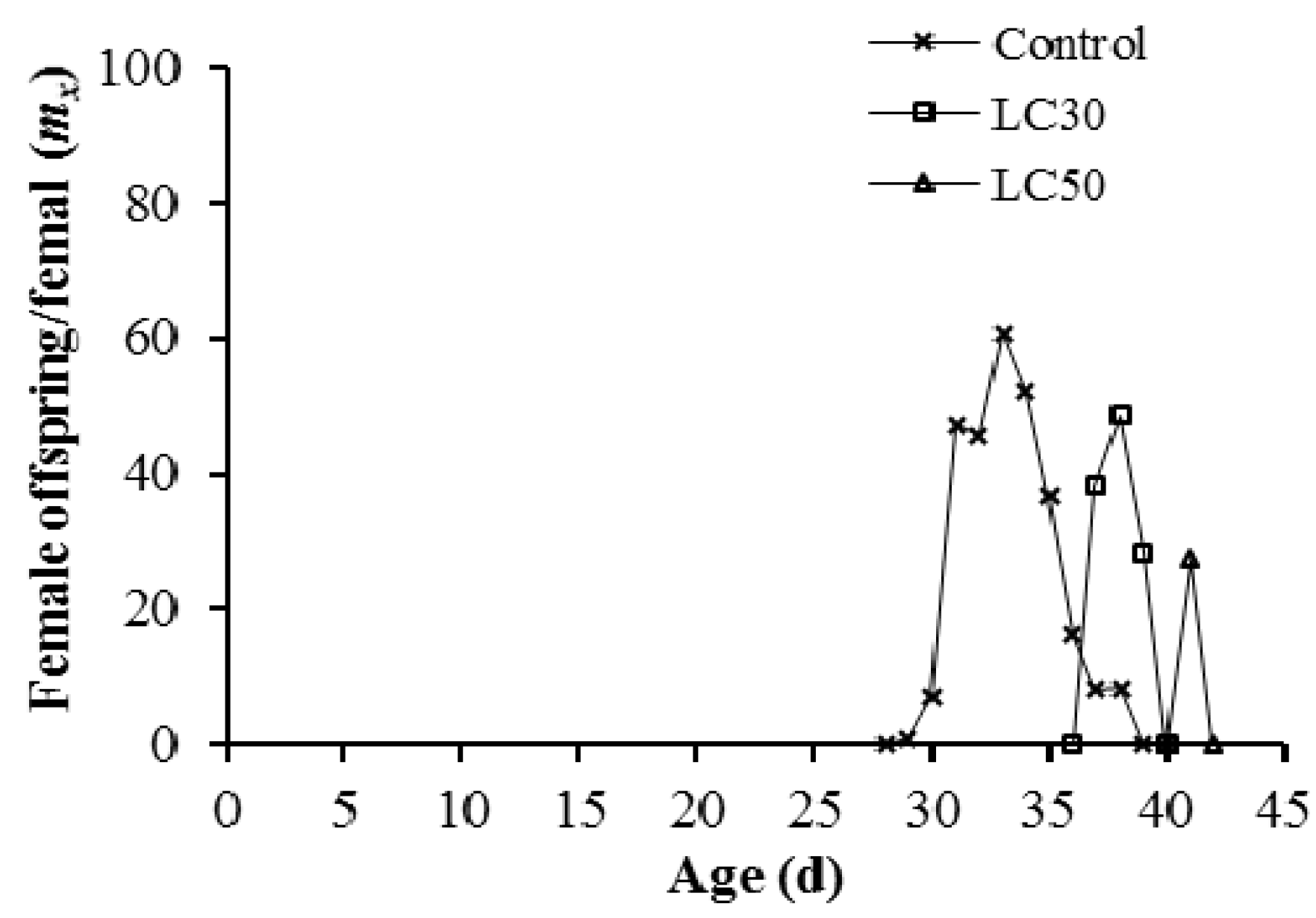
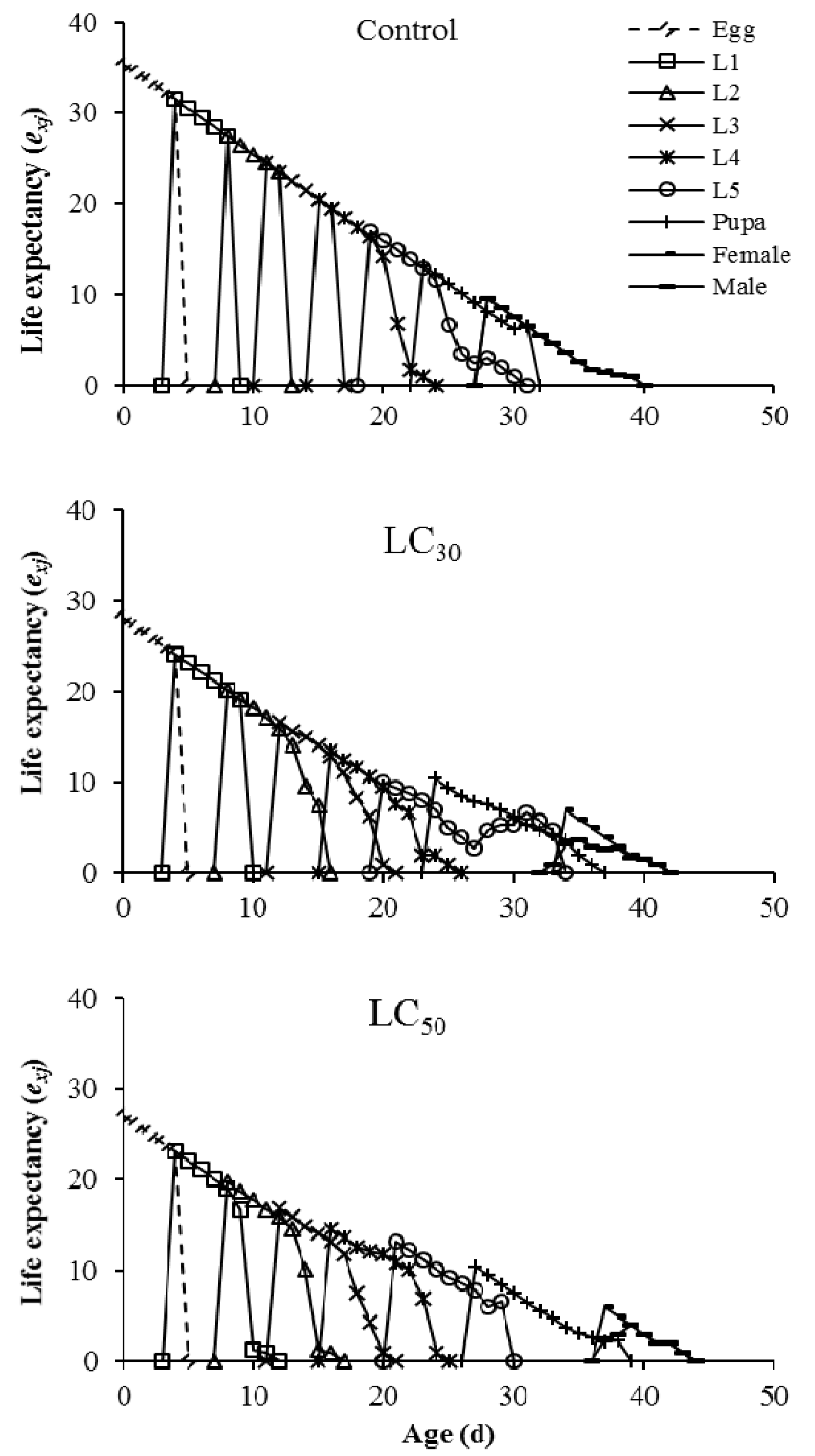
3.3. Imunit Effects on Offspring Generation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Capinera, J. Order Lepidoptera—Caterpillars, Moths and Butterflies. In Handbook of Vegetable Pests; Capinera, J., Ed.; Academic Press: San Diego, CA, USA, 2020; pp. 353–510. [Google Scholar]
- Brown, E.; Dewhurst, C. The genus Spodoptera (Lepidoptera, Noctuidae) in Africa and the near east. Bull. Entomol. Res. 1975, 65, 221–262. [Google Scholar] [CrossRef]
- Heinrichs, E.A.; Barrion, A.T. Rice-Feeding Insects and Selected Natural Enemies in West Africa: Biology, Ecology, Identification; International Rice Research Institute and Abidjan (Côte d’Ivoire), WARDA—The Africa Rice Center: Los Baños, Philippines, 2004. [Google Scholar]
- Aktar, M.W.; Sengupta, D.; Chowdhury, A. Impact of pesticides use in agriculture: Their benefits and hazards. Interdiscip. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegwart, M.; Graillot, B.; Blachere Lopez, C.; Besse, S.; Bardin, M.; Nicot, P.C.; Lopez-Ferber, M. Resistance to bio-insecticides or how to enhance their sustainability: A review. Front. Plant Sci. 2015, 6, 381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De França, S.M.; Breda, M.O.; Barbosa, D.R.; Araujo, A.M.; Guedes, C.A. The sublethal effects of insecticides in insects. In Biological Control of Pest and Vector Insects; Shields, V.D.C., Ed.; IntechOpen: London, UK, 2017; pp. 23–39. [Google Scholar]
- Storch, G.; Loak, A.E.; Borba, R.S.; Magano, D.A.; Moraes, C.L.; Grutzmacher, C.L. The effect of sub-lethal doses of insecticides on artificial diet and caterpillars of Anticarsia gemmatalis (Lepidoptera: Noctuidae). Rev. Bras. Agrociência 2007, 13, 175–179. [Google Scholar]
- Yin, X.-H.; Wu, Q.-J.; Li, X.-F.; Zhang, Y.-J.; Xu, B.-Y. Demographic changes in multigeneration Plutella xylostella (Lepidoptera: Plutellidae) after exposure to sublethal concentrations of spinosad. J. Econ. Entomol. 2009, 102, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wang, Y.-M.; Liu, H.-Y.; Xin, Z.; Xue, M. Lethal and sublethal effects of spinosad on Spodoptera exigua (Lepidoptera: Noctuidae). J. Econ. Entomol. 2013, 106, 1825–1831. [Google Scholar] [CrossRef]
- Mahmoudvand, M.; Abbasipour, H.; Garjan, A.S.; Bandani, A.R. Decrease in pupation and adult emergence of Plutella xylostella (L.) treated with hexaflumuron. Chil. J. Agric. Res. 2012, 72, 206–211. [Google Scholar] [CrossRef] [Green Version]
- Han, W.; Zhang, S.; Shen, F.; Liu, M.; Ren, C.; Gao, X. Residual toxicity and sublethal effects of chlorantraniliprole on Plutella xylostella (Lepidoptera: Plutellidae). Pest Manag. Sci. 2012, 68, 1184–1190. [Google Scholar] [CrossRef]
- Dong, J.; Wang, K.; Li, Y.; Wang, S. Lethal and sublethal effects of cyantraniliprole on Helicoverpa assulta (Lepidoptera: Noctuidae). Pest Biochem. Physiol. 2017, 136, 58–63. [Google Scholar] [CrossRef]
- Hatami, M.; Seraj, A.; Ziaee, M.; Mehrabi-Koushki, M. Efficacy of different insecticides in control of Spodoptera cilium in laboratory and field conditions. Plant Pests Res. 2020, 10, 1–15. [Google Scholar]
- Oberlander, H.; Silhacek, D. New perspectives on the mode of action of benzoylphenyl urea insecticides. In Insecticides with Novel Modes of Action; Springer: Berlin, Germany, 1998; pp. 92–105. [Google Scholar]
- Chi, H.; Liu, H. Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Chi, H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Chi, H. TIMING-MSChart: Computer Program for Population Projection Based on Age-Stage, Two-Sex Life Table; National Chung Hsing University: Taichung, Taiwan, 2020. [Google Scholar]
- Goodman, D. Optimal life histories, optimal notation, and the value of reproductive value. Am. Nat. 1982, 119, 803–823. [Google Scholar] [CrossRef]
- Huang, Y.-B.; Chi, H. Assessing the application of the jackknife and bootstrap techniques to the estimation of the variability of the net reproductive rate and gross reproductive rate: A case study in Bactrocera cucurbitae (Coquillett) (Diptera: Tephritidae). Agric. For. Entomol. 2012, 61, 37–45. [Google Scholar]
- Göldel, B.; Lemic, D.; Bažok, R. Alternatives to synthetic insecticides in the control of the colorado potato beetle (Leptinotarsa decemlineata Say) and their environmental benefits. Agriculture 2020, 10, 611. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Tennekes, H.A.; Goka, K. Impact of systemic insecticides on organisms and ecosystems. In Insecticides-Development of Safer and More Effective Technologies; InTech: Rijeka, Croatia, 2013; pp. 365–414. [Google Scholar]
- Clarke, B.S.; Jewess, P.J. The inhibition of chitin synthesis in Spodoptera littoralis larvae by flufenoxuron, teflubenzuron and diflubenzuron. Pest Sci. 1990, 28, 377–388. [Google Scholar] [CrossRef]
- Rodríguez Enríquez, C.L.; Pineda, S.; Figueroa, J.I.; Schneider, M.I.; Martínez, A.M. Toxicity and sublethal effects of methoxyfenozide on Spodoptera exigua (Lepidoptera: Noctuidae). J. Econ. Entomol. 2010, 103, 662–667. [Google Scholar] [CrossRef] [Green Version]
- Mahmoudvand, M.; Abbasipour, H.; Garjan, A.S.; Bandani, A.R. Sublethal effects of hexaflumuron on development and reproduction of the diamondback moth, Plutella xylostella (Lepidoptera: Yponomeutidae). Insect Sci. 2011, 18, 689–696. [Google Scholar] [CrossRef]
- Biddinger, D.; Hull, L. Sublethal effects of selected insecticides on growth and reproduction of a laboratory susceptible strain of tufted apple bud moth (Lepidoptera: Tortricidae). J. Econ. Entomol. 1999, 92, 314–324. [Google Scholar] [CrossRef]
- Josan, A.; Singh, G. Sublethal effects of lufenuron on the diamondback moth, Plutella xylostella (Linnaeus). Insect Sci. Appl. 2000, 20, 303–308. [Google Scholar] [CrossRef]
- Meng, Q.-W.; Wang, J.-J.; Shi, J.-F.; Guo, W.-C.; Li, G.-Q. Effect of teflubenzuron ingestion on larval performance and chitin content in Leptinotarsa decemlineata. Am. J. Potato Res. 2018, 95, 463–472. [Google Scholar] [CrossRef]
- Xu, C.; Zhang, Z.; Cui, K.; Zhao, Y.; Han, J.; Liu, F.; Mu, F. Effects of sublethal concentrations of cyantraniliprole on the development, fecundity and nutritional physiology of the black cutworm Agrotis ipsilon (Lepidoptera: Noctuidae). PLoS ONE 2016, 11, e0156555. [Google Scholar] [CrossRef] [Green Version]
- Tanzubil, P.B.; McCaffery, A.R. Effects of Azadirachtin on reproduction in the African armyworm (Spodoptera exempta). Entomol. Exp. Appl. 1990, 57, 115–121. [Google Scholar] [CrossRef]
- Adel, M.M.; Sehnal, F. Azadirachtin potentiates the action of ecdysteroid agonist RH-2485 in Spodoptera littoralis. J. Insect Physiol. 2000, 46, 267–274. [Google Scholar] [CrossRef]
- Pineda, S.; Martínez, A.M.; Figueroa, J.I.; Schneider, M.I.; Del Estal, P.; Viñuela, E.; Gómez, B.; Smagghe, G.; Budia, F. Influence of azadirachtin and methoxyfenozide on life parameters of Spodoptera littoralis (Lepidoptera: Noctuidae). J. Econ. Entomol. 2009, 102, 1490–1496. [Google Scholar] [CrossRef]
- Guo, L.; Desneux, N.; Sonoda, S.; Liang, P.; Han, P.; Gao, X.-W. Sublethal and transgenerational effects of chlorantraniliprole on biological traits of the diamondback moth, Plutella xylostella L. Crop Prot. 2013, 48, 29–34. [Google Scholar] [CrossRef]
- Ahmad, S.; Ansari, M.S. Acute toxicity and sub-lethal effects of a pyrethroid (cypermethrin) on survival, development and fitness of Helicoverpa armigera. Arch. Phytopathol. Plant Prot. 2013, 46, 1726–1739. [Google Scholar] [CrossRef]
- Song, L.; Zhang, J. Sublethal effects of indoxacarb and beta-cypermethrin on Plutella xylostella (Lepidoptera: Plutellidae). Acta Entomol. Sin. 2013, 56, 521–529. [Google Scholar]
- Quan, L.-F.; Qiu, G.-S.; Zhang, H.-J.; Sun, L.-N.; Li, Y.-Y.; Yan, W.-T. Sublethal concentration of beta-cypermethrin influences fecundity and mating behavior of Carposina sasakii (Lepidoptera: Carposinidae) adults. J. Econ. Entomol. 2016, 109, 2196–2204. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Insecticide | Number of Larvae Tested | df | LC50 (ppm) | LC30 (ppm) | Chi Square | p Value |
|---|---|---|---|---|---|---|
| Imunit | 375 | 3 | 27.7 | 18.2 | 3.60 | 0.40 |
| Life Stage | Treatments | ||
|---|---|---|---|
| Control | LC30 | LC50 | |
| Third-instar larvae (d) | 4.00 ± 0.041 a | 4.10 ± 0.03 a | 4.08 ± 0.3 a |
| Fourth-instar larvae (d) | 3.93 ± 0.042 b | 4.06 ± 0.03 a | 4.03 ± 0.02 a |
| Fifth-instar larvae (d) | 4.15 ± 0.08 a | 4.07 ± 0.05 a | 4.05 ± 0.05 a |
| Pupae (d) | 5.23 ± 0.06 c | 6.48 ± 0.17 b | 7.17 ± 0.19 a |
| Adult (d) | 8.28 ± 0.17 a | 7.52 ± 0.31 a | 5.61 ± 0.43 b |
| Oviposition (d) | 4.83 ± 0.23 a | 3.93 ± 0.25 b | 3.40 ± 0.45 b |
| Fecundity (eggs/female) | 837.4 ± 22.54 a | 514.3 ± 43.89 b | 434.3 ± 24.95 c |
| Life Stage | Treatments | ||
|---|---|---|---|
| Control | LC30 | LC50 | |
| Egg (d) | 4.03 ± 0.04 c | 4.20 ± 0.04 b | 4.78 ± 0.04 a |
| First-instar larvae (d) | 4.02 ± 0.01 c | 4.28 ± 0.04 b | 4.44 ± 0.05 a |
| Second-instar larvae (d) | 4.09 ± 0.03 b | 4.22 ± 0.04 b | 4.62 ± 0.05 a |
| Third-instar larvae (d) | 4.03 ± 0.03 a | 4.08 ± 0.03 a | 4.12 ± 0.05 a |
| Fourth-instar larvae (d) | 4.07 ± 0.04 c | 4.42 ± 0.06 b | 4.96 ± 0.26 a |
| Fifth-instar larvae (d) | 4.16 ± 0.04 b | 4.55 ± 0.27 b | 5.23 ± 0.10 a |
| Pupae (d) | 5.33 ± 0.05 b | 9.88 ± 0.12 a | 9.50 ± 0.34 a |
| Pre-adult (d) | 29.63 ± 0.09 c | 34.56 ± 0.18 b | 37.67 ± 0.33 a |
| Adult (d) | 7.91 ± 0.08 a | 3.81 ± 0.53 b | 3.83 ± 0.91 b |
| Oviposition (d) | 5.61 ± 0.17 a | 2.00 ± 0.00 b | 1.00 ± 0.00 c |
| APOP 1 (d) | 1.14 ± 0.07 c | 2.67 ± 0.33 b | 3.33 ± 0.33 a |
| TPOP 2 (d) | 30.61 ± 0.16 c | 37.33 ± 0.33 b | 41.00 ± 0.00 a |
| Fecundity (eggs/female) | 728.21 ± 85.70 a | 306.33 ± 82.28 b | 195.0 ±55.0 c |
| Adult longevity (male) (d) | 37.52 ± 0.17 b | 37.77 ± 0.68 b | 40.75 ± 1.38 a |
| Adult longevity (female) (d) | 37.57 ± 0.21 c | 41.00 ± 0.58 b | 43.00 ± 0.57 a |
| Total longevity (all individuals) (d) | 35.43 ± 0.407 a | 28.18 ± 0.690 b | 27.09 ± 0.88 b |
| Parameters | Treatments | ||
|---|---|---|---|
| Control | LC30 | LC50 | |
| GRR (offspring/female) | 282.9 ± 52.75 a | 114.9 ± 55.65 b | 97.5 ± 51.41 b |
| R0 (offspring/female) | 203.9 ± 40.15 a | 9.190 ± 5.32 b | 3.900 ± 2.58 b |
| r (d−1) | 0.160 ± 0.01 a | 0.050 ± 0.02 b | 0.032 ± 0.01 c |
| λ (d−1) | 1.170 ± 0.01 a | 1.050 ± 0.02 b | 1.030 ± 0.01 b |
| T (d) | 33.87 ± 0.27 c | 38.78 ± 0.26 b | 42.00 ± 0.02 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hatami, M.; Ziaee, M.; Seraj, A.A.; Mehrabi-Koushki, M.; Francikowski, J. Effects of Imunit Insecticide on Biological Characteristics and Life Table Parameters of Spodoptera cilium (Lepidoptera: Noctuidae). Insects 2021, 12, 1138. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121138
Hatami M, Ziaee M, Seraj AA, Mehrabi-Koushki M, Francikowski J. Effects of Imunit Insecticide on Biological Characteristics and Life Table Parameters of Spodoptera cilium (Lepidoptera: Noctuidae). Insects. 2021; 12(12):1138. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121138
Chicago/Turabian StyleHatami, Marzieh, Masumeh Ziaee, Ali Asghar Seraj, Mehdi Mehrabi-Koushki, and Jacek Francikowski. 2021. "Effects of Imunit Insecticide on Biological Characteristics and Life Table Parameters of Spodoptera cilium (Lepidoptera: Noctuidae)" Insects 12, no. 12: 1138. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121138