
Effects of Rearing Density on Developmental Traits of Two Different Biotypes of the Gypsy Moth, Lymantria Dispar L., from China and the USA


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Gypsy Moth Populations
2.2. Diet and Rearing Conditions
2.3. Rearing Gypsy Moth at Different Densities
2.4. Statistical Analysis
3. Results
3.1. Impact on Larval Development Time and Survival
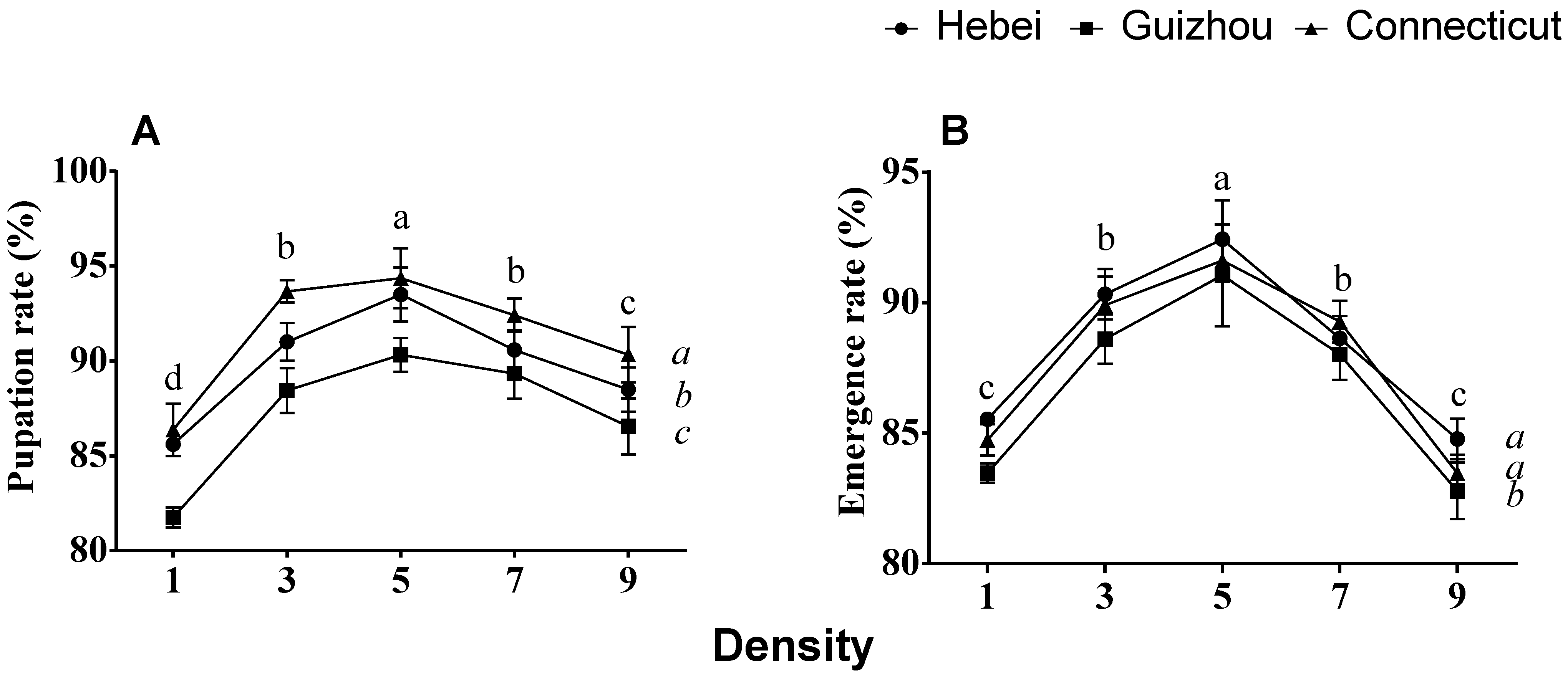
3.2. Impact on Pupation and Emergence
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Glossary of Phytosanitary Terms; International Standard for Phytosanitary Measures No. 5; FAO on behalf of the Secretariat of the International Plant Protection Convention (IPPC): Rome, Italy, 2019. [Google Scholar]
- Holmes, T.P.; Aukema, J.E.; Von Holle, B.; Liebhold, A.; Sills, E. Economic impacts of invasive species in forests: Past, present, and future. Ann. N. Y. Acad. Sci. 2009, 1162, 18–38. [Google Scholar] [CrossRef] [PubMed]
- Aukema, J.E.; Leung, B.; Kovacs, K.; Chivers, C.; Britton, K.O.; Englin, J.; Frankel, S.J.; Haight, R.G.; Holmes, T.P.; Liebhold, A.M.; et al. Economic impacts of non-native forest insects in the continental United States. PLoS ONE 2011, 6, e24587. [Google Scholar] [CrossRef]
- Keena, M.A.; Cote, M.J.; Grinberg, P.S.; Wallner, W.E. World distribution of female flight and genetic variation in Lymantria dispar (Lepidoptera: Lymantriidae). Environ. Entomol. 2008, 37, 636–649. [Google Scholar] [CrossRef] [Green Version]
- Keena, M.A.; Richards, J.Y. Comparison of survival and sevelopment of gypsy moth Lymantria dispar L. (Lepidoptera: Erebidae) populations from different geographic areas on North American conifers. Insects 2020, 11, 260. [Google Scholar] [CrossRef]
- Peterson, A.T.; Williams, R.; Chen, G. Modeled global invasive potential of Asian gypsy moths, Lymantria dispar. Entomol. Exp. Appl. 2007, 125, 39–44. [Google Scholar] [CrossRef]
- USDA/APHIS/PPQ. Asian Gypsy Moth Survey and Response Guidelines; USDA: Washington, DC, USA, 2014. [Google Scholar]
- Campbell, R.W.; Sloan, R.J. Numerical bimodality among North American gypsy moth populations. Environ. Entomol. 1978, 7, 641–646. [Google Scholar] [CrossRef]
- Schaefer, P.W.; Weseloh, R.M.; Sun, X.; Wallner, W.E.; Yan, J. Gypsy moth, Lymantria (=Ocneria) dispar (L.) (Lepidoptera: Lymantriidae), in the People’s Republic of China. Environ. Entomol. 1984, 13, 1535–1541. [Google Scholar] [CrossRef]
- Walsh, P.J. Asian gypsy moth: The risk to New Zealand. N. Z. For. 1993, 38, 41–43. [Google Scholar]
- Alalouni, U.; Schadler, M.; Brandl, R. Natural enemies and environmental factors affecting the population dynamics of the gypsy moth. J. Appl. Entomol. 2013, 137, 721–738. [Google Scholar] [CrossRef]
- Peters, T.M.; Barbosa, P. Influence of population density on size, fecundity, and developmental rate of insects in culture. Annu. Rev. Entomol. 1977, 22, 431–450. [Google Scholar] [CrossRef]
- Than, A.T.; Ponton, F.; Morimoto, J. Integrative developmental ecology: A review of density-dependent effects on life-history traits and host-microbe interactions in non-social holometabolous insects. Evol. Ecol. 2020, 34, 659–680. [Google Scholar] [CrossRef]
- Bauerfeind, S.S.; Fischer, K. Effects of food stress and density in different life stages on reproduction in a butterfly. Oikos 2005, 111, 514–524. [Google Scholar] [CrossRef]
- Leonard, D.E. Effects of density of larvae on the biology of the gypsy moth, Porthretria dispar. Entomol. Exp. Appl. 1968, 11, 291–304. [Google Scholar] [CrossRef]
- Lance, D.R.; Elkinton, J.S.; Schawlbe, C.P. Components of density-related stress as potential determinants of population quality in the gypsy moth (Lepidoptera: Lymantriidae). Environ. Entomol. 1986, 15, 914–918. [Google Scholar] [CrossRef]
- Lazarevic, J.; Peric-Mataruga, V.; Vlahovic, M.; Mrdakovic, M.; Cvetanovic, D. Effects of rearing density on larval growth and activity of digestive enzymes in Lymantria dispar L. (Lepidoptera: Lymantriidae). Folia Biol. 2004, 52, 105–112. [Google Scholar]
- Pavlushin, S.V.; Belousova, I.A.; Chertkova, E.A.; Kryukova, N.A.; Glupov, V.V.; Martemyanov, V.V. The effect of population density of Lymantria dispar (Lepidoptera: Erebidae) on its fitness, physiology and activation of the covert nucleopolyhedrovirus. Eur. J. Entomol. 2019, 116, 85–91. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, S.; Wang, H.; Wang, M.; Li, G. Mitochondrial gene sequence (COI) reveals the genetic structure and demographic history of Lymantria dispar (Lepidoptera: Erebidae: Lymantriinae) in and around China. Insects 2019, 10, 146. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Wu, Y.; Kurenshchikov, D.K.; Ilyinykh, A.V.; Shi, J. Underestimated mitochondrial diversity in gypsy moth Lymantria dispar from Asia. Agric. For. Entomol. 2019, 21, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Zuo, Y.; Kurenshchikov, D.K.; Yu, J.; Zou, Y.; Wang, Y.; Wang, Y.; Shi, J. Microsatellite and morphological analyses reveal unexpected diversity in Lymantria dispar in China. Forests 2019, 10, 1100. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.J.; Yang, F.; Tang, S.Y.; Chen, M. Development of an artificial diet for three lepidopteran insects. Chin. J. Appl. Entomol. 2014, 51, 1376–1386. [Google Scholar]
- Campbell, R.W. Some effects of gypsy moth density on rate of development, pupation time, and fecundity. Ann. Entomol. Soc. Am. 1978, 71, 442–448. [Google Scholar] [CrossRef]
- Goulson, D.; Cory, J.S. Responses of Mamestra brassicae (Lepidoptera: Noctuidae) to crowding: Interactions with disease resistance, colour phase and growth. Oecologia 1995, 104, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Juan, W.; Jiang, X.-F.; Wu, D.-L.; Luo, L.-Z. Effect of larval rearing density on development and fecundity of the beet armyworm, Spodoptera exigua (Lepidoptera: Noctuidae). Acta Entomol. Sin. 2008, 51, 889–894. [Google Scholar]
- Myers, J.H.; Cory, J.S. Ecology and evolution of pathogens in natural populations of Lepidoptera. Evol. Appl. 2016, 9, 231–247. [Google Scholar] [CrossRef] [Green Version]
- Reilly, J.R.; Hajek, A.E. Density-dependent resistance of the gypsy moth Lymantria dispar to its nucleopolyhedrovirus, and the consequences for population dynamics. Oecologia 2008, 154, 691–701. [Google Scholar] [CrossRef]
- Fuxa, J.R.; Sun, J.Z.; Weidner, E.H.; LaMotte, L.R. Stressors and rearing diseases of Trichoplusia ni: Evidence of vertical transmission of NPV and CPV. J. Invertebr. Pathol. 1999, 74, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Opoku-Debrah, J.K.; Hill, M.P.; Knox, C.; Moore, S.D. Overcrowding of false codling moth, Thaumatotibia leucotreta (Meyrick) leads to the isolation of five new Cryptophlebia leucotreta granulovirus (CrleGV-SA) isolates. J. Invertebr. Pathol. 2013, 112, 219–228. [Google Scholar] [CrossRef] [Green Version]
- Gruntenko, N.E.; Rauschenbach, I.Y. The role of insulin signalling in the endocrine stress response in Drosophila melanogaster: A mini-review. Gen. Comp. Endocrinol. 2018, 258, 134–139. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Parameters | Factor | Two-Way ANOVA | ||
|---|---|---|---|---|
| df | F ( DFn, DFd ) | p-Value | ||
| Larval duration (d) | Larval density (n/cup) | 4 | F (4, 30) = 20.73 | <0.0001 |
| Population | 2 | F (2, 30) = 14.99 | <0.0001 | |
| Larval density (n/cup) × population | 8 | F (8, 30) = 0.79 | =0.6177 | |
| Larval survival rate (%) | Larval density (n/cup) | 4 | F (4, 30) = 109.1 | <0.0001 |
| Population | 2 | F (2, 30) = 53.62 | <0.0001 | |
| Larval density (n/cup) × population | 8 | F (8, 30) = 1.123 | =0.3767 | |
| Pupation rate (%) | Larval density (n/cup) | 4 | F (4, 30) = 68.50 | <0.0001 |
| Population | 2 | F (2, 30) = 50.39 | <0.0001 | |
| Larval density (n/cup) × population | 8 | F (8, 30) = 0.89 | =0.5313 | |
| Emergence rate (%) | Larval density (n/cup) | 4 | F (4, 30) = 106.2 | <0.0001 |
| Population | 2 | F (2, 30) = 9.507 | <0.0001 | |
| Larval density (n/cup) × population | 8 | F (8, 30) = 0.52 | =0.8332 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Harrison, R.L.; Shi, J. Effects of Rearing Density on Developmental Traits of Two Different Biotypes of the Gypsy Moth, Lymantria Dispar L., from China and the USA. Insects 2021, 12, 175. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020175
Wang Y, Harrison RL, Shi J. Effects of Rearing Density on Developmental Traits of Two Different Biotypes of the Gypsy Moth, Lymantria Dispar L., from China and the USA. Insects. 2021; 12(2):175. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020175
Chicago/Turabian StyleWang, Yiming, Robert L. Harrison, and Juan Shi. 2021. "Effects of Rearing Density on Developmental Traits of Two Different Biotypes of the Gypsy Moth, Lymantria Dispar L., from China and the USA" Insects 12, no. 2: 175. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020175