
Aphid Assemblages Associated with Urban Park Plant Communities

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Methods of Botanical Research
2.3. Methods of Entomological Research
3. Results and Discussion
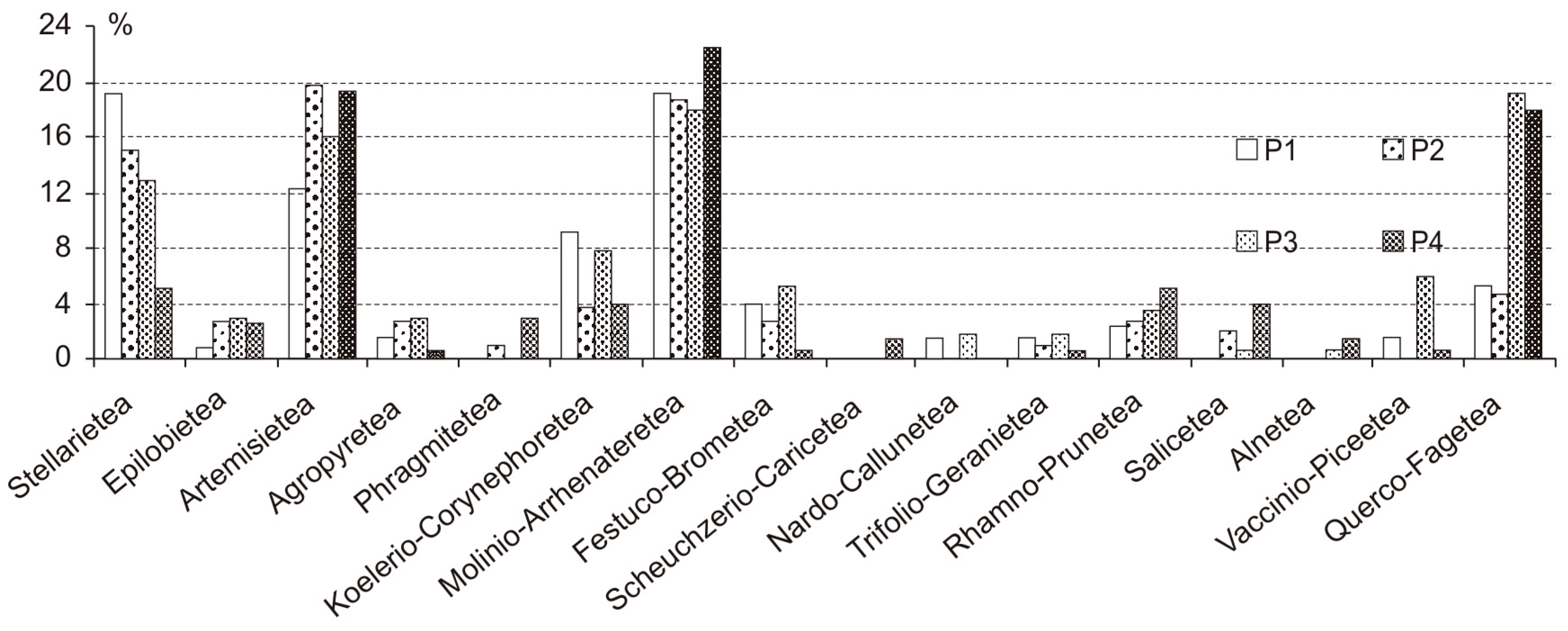
3.1. Flora in the Parks
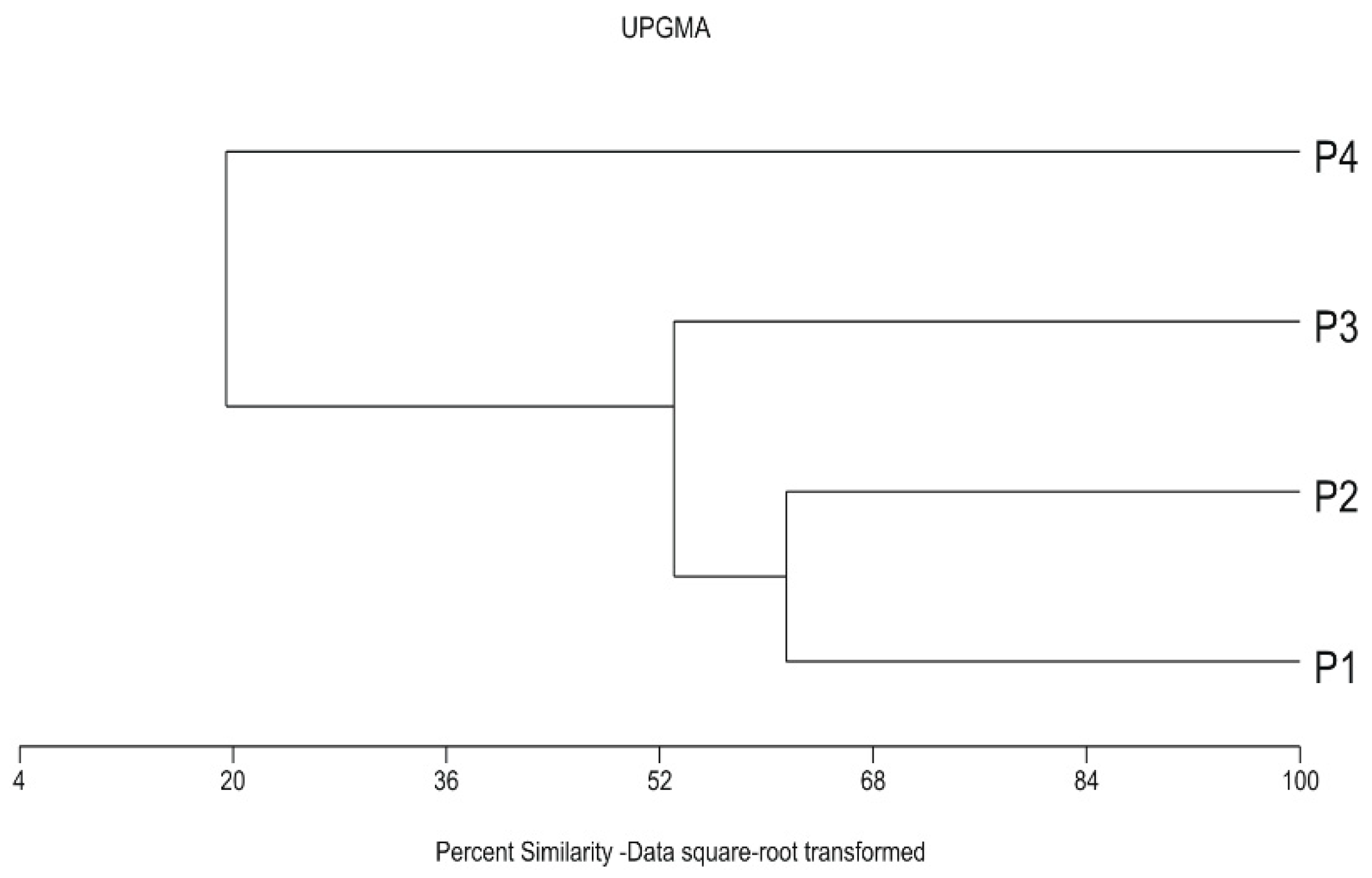
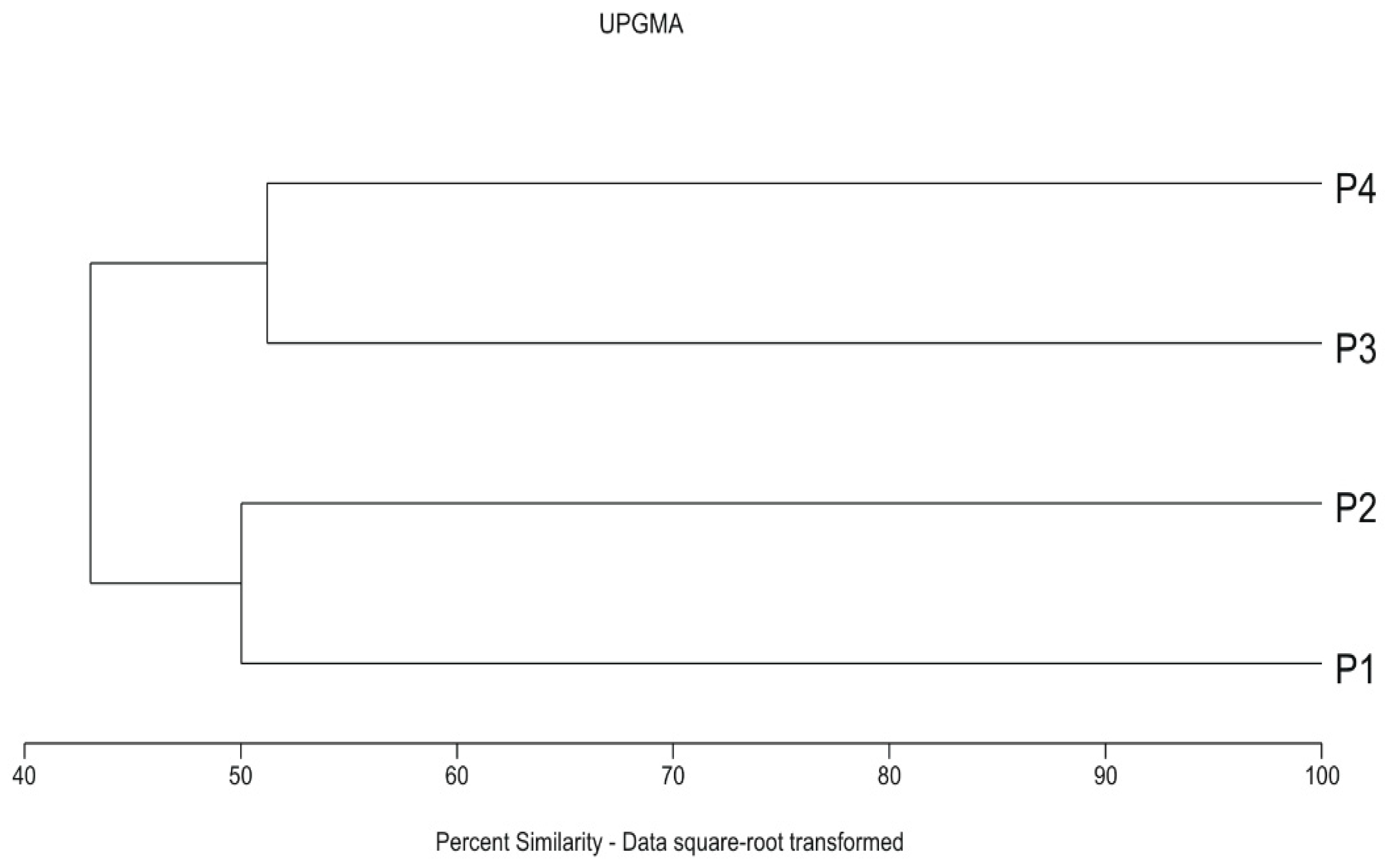
3.2. Aphid Assemblages
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zapparoli, M. Fratelli Palombi. In Gli Insetti di Roma; Editori: Rome, Italy, 1997; ISBN 88-7621-125-X. [Google Scholar]
- Gibson, C.W.D. Brownfield. Red Data. The Values Artificial Habitats Have for Uncommon Invertebrates; No. 273-English Nature Research Reports; English Nature: Oxford, UK, 1998.
- Weller, B.; Ganzhorn, J.U. Carabid Beetle Community Composition, Body Size, and Fluctuating Asymmetry along an Urban-Rural Gradient. Basic Appl. Ecol. 2004, 5, 193–201. [Google Scholar] [CrossRef]
- Breuste, J.; Niemelä, J.; Snep, R.P.H. Applying Landscape Ecological Principles in Urban Environments. Landsc. Ecol. 2008, 23, 1139–1142. [Google Scholar] [CrossRef]
- Bennewicz, J.; Barczak, T. Aphids in Urban Environments. In Urban Fauna. Animal, Man and the City-Intractions and Relationships; Indykiewicz, P., Böhner, J., Eds.; ArtStudio: Bydgoszcz, Poland, 2014; pp. 65–75. ISBN 978-83-936060-5-4. [Google Scholar]
- Jones, E.L.; Leather, S.R. Invertebrates in Urban Areas: A Review. Eur. J. Entomol. 2012, 109, 463–478. [Google Scholar] [CrossRef] [Green Version]
- Czechowski, W. Occurrence of Carabids (Coleoptera, Carabidae) in the Urban Greenery of Warsaw according to the Land Utilization and Cultivation. Memorab. Zool. 1982, 39, 3–108. [Google Scholar]
- McIntyre, N.E. Ecology of Urban Arthropods: A Review and a Call to Action. Ann. Entomol. Soc. Am. 2000, 93, 825–835. [Google Scholar] [CrossRef]
- Haq, S.M.A. Urban Green Spaces and an Integrative Approach to Sustainable Environment. J. Environ. Prot. 2011, 2, 601–608. [Google Scholar] [CrossRef] [Green Version]
- McIntyre, N.E.; Rango, J.; Fagan, W.F.; Faeth, S.H. Ground Arthropod Community Structure in a Heterogeneous Urban Environment. Landsc. Urban Plan. 2001, 52, 257–274. [Google Scholar] [CrossRef]
- Jones, M.E.; Paine, T.D. Detecting Changes in Insect Herbivore Communities along a Pollution Gradient. Environ. Pollut. 2006, 143, 377–387. [Google Scholar] [CrossRef]
- Wäckers, F.L.; van Rijn, P.C.J.; Heimpel, G.E. Honeydew as a Food Source for Natural Enemies: Making the Best of a Bad Meal? Biol. Control. 2008, 45, 176–184. [Google Scholar] [CrossRef] [Green Version]
- Kamiński, P.; Barczak, T.; Bennewicz, J.; Jerzak, L.; Bogdzińska, M.; Aleksandrowicz, O.; Koim-Puchowska, B.; Szady-Grad, M.; Klawe, J.J.; Woźniak, A. Effects of Chemical Elements in the Trophic Levels of Natural Salt Marshes. Environ. Geochem. Health 2016, 38, 783–810. [Google Scholar] [CrossRef]
- Tena, A.; Wackers, F.L.; Heimpel, G.E.; Urbaneja, A.; Pekas, A. Parasitoid Nutritional Ecology in a Community Context: The Importance of Honeydew and Implications for Biological Control. Curr. Opin. Insect Sci. 2016, 14, 100–104. [Google Scholar] [CrossRef]
- Ratajczak, J.; Wilkaniec, B.; Wilkaniec, A. Infestatoin of Dendrological Collection in Central Poland by Aphids. Dendrobiology 2011, 66, 85–97. [Google Scholar]
- Jaśkiewicz, B. Analysis of the Aphid Population Colonizing Roses in Different Types of City Green Areas of Lublin. Acta Sci. Pol. Hortorum Cultus 2005, 4, 129–137. [Google Scholar]
- Trojan, P.; Winiarska, G. City as an Archipelago of Inland Isles. In Biodiversity and Ecology of Animal Population in Urban Environments; Indykiewicz, P., Barczak, T., Kaczorowski, G., Eds.; Wyd. NICE: Bydgoszcz, Poland, 2001; pp. 10–16. ISBN 83-88716-11-5. [Google Scholar]
- Barczak, T. Some Aspects of Bionomics and Ecology of Spiral Gall Aphid, Pemphigus Spirothecae Pass. (Homoptera: Pemphigidae) in Urban Environment. In Urban Fauna of Central Europe in the 21st Century; Indykiewicz, P., Barczak, T., Eds.; P.M. LOGO: Bydgoszcz, Poland, 2004; pp. 235–255. ISBN 83-87586-34-X. [Google Scholar]
- Bennewicz, J.; Barczak, T. Mszyce Środowisk Zurbanizowanych. In Fauna Miast. Zwierzę, Człowiek, Miasto–Wzajemne Oddziaływania i Zależnośc. Abstract Volume; Indykiewicz, P., Böhner, J., Eds.; UTP: Bydgoszcz, Poland, 2014; p. 14. [Google Scholar]
- Bennewicz, J.; Barczak, T.; Korczyński, M. Urban Greenery Aphids (Hemiptera: Aphididae). Arch. Biol. Sci. 2013, 65, 1053–1061. [Google Scholar] [CrossRef] [Green Version]
- Bennewicz, J.; Barczak, T. Aphids (Hemiptera: Aphididae) of Different Plant Communities in an Urban Environment. Biologia 2016, 71, 583–592. [Google Scholar] [CrossRef]
- Ruszkowska, M.; Strażyński, P. Elements of Changes in the Bionomy of Aphids on Trees in the Urbanized Environment. Zesz. Probl. Post. Nauk Roln. 2011, 562, 221–227. [Google Scholar]
- Jaśkiewicz, B.; Kmieć, K. Population Dynamics of Chaetosiphon Tetrarhodus (Walk) in Urban Conditions. Aphids Other Hemipterous Insects 2005, 11, 53–61. [Google Scholar]
- Cichocka, E.; Goszczyński, W. The Impact of Urban Pressure on Species Composition and Number of Arthropoda on Trees in a City on the Example of Warsaw. Aphids Other Hemipterous Insects 2008, 14, 63–72. [Google Scholar]
- Maćkoś, E. Aphids (Hemiptera, Aphidoidea) on Mape Acer Platanoiddes in the Urban Green Areas of the City of Lublin. Aphids Other Hemipterous Insects 2008, 14, 91–102. [Google Scholar]
- Lubiarz, M. Number Dynamics of Thelaxes dryophila (Schrank, 1801) Hemiptera, Aphidoidea on Common Oak (Quercus Robur) in Natural and Degraded Landscape. Aphids Other Hemipterous Insects 2008, 14, 73–82. [Google Scholar]
- Harrington, R.; Bale, J.S.; Tatchell, G.M. Aphids in a Changing Climate. In Insects in a Changing Environment; Harrington, R., Stork, N.E., Eds.; Academic Press: London, UK, 1995; pp. 125–155. [Google Scholar]
- Wieczorek, K.; Osiadacz, B. Dendrophilous Aphids (Hemiptera: Aphidoidea) of the Managed Urban Greenery of Katowice—Part I “Tadeusz Kościuszko Park”. Acta Entomol. Sil. 2005, 12–13, 155–160. [Google Scholar]
- Osiadacz, B.; Wieczorek, K. Aphids (Hemiptera: Aphidoidea) of Parks in Bytom (Upper Silesia, Poland). Acta Entomol. Sil. 2003, 11, 39–46. [Google Scholar]
- Borowiak-Sobkowiak, B.; Wilkaniec, B. Occurrence of Aphids (Hemiptera: Aphididae) on Tree and Shrubs in Cytadela Park in Poznań. Aphids Other Hemipterous Insects 2010, 16, 27–35. [Google Scholar]
- Braun-Blanquet, J. Pflanzensoziologie. Grundzűge der Vegetationskunde. Biologische Studientbűcher 7; Springer: Berlin/Heidelberg, Germany, 1928; ISBN 978-3-662-02056-2. [Google Scholar]
- Rutkowski, L. The Key for the Determination of Lowland Poland Vascular Plants, 13th ed.; Wydawnictwo Naukowe PWN (Polish Scientific Publishers): Warszawa, Poland, 2019; ISBN 978-83-01-14342-8. [Google Scholar]
- Mirek, Z.; Piękoś-Mirkowa, H.; Zając, A.; Zając, M. Flowering Plants and Pteridophytes of Poland. A Checklist. In Biodiversity of Poland; Mirek, Z., Ed.; Szafer Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 2002; pp. 21–188. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1963; ISBN 02527254849780252725487. Available online: https://pure.mpg.de›items›file_2383163›content (accessed on 8 January 2021).
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, V.; Werner, W.; Paulissen, D. Zeigerwerte von Pflanzen in Mitteleuropa. 2. und Verbesserte Auflage. Scr. Geobot. 1992, 18, 1–258. [Google Scholar]
- Chmiel, J. Flora of Vascular Plants in the Eastern Part of the Gniezno Lake District and Its Anthropogenic Transformations in the 19th and 20th Centuries. Pr. Zakładu Taksonomii Uniw. A Mickiewicza 1993, 1, 1–199. [Google Scholar]
- Kovach, W.L. MVSP-A Multi Variate Statistical Package for Windows, Ver. 3.1; Kovach Computing Services: Wales, UK, 2007; Available online: https://www.kovcomp.co.uk›mvsp3man (accessed on 8 January 2021).
- Matuszkiewicz, W. Guide to Marking Plant Communities in Poland; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2005; ISBN 9788301167073. [Google Scholar]
- Luszkiewicz, A.; Słaby, T. Statistics with the STATISTICA.PL Computer Package. Theory and Applications; Wydawnictwo, C.H. Beck: Warszawa, Poland, 2008; ISBN 978-83-255-0113-6. [Google Scholar]
- Oktaba, W. Elements of Mathematical Statistics and Experimental Methodology; Wydawnictwo AR: Lublin, Poland, 1988. [Google Scholar]
- Zar, J.H. Biostatistical Analysis, 4th ed.; Prentice-Hall, Inc.: Englewood Cliffs, NJ, USA, 1998; ISBN 9780130815422. [Google Scholar]
- Hill, T.; Lewicki, P. Statistics. Methods and Applications. A Comprehensive Reference for Science, Industry and Data Mining, 1st ed.; StatSoft Inc.: Tulsa, OK, USA, 2006. Available online: https://trove.nla.gov.au (accessed on 8 January 2021).
- Blackman, R.L.; Eastop, V.F. Aphids on the World’s Crops: An Identification and Information Guide; J. Wiley & Sons Ltd.: New York, NY, USA, 2000; ISBN 978-0-471-85191-2. [Google Scholar]
- Szelegiewicz, H. Część XVII. Pluskwiaki równoskrzydłe—Homoptera. Zeszyt 5b. Mszyce—Aphidoidea. Chaitophoridae. In Klucze do Oznaczania Owadów Polski; Polskie Towarzystwo Entomologiczne-PWN: Warszawa, Poland, 1985; ISBN 83-01-06549-4. [Google Scholar]
- Taylor, L.R. A Handbook for Aphid Identification. Manuel D‘Identification des Pucerons; Euraphid-Rothamsted: Harpenden, UK, 1980. [Google Scholar]
- Klimaszewski, S.M.; Wojciechowski, W.; Czylok, A.; Gębicki, C.; Herczek, A.; Jasińska, J. Communities of Selected Groups of Homopterous Insects (Homoptera) and Heteropterous Insects (Heteroptera) in the Forests of the Katowice Region. Acta Biol. Sil. 1980, 8, 22–39. [Google Scholar]
- Pawlikowski, T. Wild Bee (Hymenoptera, Apoidea) Communities of the Different Parcel out Agricultural Areas. Stud. Soc. Sci. Tor. Sect. E 1985, 10, 1–82. [Google Scholar]
- Hutcheson, K. A Test for Comparing Diversities Based on the Shannon Formula. J. Theor. Biol. 1970, 29, 151–154. [Google Scholar] [CrossRef]
- Gower, J.C. Some Distance Properties of Latent Root and Vector Methods Used in Multivariate Analysis. Biometrika 1966, 53, 325–338. [Google Scholar] [CrossRef]
- Kavallieratos, N.G.; Athanassiou, C.G.; Stathas, G.J.; Tomanovic, Z. Aphid Parasitoids (Hymenoptera: Braconidae: Aphidiinae) on Citrus: Seasonal Abundance, Association with the Species of Host Plant, and Sampling Indices. Phytoparasitica 2002, 30, 365–377. [Google Scholar] [CrossRef]
- Barnes, D.K.A.; de Grave, S. Ecological Biogeography of Southern Polar Encrusting Faunas. J. Biogeogr. 2001, 28, 359–365. [Google Scholar] [CrossRef]
- Heywood, V.; Brunel, S. Code of Conduct on Horticulture and Invasive Alien Plants. Nature and Environment No. 162; Council of Europe Publishing: Strasbourg, France, 2011; Available online: https://www.academia.edu/28825644/European_Code_of_Conduct_or_Botanic_Gardens_on_Invasive_Alien_Species (accessed on 8 January 2021).
- Tokarska-Guzik, B.; Bzdęga, K.; Nowak, T.; Lewandowska, A.; Gancarek, M.; Frelich, M. Alien Plants in Poland: Research Directions and Putting the Results into Practice. Biodiv. Res. Conserv. 2014, 35, 57–74. [Google Scholar] [CrossRef] [Green Version]
- Paker, Y.; Yom-Tov, Y.; Alon-Mozes, T.; Barnea, A. The Effect of Plant Richness and Urban Garden Structure on Bird Species Richness, Diversity and Community Structure. Landsc. Urban Plan. 2014, 122, 186–195. [Google Scholar] [CrossRef]
- Wilkaniec, B.; Borowiak-Sobkowiak, B.; Wilkaniec, A.; Trzciński, P.; Kozłowska, M. Effect of Climate Change on Seasonal Flight Activity of Aphid Males in Urban Green Area. Acta Sci. Pol. Hortorum Cultus 2016, 15, 157–169. [Google Scholar]
- Detrain, C.; Verheggen, F.J.; Diez, L.; Wathelet, B.; Haubruge, E. Aphid—Ant Mutualism: How Honeydew Sugars Influence the Behaviour of Ant Scouts. Physiol. Entomol. 2010, 35, 168–174. [Google Scholar] [CrossRef]
- Barczak, T. Biocontrol in Agrocenosis and Plant Protection. Adv. Agric. Sci. 1994, 4, 19–28. [Google Scholar]
- Winiarska, W. Parasitic Larvae of Erythraeid Mites Associated with Aphis fabae Colonies. Pol. J. Entomol. 1998, 67, 151–153. [Google Scholar]
- Kean, J.M.; Müller, C.B. Can Competition Explain Local Rarity of a Nettle Aphid? Ecol. Entomol. 2004, 29, 706–710. [Google Scholar] [CrossRef]
- Wojciechowicz-Żytko, E.; Jankowska, B. Aphids and Their Predators Occurring on Same Shrubs in the Botanic Garden of the Jagiellonian University in Krakow. Aphids Other Hemipterous Insects 2011, 17, 147–156. [Google Scholar]
- Buitenhuis, R.; McNeil, J.N.; Boivin, G.; Brodeuri, J. The Role of Honeydew in Host Searching of Aphid Hyperparasitoids. J. Chem. Ecol. 2004, 30, 273–285. [Google Scholar] [CrossRef]
- Kant, R.; Minor, M.A.; Trewick, S.A. Reproductive Strategies of Diaeretiella rapae (Hymenoptera: Aphidiinae) during Fluctuating Temperatures of Spring Season in New Zealand. Biocontrol Sci. Techn. 2012, 22, 1–9. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Plant Species | Shannon Formula H’ | Anthropophytisation Rate | L | T | K | F | R | N | |
|---|---|---|---|---|---|---|---|---|---|
| P1 | 161 | 6.458 | 52.8% | 7.12 | 5.84 | 4.31 | 4.50 | 6.14 | 5.13 |
| P2 | 129 | 6.025 | 48.1% | 6.84 | 5.90 | 4.31 | 5.00 | 6.75 | 6.33 |
| P3 | 176 | 6.359 | 37.1% | 7.08 | 5.80 | 4.62 | 4.47 | 6.22 | 5.23 |
| P4 | 163 | 6.351 | 19.8% | 5.76 | 5.50 | 3.76 | 6.16 | 6.77 | 7.15 |
| Ecological Indicators | (A) All Plant Species | (B) All Aphid Species | (C) All Aphid Species and Potential Host Plant Species | |||
|---|---|---|---|---|---|---|
| No. of Axis | No. of Axis | No. of Axis | ||||
| 1 | 2 | 1 | 2 | 1 | 2 | |
| L | −0.9921 * | 0.0622 | 0.5577 | 0.7098 | 0.3791 | −0.8423 |
| T | −0.9662 * | −0.2640 | 0.5745 | 0.6899 | 0.3195 | −0.9530 * |
| K | −0.9028 | 0.3474 | 0.2059 | 0.8966 | 0.0318 | −0.8251 |
| F | 0.9694 * | −0.1701 | −0.5273 | −0.7004 | −0.3781 | 0.7829 |
| R | 0.6491 | −0.4549 | −0.5155 | −0.2926 | −0.5607 | 0.2581 |
| N | 0.8643 | −0.3180 | −0.5496 | −0.5383 | −0.4874 | 0.5680 |
| Plant Species | Aphid Species | Parks in Bydgoszcz | |||
|---|---|---|---|---|---|
| P1 | P2 | P3 | P4 | ||
| Aegopodium podagraria | Aphis podagrariae | 34.00 | |||
| Acer platanoides | Drepanosiphum platanoidis | 30.00 | |||
| Periphyllus testudinaceus | 468.67 | 259.01 | 495.01 | 1682.99 | |
| Acer pseudoplatanus | Drepanosiphum acerinum | 403.32 | 27.00 | 1.00 | |
| Periphyllus testudinaceus | 171.66 | 536.33 | 322.33 | 13.33 | |
| Acer negundo | Periphyllus testudinaceus | 23.00 | 39.67 | 135.00 | |
| Acer saccharinum | Periphyllus testudinaceus | 69.66 | |||
| Alnus glutinosa | Pterocallis albidus | 103.33 | 106.01 | ||
| Alnus viridis | Pterocallis albida | 127.64 | |||
| Angelica archangelica | Aphis fabae | 11.00 | |||
| Arctium sp. | Aphis fabae | 235.33 | 790.00 | ||
| Arctium minus | Aphis fabae | 246.34 | |||
| Artemisia vulgaris | Macrosiphoniella artemisiae | 171.34 | |||
| Berberis vulgaris | Liosomaphis berberidis | 70.67 | 1.00 | ||
| Betula pendula | Euceraphis punctipennis | 24.00 | 132.33 | 91.66 | |
| Carpinus betulus | Myzocallis carpini | 162.68 | |||
| Cerasus avium | Myzus cerasi | 972.99 | |||
| Cerasus vulgaris | Mysus cerasi | 1645.68 | |||
| Cicorium intybus | Uroleucon cichorii | 383.33 | 33.33 | ||
| Cirsium arvense | Aphis fabae | 76.67 | 83.67 | ||
| Cornus alba | Anoecia corni | 186.34 | |||
| Cornus sanguinea | Anoecia corni | 128.66 | 48.33 | ||
| Corylus avellana | Myzocallis coryli | 74.32 | |||
| Cotoneaster lucidus | Aphis pomi | 174.29 | 43.66 | ||
| Crataegus monogyna | Aphis pomi | 146.67 | |||
| Crataegus sp. | Prociphilus pini | 58.67 | |||
| Euonymus europaea | Aphis fabae | 119.66 | |||
| Fagus sylvatica | Phyllaphis fagi | 118.00 | |||
| Galium aparine | Myzus cerasi | 63.33 | |||
| Geum urbanum | Macrosiphum gei | 40.00 | |||
| Impatiens sp. | Impatientinum balsamines | 13.00 | |||
| Juglans regia | Chromaphis juglandicola | 57.34 | 25.00 | ||
| Lonicera xylosteum | Rhopalomyzus lonicerae | 139.67 | 133.33 | ||
| Lapsana communis | Uroleucon cichorii | 26.67 | |||
| Malus domestica | Aphis pomi | 22.33 | 84.00 | ||
| Rhopalosiphum insertum | 24.66 | 22.33 | 7.00 | ||
| Malus sylvestris | Aphis pomi | 5.66 | |||
| Dysaphis plantaginea | 1.67 | ||||
| Silene latifolia | Brachycaudus lychnidis | 34.00 | |||
| Philadelphus inodorus | Aphis fabae | 689.00 | 649.34 | ||
| Pinus nigra | Cinara pini | 201.00 | |||
| Pinus mugo | Cinara piceae | 73.34 | |||
| Pinus sylvestris | Cinara pinea | 63.00 | |||
| Populus alba | Chaitophorus populialbae | 120.10 | 193.67 | ||
| Populus tremula | Pachypappa tremulae | 113.68 | |||
| Chaitophorus populialbae | 27.33 | 66.00 | |||
| Populus nigra | Chaitophorus leucomelas | 187.33 | |||
| Prunus cerasifera | Hyalopterus pruni | 702.09 | 83.67 | 329.34 | |
| Rhopalosiphum nymphaeae | 16.33 | 188.34 | 78.01 | 425.64 | |
| Prunus domestica | Brachycaudus cardui | 334.67 | 390.00 | ||
| Hyalopterus pruni | 139.33 | 693.33 | 300.33 | 153.01 | |
| Phorodon humuli | 22.67 | ||||
| Rhopalosiphum nymphaeae | 470.99 | 72.66 | |||
| Phragmites australis | Hyalopterus pruni | 2296.33 | 4016.65 | ||
| Rhamnus cathartica | Aphis nasturtii | 25.76 | 69.66 | ||
| Ribes aureum | Aphis schneideri | 619.00 | |||
| Robinia pseudoacacia | Aphis craccivora | 2.67 | 141.67 | 4.33 | 377.00 |
| Appendiseta robiniae | 3.67 | 11.67 | 1.00 | ||
| Rosa rugosa | Macrosiphum rosae | 1261.00 | |||
| Metopolophium dirhodum | 64.66 | ||||
| Chaetosiphon tetrarhodum | 180.00 | 236.66 | |||
| Rosa canina | Metopolophium dirhodum | 6.33 | |||
| Rumex acetosa | Aphis fabae | 299.99 | |||
| Rumex crispus | Aphis fabae | ||||
| Rumex hydrolapathum | Aphis fabae | 23.33 | |||
| Qercus robur | Tuberculatus annulatus | 15.67 | 83.65 | 48.67 | |
| Qercus rubra | Tuberculatus annulatus | 157.99 | 274.34 | ||
| Salix fragilis | Chaitophorus salijaponicus | 59.00 | |||
| Salix xsepulcralis | Chaitophorus salijaponicus | 126.33 | |||
| Salix viminalis | Chaitophorus horii | 31.67 | |||
| Chaitophorus salijaponicus | 205.32 | ||||
| Pterocomma salicis | 1344.67 | ||||
| Sambucus nigra | Aphis sambuci | 2682.33 | 499.99 | 1183.35 | |
| Sonchus asper | Aphis craccivora | 37.33 | |||
| Hyperomyzus lactucae | 129.33 | ||||
| Sonchus arvensis | Aphis craccivora | 10.00 | |||
| Hyperomyzus lactucae | 60.00 | ||||
| Sorbus intermedia | Aphis pomi | 117.33 | |||
| Rhopalosiphum insertum | 1.67 | 41.34 | |||
| Sonchus oleraceus | Hyperomyzus lactucae | 91.67 | |||
| Spiraea sp. | Acyrthosiphon ignotum | 11.00 | |||
| Aphis spiraephaga | 616.34 | 890.33 | |||
| Tilia cordata | Eucallipterus tiliae | 0.33 | 175.01 | 162.67 | |
| Tilia xEuropaea | Eucallipterus tiliae | 99.99 | 314.65 | 148.02 | |
| Tilia „Euchlora” | Eucallipterus tiliae | 197.66 | |||
| Tilia platyphyllos | Eucallipterus tiliae | 7.33 | |||
| Tilia tomentosa | Eucallipterus tiliae | 58.00 | 50.33 | ||
| Ulmus glabra | Eriosoma ulmi | 379.01 | |||
| Tinocallis platani | 285.66 | ||||
| Ulmus laevis | Tetraneura ulmi | 43.00 | 897.68 | ||
| Tinocallis platani | 208.00 | ||||
| Ulmus minor | Tetraneura ulmi | 524.00 | |||
| Eriosoma ulmi | 40.00 | ||||
| Urtica dioica | Aphis urticata | 140.00 | 302.34 | ||
| Microlophium carnosum | 647.67 | ||||
| Viburnum opulus | Aphis fabae | 398.67 | |||
| 2012 | 2013 | 2014 | 2012–2014 | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Parks | P1 | P2 | P3 | P4 | P1 | P2 | P3 | P4 | P1 | P2 | P3 | P4 | P1 | P2 | P3 | P4 | ||
| Mean number of specimens | 3225.34 | 8220.65 | 2425.50 | 6242.00 | 984.99 | 2290.32 | 1246.35 | 3054.37 | 3143.67 | 3820.35 | 1698.62 | 5852.33 | 7354.00 | 14,331.32 | 5370.47 | 15,148.70 | ||
| Number of species | 17 | 19 | 18 | 19 | 15 | 15 | 16 | 17 | 21 | 16 | 19 | 24 | 25 | 28 | 28 | 33 | ||
| H’ | 3.427 | 3.192 | 3.550 | 3.095 | 3.0558 | 3.050 | 2.960 | 3.052 | 3.511 | 3.350 | 3.361 | 3.108 | 3.873 | 3.575 | 3.864 | 3.648 | ||
| P3*-P4 | P1*-P4 | |||||||||||||||||
| Re (%) | P1-P4 | P2-P4 | P3-P4 | No similarities | No similarities | P2-P4 | ||||||||||||
| 56.2 | 50.9 | 59.6 | 51.9 | |||||||||||||||
| 2012 | 2013 | 2014 | 2012–2014 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Aphid Species | P1 | P2 | P3 | P4 | P1 | P2 | P3 | P4 | P1 | P2 | P3 | P4 | P1 | P2 | P3 | P4 |
| Aphis craccivora | D3 | |||||||||||||||
| Aphis fabae | D3 | D3 | D3 | D3 | ||||||||||||
| Aphis pomi | D3 | |||||||||||||||
| Aphis sambuci | D4 | D4 | D3 | D3 | D3 | |||||||||||
| Aphis spiraephaga | D4 | D3 | ||||||||||||||
| Brachycaudus cardui | D4 | D3 | ||||||||||||||
| Drepanosiphum acerinum | D3 | |||||||||||||||
| Eucallipterus tilliae | D3 | D4 | D3 | D3 | ||||||||||||
| Hyalopterus pruni | D3 | D4 | D3 | D4 | D3 | D4 | D3 | D4 | D3 | D4 | D4 | |||||
| Macrosiphum rosae | D3 | D4 | D3 | |||||||||||||
| Myzus cerasi | D3 | D3 | D4 | D3 | D4 | D3 | D3 | |||||||||
| Periphyllus testudinaceus | D4 | D4 | D4 | D3 | D3 | D3 | ||||||||||
| Pterocomma salicis | D4 | |||||||||||||||
| Tinocallis platani | D3 | |||||||||||||||
| Tuberculatus annulatus | D3 | |||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barczak, T.; Bennewicz, J.; Korczyński, M.; Błażejewicz-Zawadzińska, M.; Piekarska-Boniecka, H. Aphid Assemblages Associated with Urban Park Plant Communities. Insects 2021, 12, 173. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020173
Barczak T, Bennewicz J, Korczyński M, Błażejewicz-Zawadzińska M, Piekarska-Boniecka H. Aphid Assemblages Associated with Urban Park Plant Communities. Insects. 2021; 12(2):173. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020173
Chicago/Turabian StyleBarczak, Tadeusz, Janina Bennewicz, Maciej Korczyński, Małgorzata Błażejewicz-Zawadzińska, and Hanna Piekarska-Boniecka. 2021. "Aphid Assemblages Associated with Urban Park Plant Communities" Insects 12, no. 2: 173. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020173