Molecular Identification and Immunity Functional Characterization of Lmserpin1 in Locusta migratoria manilensis

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Expression Analysis of Lmserpin1 Gene in Different Tissues and Developmental Stages
2.3. Cloning of the Lmserpin1 Gene and RNA Interference
2.4. Expression Analysis of Lmserpin1 after Metarhizium Anisopliae Infection
2.5. Bioassay
2.6. Enzyme Activity Assay
2.7. Quantitative Analysis of Immune Related Gene Expression
2.8. Statistical Analysis
3. Result
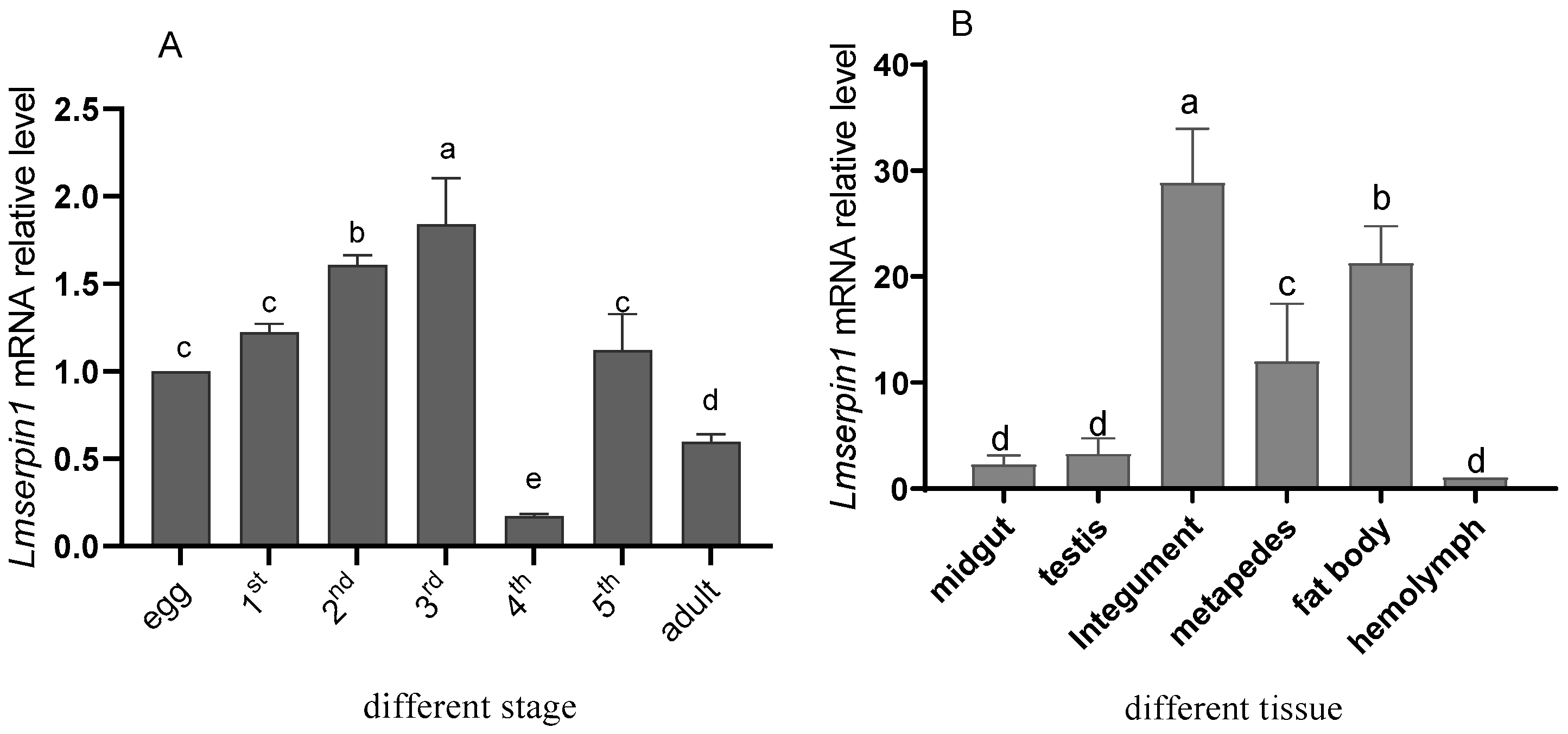
3.1. Expression of Lmserpin1 in Different Tissues and Developmental Stages of L. migratoria manilensis
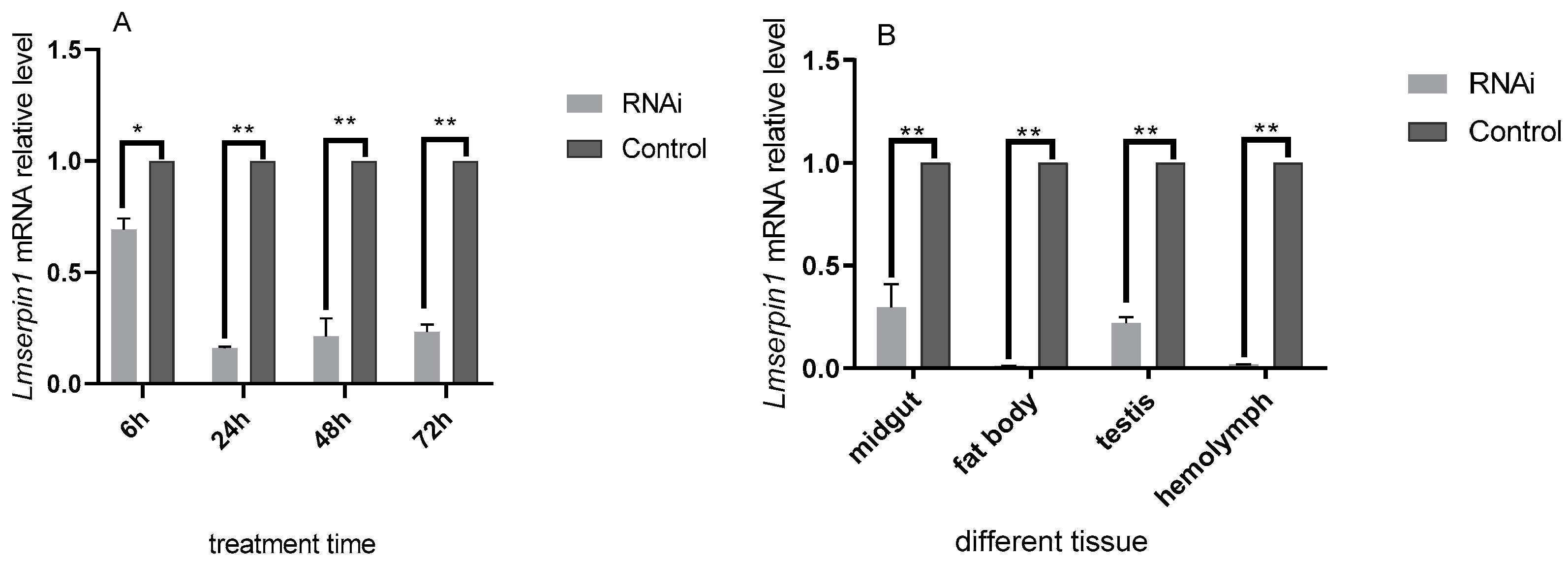
3.2. The Expression of Lmserpin1 of L. migratoria manilensis in 3rd Instars after RNAi
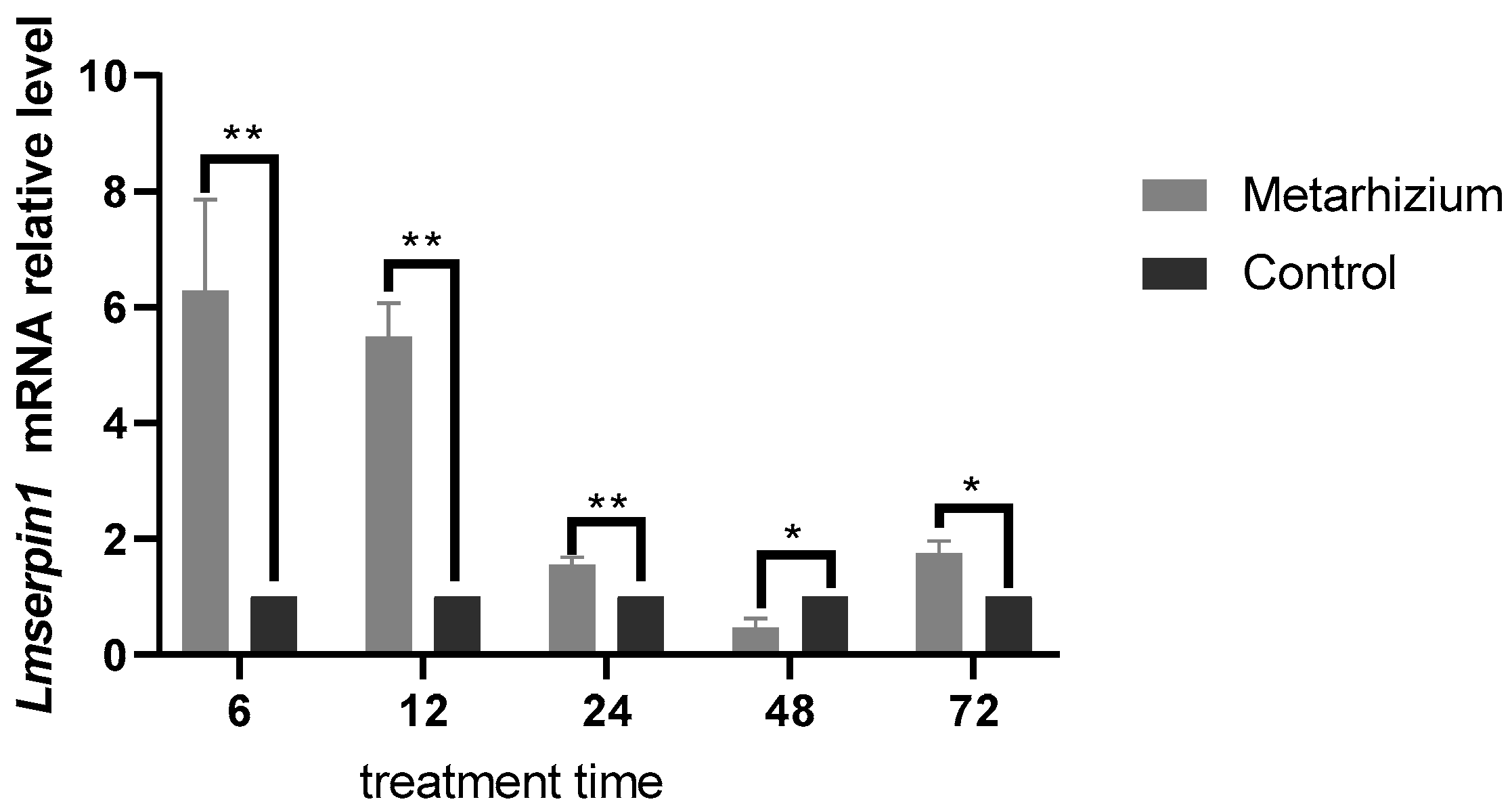
3.3. The Expression of Lmserpin1 with Application of M. anisopliae
3.4. Mortality Comparison under Different Bioassay Treatments
3.5. The Enzyme Activity of L. migratoria manilensis among Different Treatment Groups
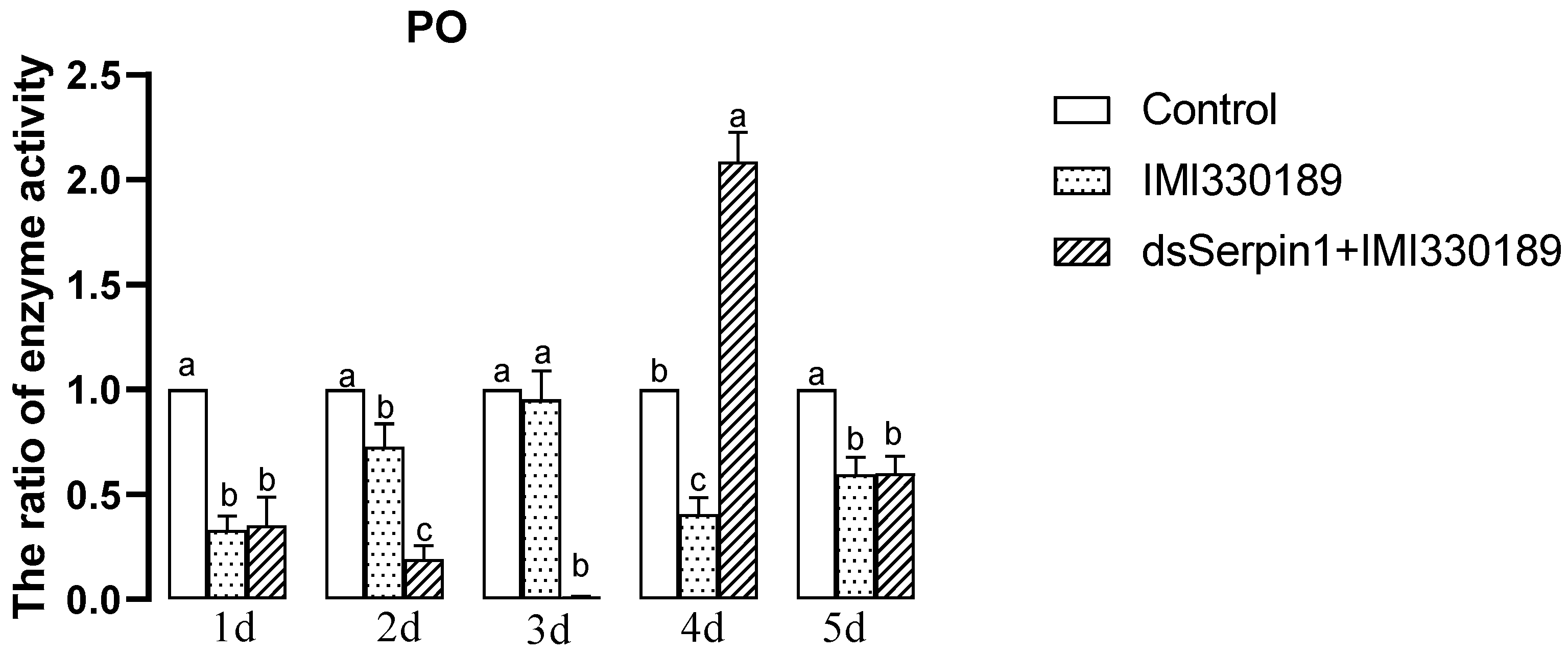
3.5.1. Effects of Different Treatment Groups on PO Activity of L. migratoria manilensis
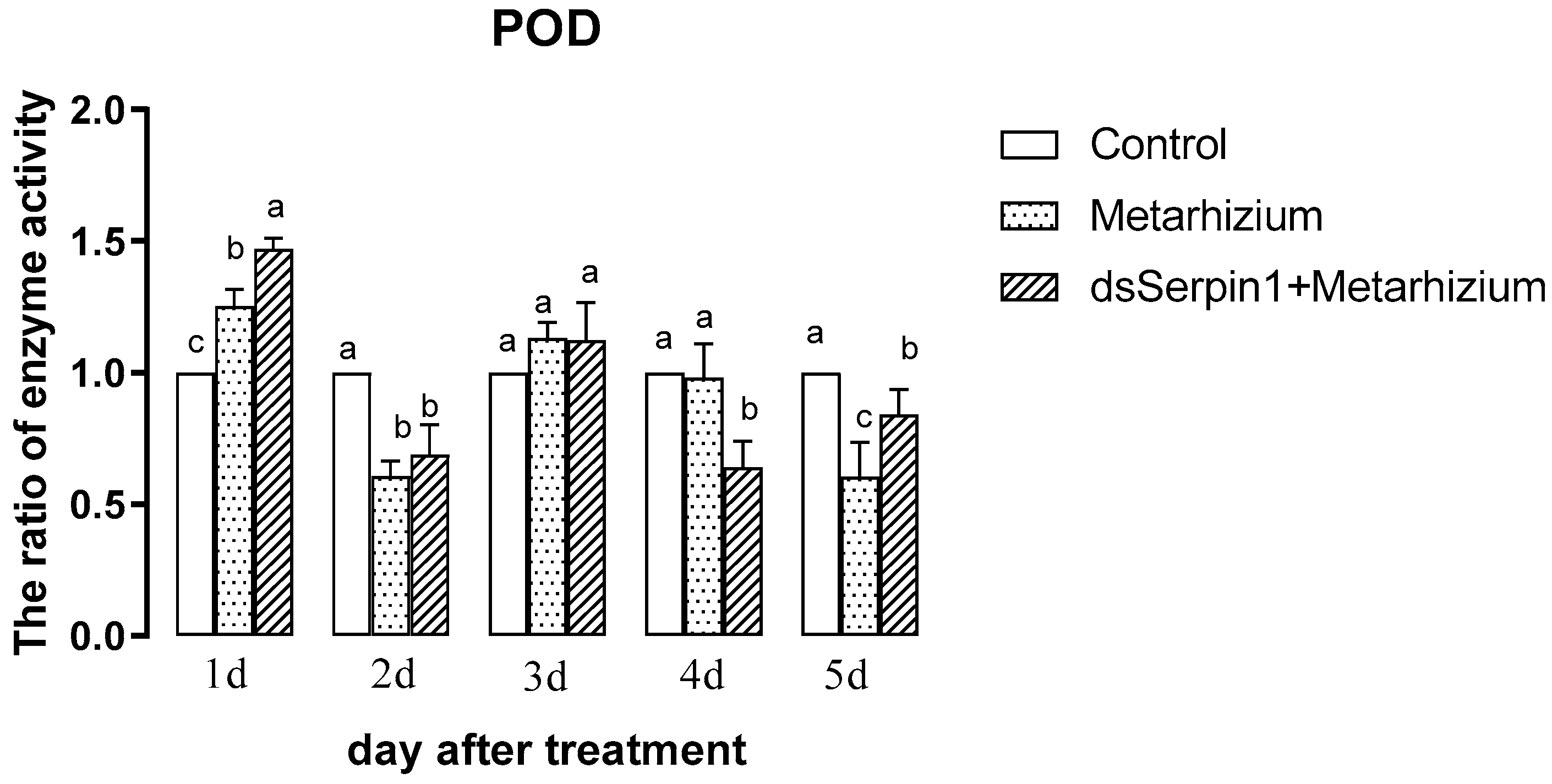
3.5.2. Effects of Different Treatment Groups on POD Activity of L. migratoria manilensis
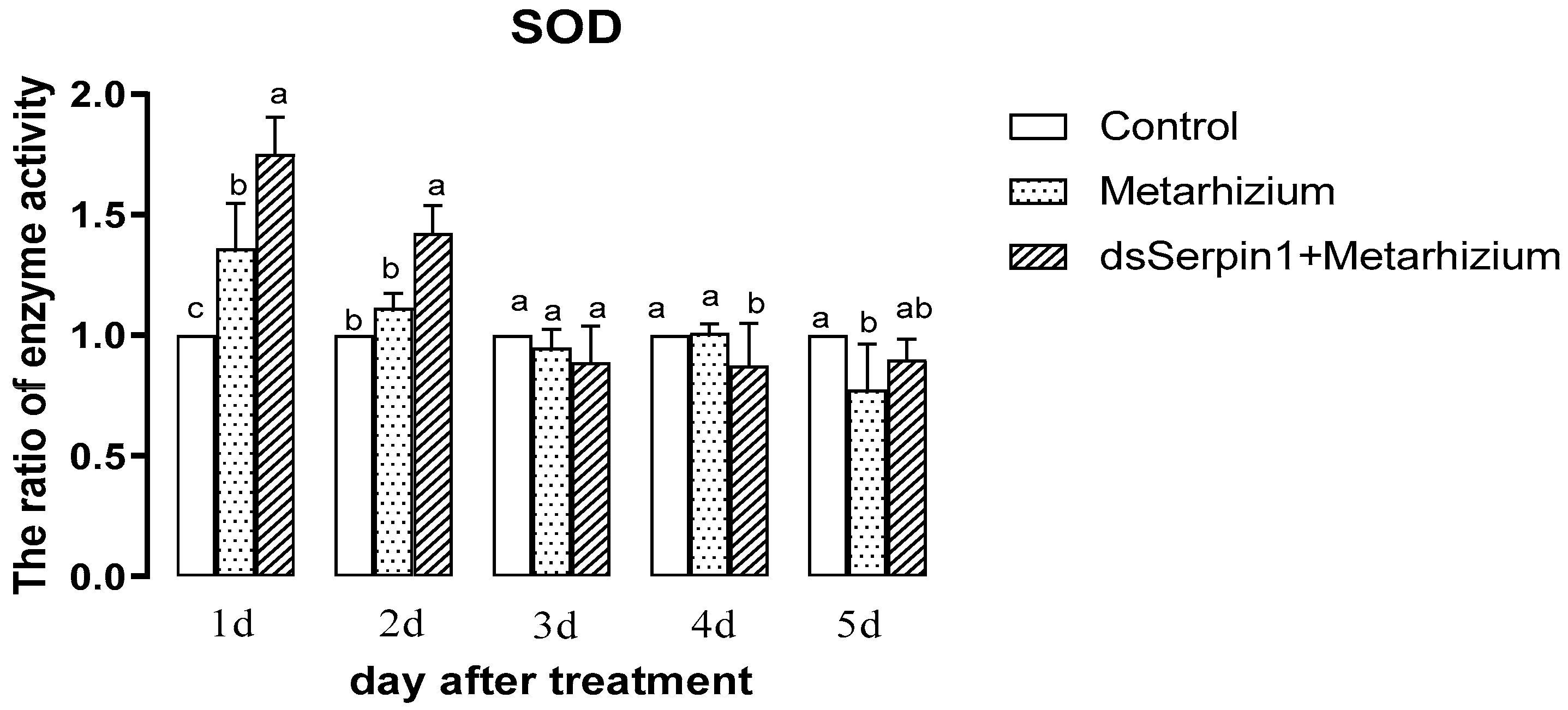
3.5.3. Effects of Different Treatment Groups on SOD Activity of L. migratoria manilensis
3.5.4. Effects of Different Treatment Groups on MFO Activity of L. migratoria manilensis
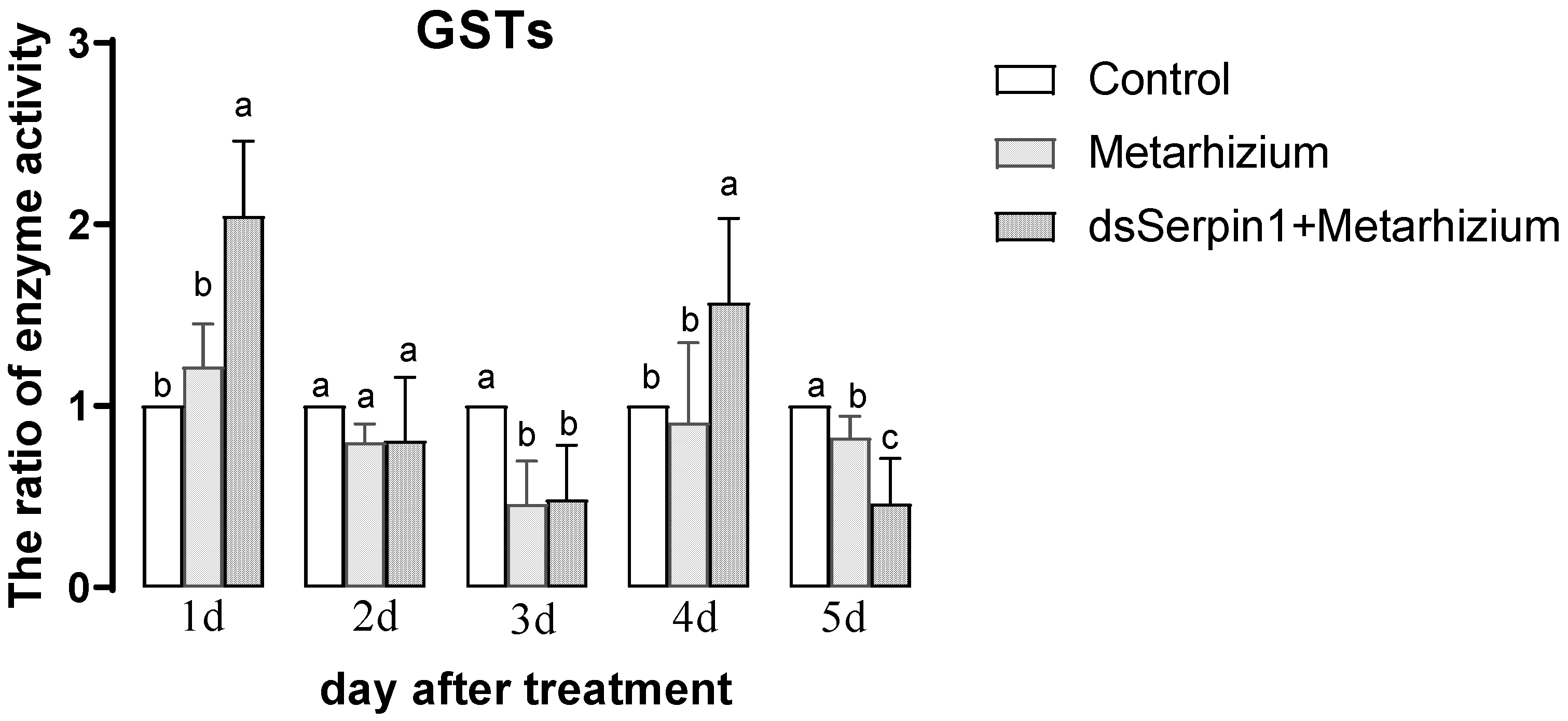
3.5.5. Effects of Different Treatment Groups on GSTs Activity of L. migratoria manilensis
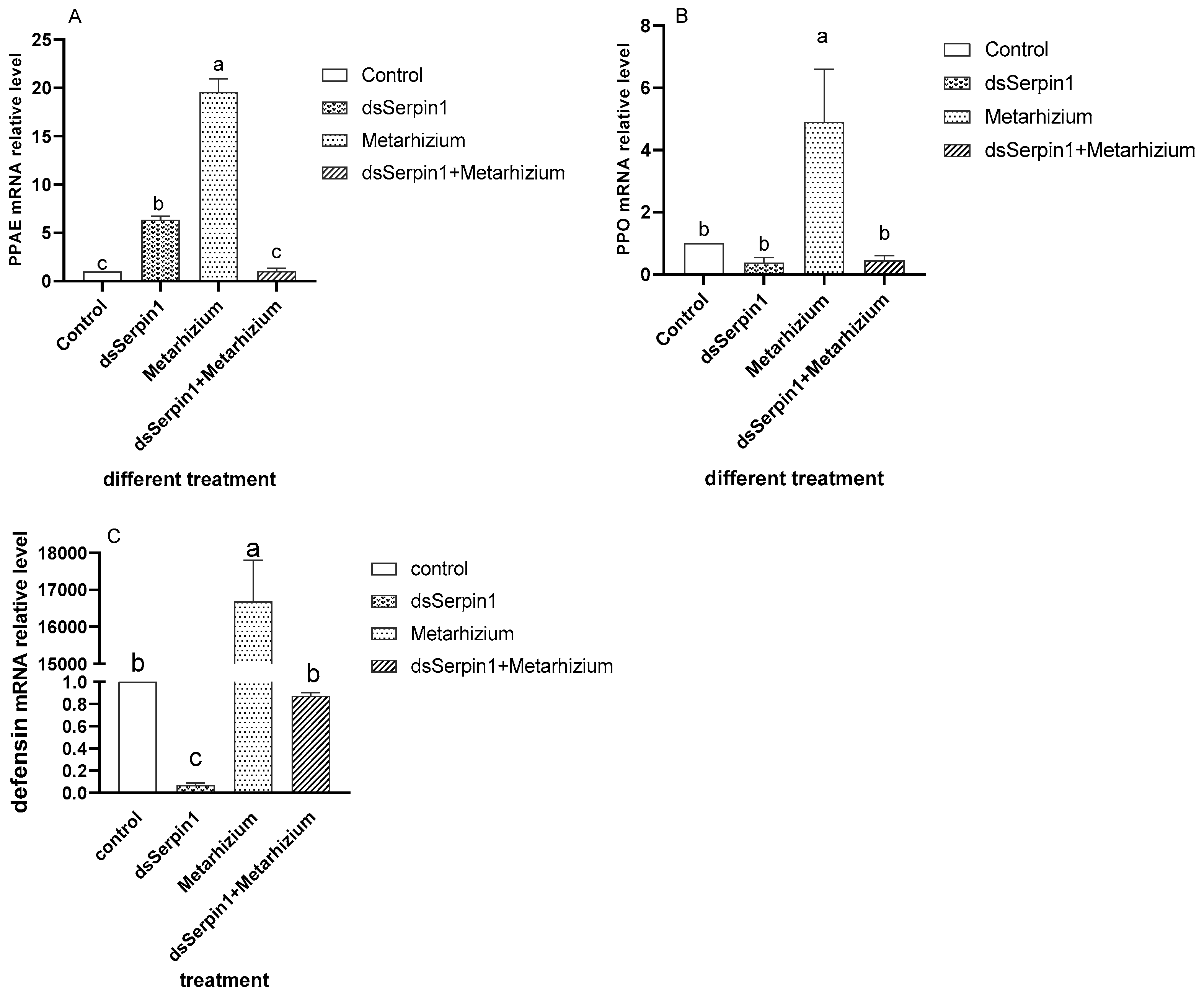
3.6. Effect of RNAi on mRNA Levels of Immune Related Pathway Gene
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Celli, J.; Finlay, B.B. Bacterial avoidance of phagocytosis. Trends. Microbiol. 2002, 10, 232–237. [Google Scholar] [CrossRef]
- Nielsen-LeRoux, C.; Gaudriault, S.; Ramarao, N.; Lereclus, D.; Givau-dan, A. How the insect pathogen bacteria Bacillus thuringiensis and Xenorhabdus/Photorhabdus occupy their hosts. Curr. Opin. Microbiol. 2012, 15, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Hultmark, D.; Engström, A.; Andersson, K.; Steiner, H.; Bennich, H.; Boman, H.G. Insect immunity. attacins, a family of antibacterial proteins from hyalophora cecropia. Embo. J. 1983, 2, 571–576. [Google Scholar] [CrossRef]
- Iwona, W.; Małgorzata, C.; Agnieszka, Z.B.; Jakub, K. Insect Defense Proteins and Peptides. Subcell. Biochem. 2020, 94, 81–121. [Google Scholar]
- Cerenius, L.; Söderhäll, K. The prophenoloxidase-activating system in invertebrates. Immunol. Rev. 2004, 198, 116–126. [Google Scholar] [CrossRef]
- Jiravanichpaisal, P.; Lee, B.L.; Söderhäll, K. Cell-mediated immunity in arthropods: Hematopoiesis, coagulation, melanization and opsonization. Immunobiology 2006, 211, 213–236. [Google Scholar] [CrossRef] [PubMed]
- Olson, S.T.; Richard, B.; Izaguirre, G.; Schedin-Weiss, S.; Gettins, P.G. Molecular mechanisms of antithrombin-heparin regulation of blood clotting proteinases. A paradigm for understanding proteinase regulation by serpin family proteinproteinase inhibitors. Biochimie 2010, 92, 1587–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemaitre, B.; Hoffmann, J. The host defense of Drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697–743. [Google Scholar] [CrossRef] [Green Version]
- Satoh, D.; Horii, A.; Ochiai, M.; Ashida, M. Prophenoloxidase-activating enzyme of the silkworm, Bombyx mori. Purification, characterization, and cDNA cloning. J. Biol. Chem. 1999, 274, 7441–7453. [Google Scholar] [CrossRef] [Green Version]
- Kanost, M.R. Serine proteinase inhibitors in arthropod immunity. Dev. Comp. Immunol. 1999, 23, 291–301. [Google Scholar] [CrossRef]
- Silverman, G.A.; Bird, P.L.; Carrell, R.W.; Church, F.C.; Coughlin, P.B.; Gettins, P.G. The serpins are an expanding uperfamily of structurally similar but functionally diverse proteins. Evolution mechanism of inhibition, novel functions, and a revised nomenclature. J. Biol. Chem. 2001, 276, 33293–33296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Law, R.H.; Zhang, Q.; Mcgowan, S.; Buckle, A.M.; Silverman, G.A.; Wong, W.; Rosado, C.J.; Langendorf, C.G.; Pike, R.N.; Bird, P.I.; et al. An overview of the serpin superfamily. Genome Biol. 2006, 7, 216. [Google Scholar] [CrossRef] [Green Version]
- Laskowski, M.; Kato, I. Protein inhibitors of proteinases. Annu. Rev. Biochem. 1980, 49, 593–626. [Google Scholar] [CrossRef] [PubMed]
- Gregorio, E.D.; Han, S.J.; Lee, W.J.; Baek, M.J.; Osaki, T.; Kawabata, S.I.; Lee, B.L.; Iwanaga, S.; Lemaitre, B.; Brey, P.T. An immune-responsive serpin regulates the melanization cascade in drosophila. Dev. Cell. 2002, 3, 581–592. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Kanost, M.R. Characterization and functional analysis of 12 naturally occurring reactive site variants of serpin-1 from Manduca sexta. J. Biol. Chem. 1997, 272, 1082–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, P.; Dong, Z.; Duan, J.; Wang, G.; Wang, L.; Li, Y.; Xiang, Z.; Xia, Q. Genome-wide identification and immune response analysis of serine protease inhibitor genes in the silkworm, Bombyx mori. PLoS ONE 2012, 7, e31168. [Google Scholar] [CrossRef]
- Zheng, Y.P.; He, W.Y.; Béliveau, C.; Nisole, A.; Stewart, D.; Zheng, S.C.; Douceta, D.; Cusson, M.; Fengb, Q.L. Cloning, expression and characterization of four serpin-1 cDNA variants from the spruce budworm, Choristoneura fumiferana. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2009, 154, 165–173. [Google Scholar] [CrossRef]
- Iwanaga, S.; Kawabata, S.; Muta, T. New types of clotting factors and defense molecules found in horseshoe crab hemolymph: Their structures and functions. J. Biochem. 1998, 123, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.Z.; Keyhani, N.O.; Tang, G.; Tian, C.; Fan, Y.H. Expression of a toll signaling regulator serpin in a mycoinsecticide for increased virulence. Appl. Environ. Microb. 2014, 80, 4531–4539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, Y.; Kanost, M.R. Manduca sexta serpin-4 and serpin-5 inhibit the prophenoloxidase activation pathway: cDNA cloning, protein expression, and characterization. J. Biol. Chem. 2005, 280, 14923–14931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, C.; Jiang, H.; Kanost, M.R. Proteolytic activation and function of the cytokine Spätzle in the innate immune response of a lepidopteran insect, Manduca sexta. FEBS J. 2010, 277, 148–162. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Davids, J.A.; Zheng, D.; Bryant, M.; Bot, I.; Theo, J.C.V.B.; Biessen, E.; Pepine, C.; Ryman, K.; Progulski-Fox, A.; et al. The serpin solution; targeting thrombotic and thrombolytic serine proteases in inflammation. Cardiovasc. Hematol. Disord. Drug Targets 2013, 13, 99–110. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, Y.; Gorman, M.J. Manduca sexta serpin-3 regulates prophenoloxidase activation in response to infection by inhibiting prophenoloxidase-activating proteinases. J. Biol. Chem. 2003, 278, 46556–46564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, Y.; Kanost, H.J.; Identiiang, M. Identification of plasma proteases inhibited by Manduca sexta serpin-4 and serpin-5 and their association with components of the prophenoloxidase activation pathway. J. Biol. Chem. 2005, 280, 14932–14942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nappi, A.J.; Frey, F.; Carton, Y. Drosophila serpin 27A is a likely target for immune suppression of the blood cell-mediated melanotic encapsulation response. J. Insect Physiol. 2005, 51, 197–205. [Google Scholar] [CrossRef]
- Zou, Z.; Picheng, Z.; Weng, H.; Mita, K.; Jiang, H. A comparative analysis of serpin genes in the silkworm genome. Genomics 2009, 93, 367–375. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.H.; Kim, S.J.; Kan, H.; Kwon, H.M.; Roh, K.B.; Jiang, R.; Park, J.W.; Lee, H.H.; Ha, N.C.; Kang, H.J.; et al. A three-step proteolytic cascade mediates the activation of the peptidoglycan-induced toll pathway in an insect. J. Biol. Chem. 2008, 283, 7599–7607. [Google Scholar] [CrossRef] [Green Version]
- Zhong, X.; Xu, X.X.; Yi, H.Y.; Lin, C.; Yu, X.Q. A toll-spatzle pathway in the tobacco hornworm, Manduca sexta. Insect Biochem. Mol. Biol. 2012, 42, 514–524. [Google Scholar] [CrossRef] [Green Version]
- COPR (Centre of Overseas Pest Research). The Locust and Grasshopper Agricultural Manual; ACIAR: Canberra, Australia, 1982; pp. 449–467.
- Cao, G.; Jia, M.; Zhao, X.; Wang, L.; Zhang, Z. Different effects of metarhizium anisopliae strains IMI330189 and IBC200614 on enzymes activities and hemocytes of Locusta migratoria L. PLoS ONE 2016, 11, e0155257. [Google Scholar] [CrossRef]
- Chen, J.; Cui, D.; Ullah, H.; Hao, K.; Tu, X.; Zhang, Z. Serpin7 controls egg diapause of migratory locust (locusta migratoria) by regulating polyphenol oxidase. FEBS Open Bio 2020, 10, 707–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowshani, A.T.; Strik, M.C.M.; Rosalie, M.; Si-La, Y.; Wolbink, A.M.; Bemelman, F.J. The granzyme b inhibitor serpinb9 (protease inhibitor 9) circulates in blood and increases on primary cytomegalovirus infection after renal transplantation. J. Infect. Dis. 2006, 192, 1908–1911. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2 −ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Gao, Q.; Yang, L.; Dai, J.; Yuan, G.; Wang, L.; Qian, C.; Zhu, B.; Liu, C.; Wei, G. Characterization and functional analysis of serpin-28 gene from silkworm, bombyx mori. J. Inverteb. Pathol. 2018, 159, 18–27. [Google Scholar] [CrossRef]
- Hao, K.; Ullah, H.; Jarwar, A.R.; Nong, X.; Tu, X.; Zhang, Z. Molecular identification and diapause-related functional characterization of a novel dual-specificity kinase gene, MPKL, in Locusta migratoria. FEBS Lett. 2019, 593, 3064–3074. [Google Scholar] [CrossRef]
- Mullen, L.M.; Goldsworthy, G.J. Immune responses of locusts to challenge with the pathogenic fungus Metarhizium or high doses of laminarin. J. Insect Physiol. 2006, 52, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.B.; Luo, W.C.; Jiang, L.; Xie, X.Y.; Xiao, T.; Yan, L. Inhibition kinetics of cabbage butterfly (Pieris rapae L.) larvae phenoloxidase activity by 3-hydroxy-4-methoxybenzaldehyde thiosemicarbazone. Appl. Biochem. Biotechnol. 2007, 143, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Moores, G.D.; Denholm, I.; Devonshire, A.L. Association between biochemical markers and insecticide resistance in the cotton aphid, aphis gossypiiglover. Pestic. Biochem. Phys. 1998, 62, 164–171. [Google Scholar] [CrossRef]
- Oppenoorth, F.J.; Welling, W. Biochemistry and physiology of resistance. In Wilkinson CF. Insecticide Biochemistry and Physiology; Springer: New York, NY, USA, 1976; pp. 507–551. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein–dye binding. Nature 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Shia, A.K.H.; Glittenberg, M.; Thompson, G.; Weber, A.N.; Reichhart, J.; Ligoxygakis, P. Toll-dependent antimicrobial responses in Drosophila larval fat body require Spätzle secreted by haemocytes. J. Cell Sci. 2009, 122, 4505–4515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Faria, M.R.; Wraight, S.P. Mycoinsecticides and mycoacaricides: A comprehensive list with worldwide coverage and international classification of formulation types. Biol. Control. 2007, 43, 237–256. [Google Scholar] [CrossRef]
- Abro, N.A.; Wang, G.; Ullah, H.; Long, G.L.; Hao, K.; Nong, X.; Cai, N.; Tu, X.; Zhang, Z. Influence of Metarhizium anisopliae (IMI330189) and Mad1 protein on enzymatic activities and Toll-related genes of migratory locust. Environ. Sci. Pollut. R. 2019, 26, 17797–17808. [Google Scholar] [CrossRef] [PubMed]
- Dan, S.; Akio, H.; Masanori, O.; Masaaki, A. Prophenoloxidase-activating enzyme of the silkworm, Bombyx mori Purification, characterization and cDNA cloning. J. Biol. Inorg. Chem. 2007, 274, 7441–7453. [Google Scholar]
- Duan, L.; Otvosi, S. Influence of larval age and virus concentration on mortality and sublethal effects of a nucleopolyhedrovirus on the west spruce budworm (Lepidoptera: Tortricidae). Environ. Entomol. 2001, 30, 136–146. [Google Scholar] [CrossRef]
- Zou, Z.; Jiang, H. Manduca sexta serpin-6 regulates immune serine proteinases PAP-3 and HP8. cDNA cloning, protein expression, inhibition kinetics, and function elucidation. J. Biol. Chem. 2005, 280, 14341–14348. [Google Scholar] [CrossRef] [Green Version]
- Somnuk, S.; Tassanakajon, A.; Rimphanitchayakit, V. Gene expression and characterization of a serine proteinase inhibitor pmserpin8 from the black tiger shrimp penaeus monodon. Fish Shellfish Immunol. 2012, 33, 332–341. [Google Scholar] [CrossRef]
- Wang, Z.H. Virulence of Metarhizium anisopliae extracellular proteinases in midgut of Locusta migratoria manilensis Meyen. Master’s Thesis, Gansu Agricultural University, Lanzhou, Gansu, 2016. (In Chinese). [Google Scholar]
- Chen, E.H.; Wei, D.; Wei, D.D.; Yuan, G.R.; Wang, J.J. The effect of dietary restriction on longevity, fecundity, and antioxidant responses in the oriental fruit fly, Bactrocera dorsalis (Hendel) (Diptera: Tephritidae). J. Insect Physiol. 2013, 59, 1008–1016. [Google Scholar] [CrossRef]
- Felton, G.W.; Summers, C.B. Antioxidant systems in insects. Arch. Insect Biochem. Physiol. 1995, 29, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Schuler, M.A.; Berenbaum, M.R. Molecular mechanisms of metabolic resistance to synthetic and natural xenobiotics. Annu. Rev. Entomol. 2007, 52, 231–253. [Google Scholar] [CrossRef]
- Feyereisen, R. Insect cytochrome P450. In Comprehensive Molecular Insect Science; Gilbert, L.I., Latrou, K., Gill, S.S., Eds.; Elsevier: Oxford, UK; pp. 1–77.
- Enayati, A.A.; Ranson, H.; Hemingway, J. Insect glutathione transferases and insecticide resistance. Insect Mol. Biol. 2005, 14, 3–8. [Google Scholar] [CrossRef] [Green Version]
- Lumjuan, N.; McCarroll, L.; Prapanthadara, L.; Hemingway, J.; Ranson, H. Elevated activity of an epsilon class glutathione transferase confers DDT resistance in the dengue vector, Aedes aegypti. Insect Biochem. Mol. Biol. 2005, 35, 861–871. [Google Scholar] [CrossRef]
- Liu, D.; Wang, L.; Yang, L.; Qian, C.; Wei, G.; Dai, L.; Li, J.; Zhu, B.; Liu, C. Serpin-15 from Bombyx mori inhibits prophenoloxidase activation and expression of antimicrobial peptides. Dev. Comp. Immunol. 2015, 51, 22–28. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
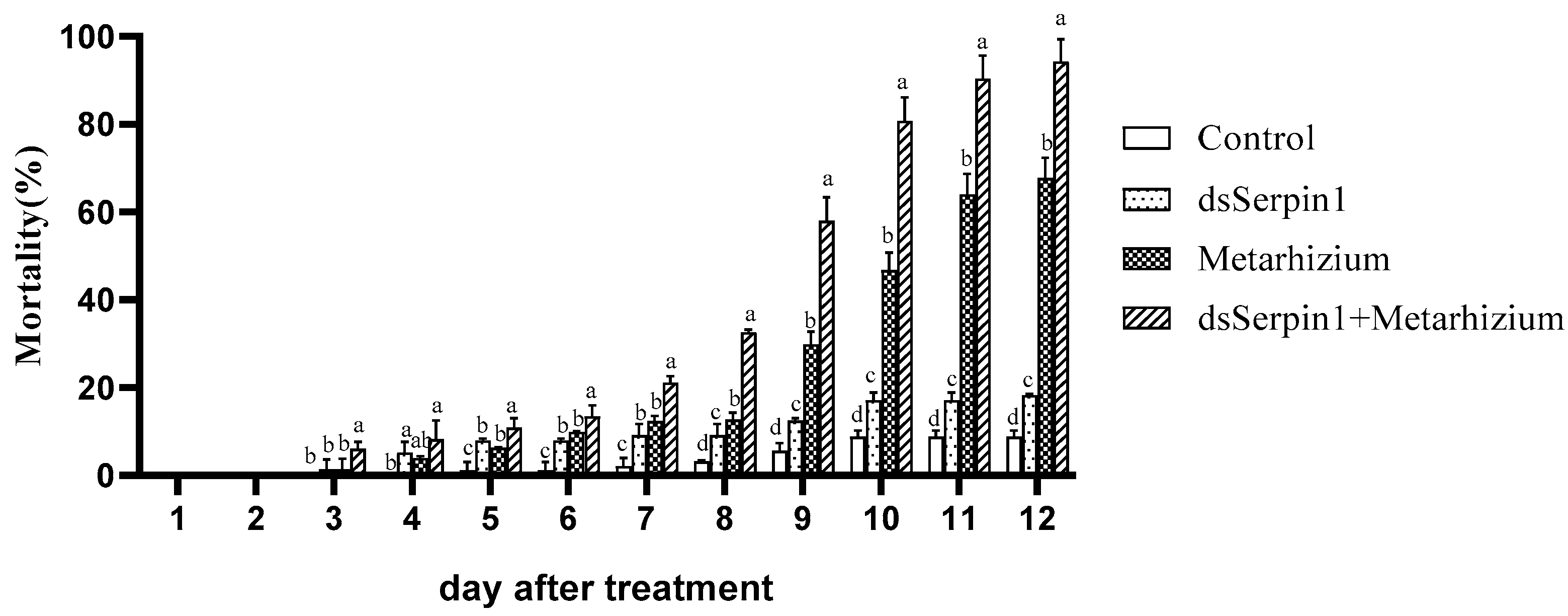{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers | Purpose | Sequence (5′–3′) |
|---|---|---|---|
| serpin1 | serpin1-F | Expression | CGCGGATCC GATGCCAGTCCGCGCCTTCTC |
| serpin1-R | Expression | CCC AAGCTT TTGCGGAGGC CTTTGTGG | |
| qPCR-serpin1-F | Real-Time PCR | TACGCAGGCAAAGGAAAG | |
| qPCR-serpin1-R | Real-Time PCR | ATGGGTTTACGGTGCTC | |
| dsSerpin1-F | RNAi | TAATACGACTCACTATAGGATCAGCACAGCCAGGAAAC | |
| dsSerpin1-R | RNAi | TAATACGACTCACTATAGGCGGCATCGGAGAAGTATTG | |
| Actin | actin-F | Real-Time PCR | GTTACAAACTGGGACGACAT |
| actin-R | Real-Time PCR | AGAAAGCACAGCCTGAATAG | |
| PPAE | PPAE-F | Real-Time PCR | CACCAGCACAAATGAATGAC |
| PPAE-R | Real-Time PCR | CAACGACAATGAGGCACAG | |
| PPO | PPO-F | Real-Time PCR | AAAGACCGCAGAGGAGAA |
| PPO-R | Real-Time PCR | CCAACGATAGAACACAGGA | |
| defensin | defensin-F | Real-Time PCR | CCAGAAAGCGATGATGCCACTA |
| defensin-R | Real-Time PCR | CACCACAAATCAACGCCAAAGT |
| No. | Treatments | The Concentration of M. anisonliae (Spore/g Bran) | Dosage of ds Serpin1 RNA (mL) |
|---|---|---|---|
| (1) | Metarhizium | 2.50 × 108 | 0 |
| (2) | dsSerpin1 | 0 | 5 |
| (3) | Metarhizium + dsSerpin1 | 2.50 × 108 | 5 |
| (4) | Control | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, B.; Li, H.; Tian, Y.; Abro, N.A.; Nong, X.; Zhang, Z.; Wang, G. Molecular Identification and Immunity Functional Characterization of Lmserpin1 in Locusta migratoria manilensis. Insects 2021, 12, 178. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020178
Li B, Li H, Tian Y, Abro NA, Nong X, Zhang Z, Wang G. Molecular Identification and Immunity Functional Characterization of Lmserpin1 in Locusta migratoria manilensis. Insects. 2021; 12(2):178. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020178
Chicago/Turabian StyleLi, Beibei, Hongmei Li, Ye Tian, Nazir Ahmed Abro, Xiangqun Nong, Zehua Zhang, and Guangjun Wang. 2021. "Molecular Identification and Immunity Functional Characterization of Lmserpin1 in Locusta migratoria manilensis" Insects 12, no. 2: 178. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020178