
Unraveling the Morphological Variation of Triatoma infestans in the Peridomestic Habitats of Chuquisaca Bolivia: A Geometric Morphometric Approach

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
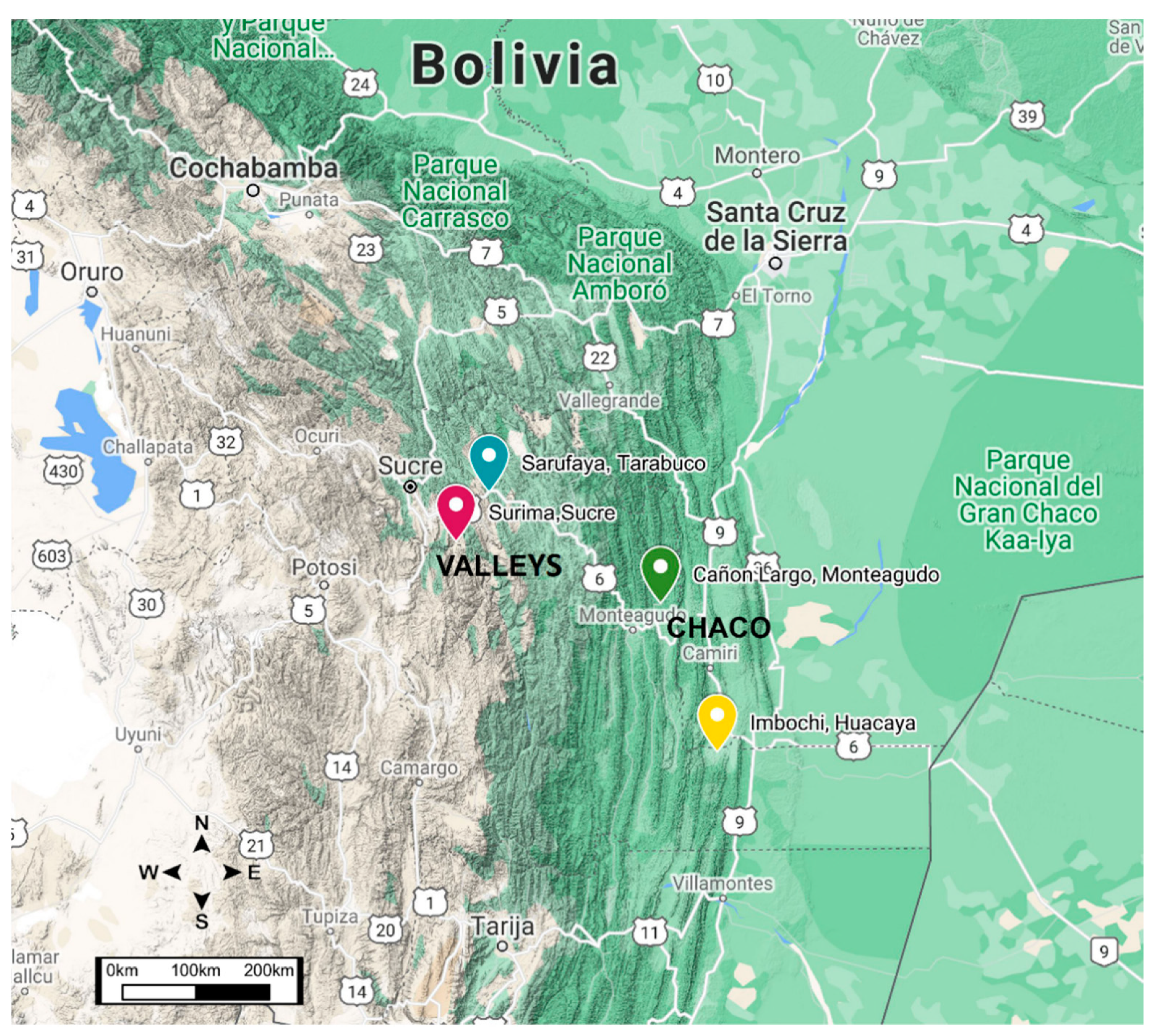
2.1. Study Area
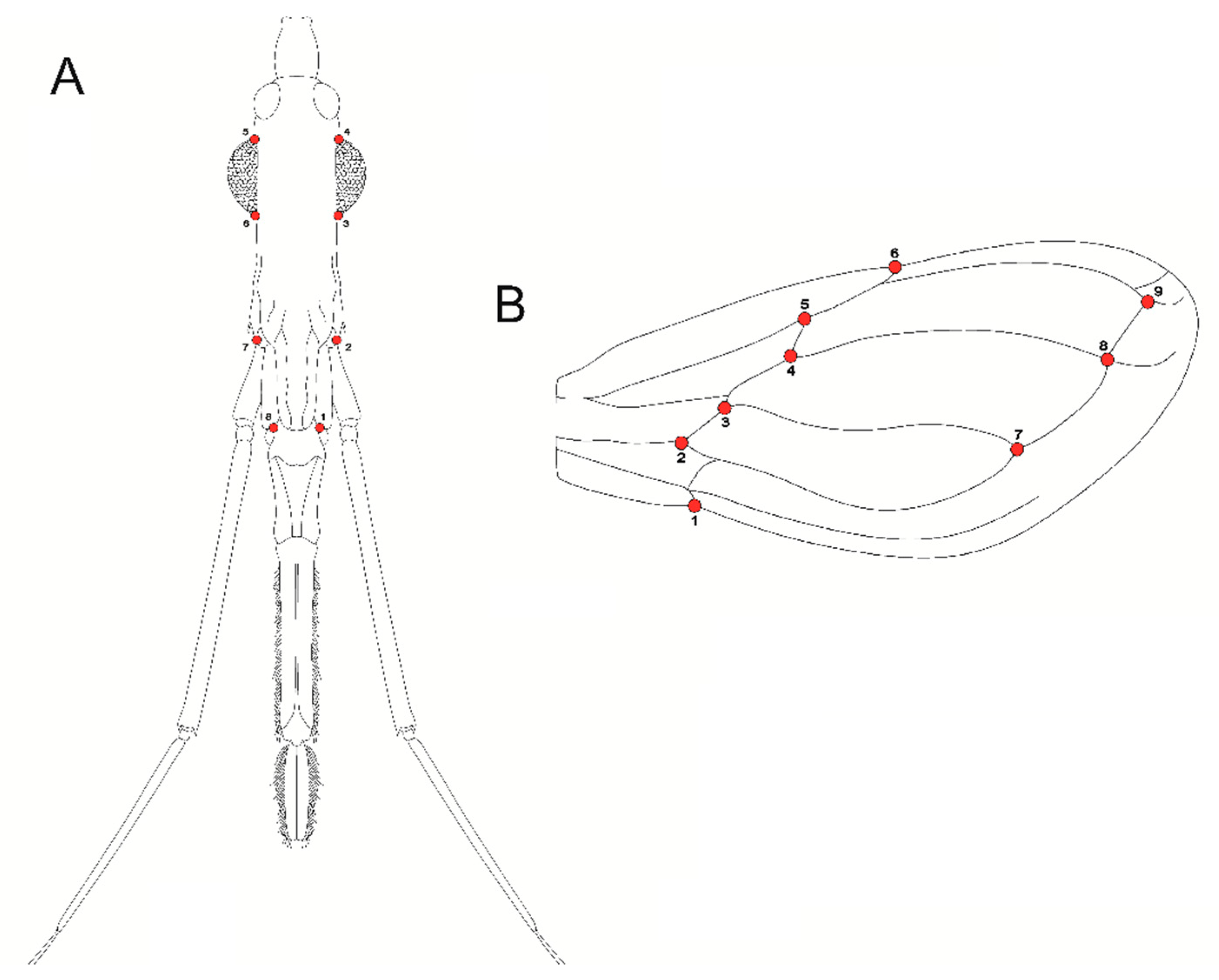
2.2. Insect Sampling and Preparation
2.3. Morphometric Analysis
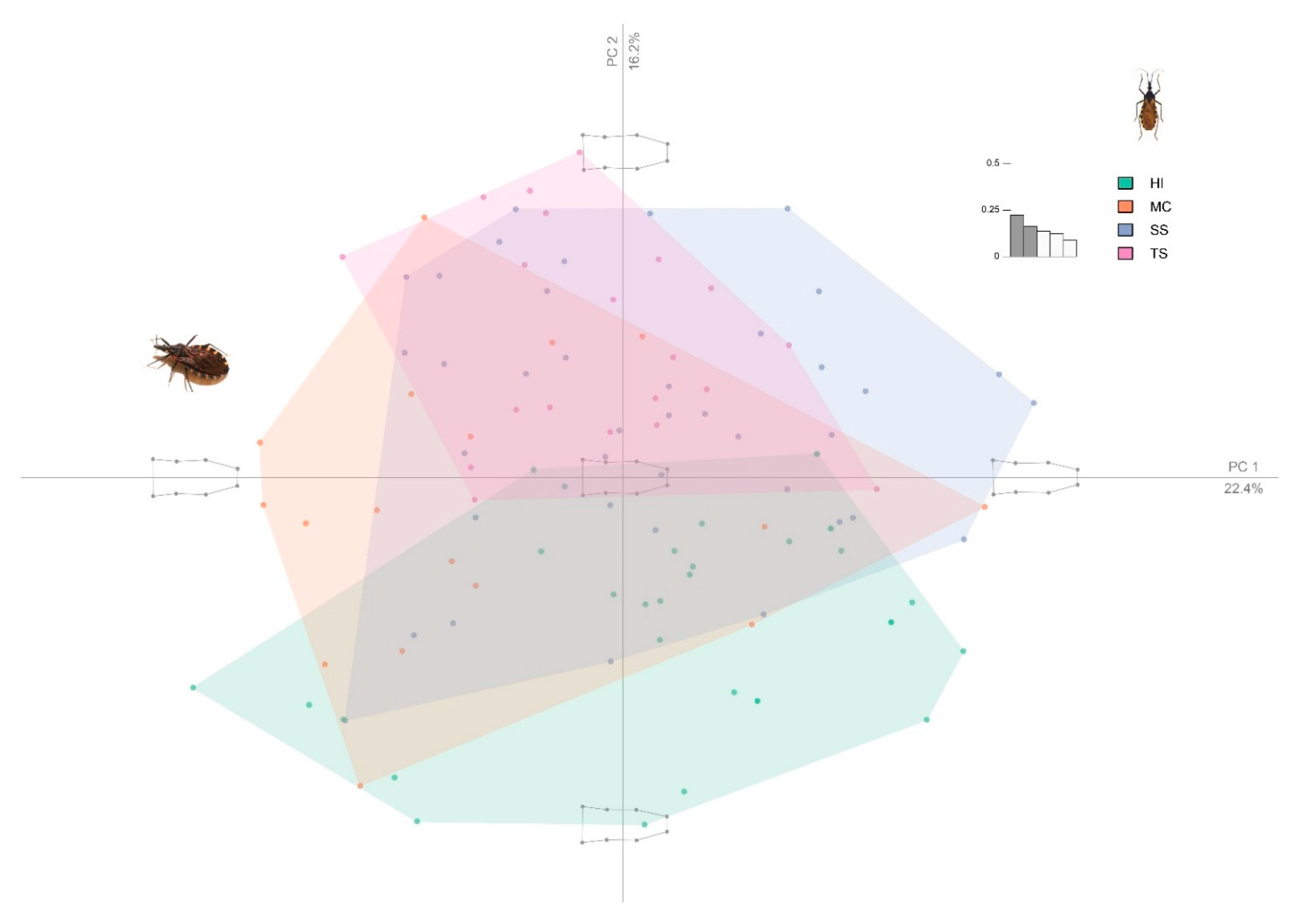
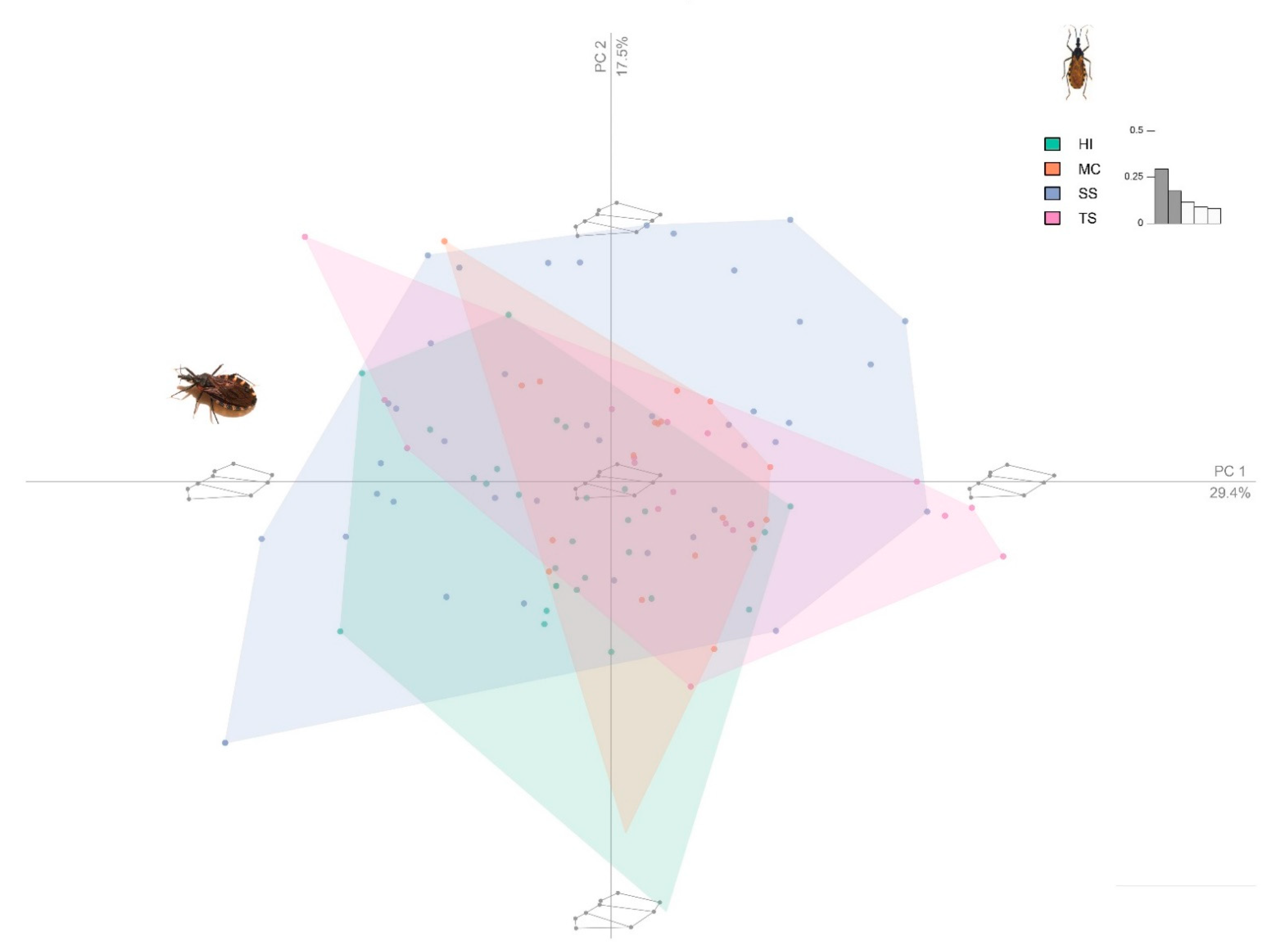
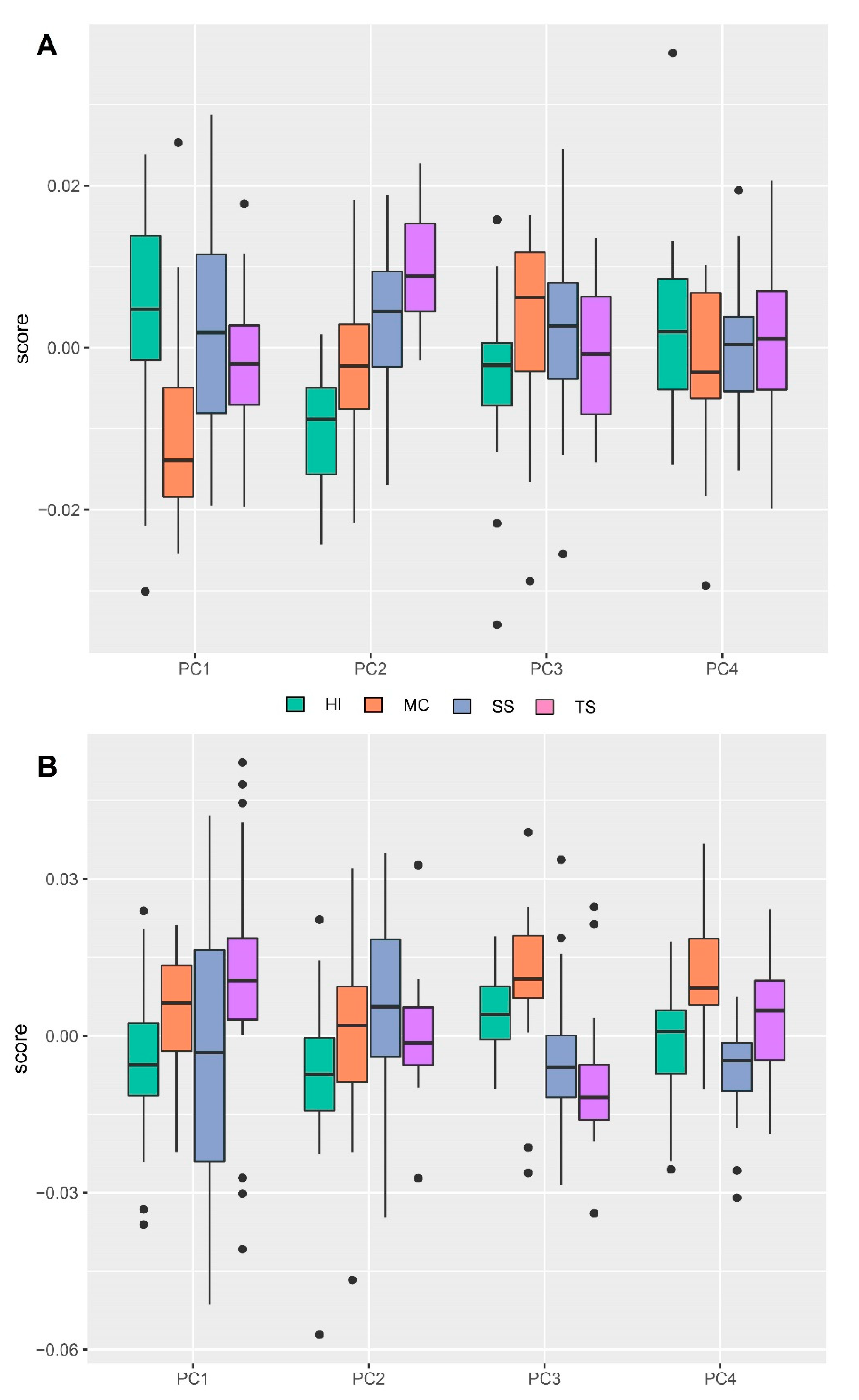
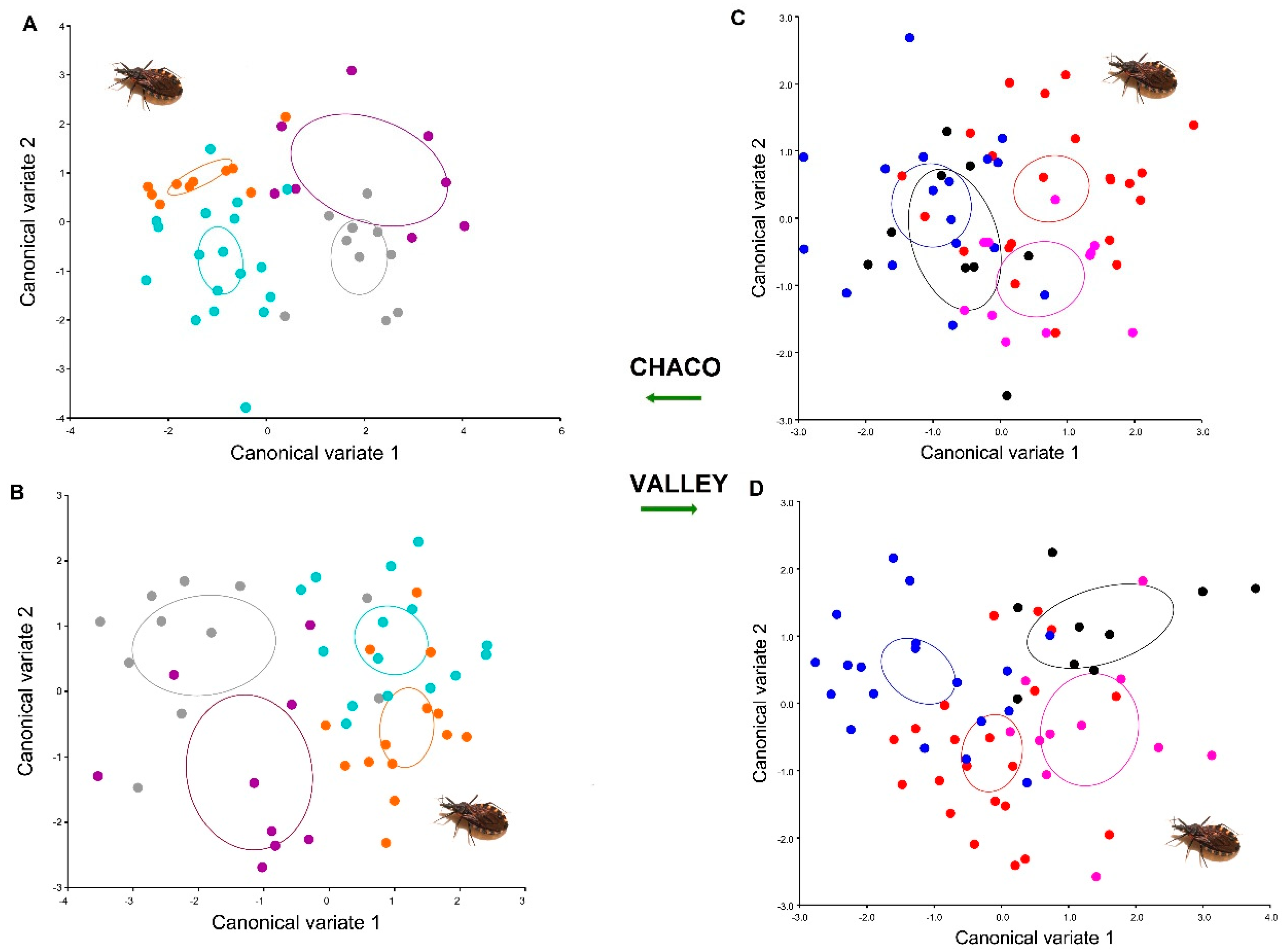
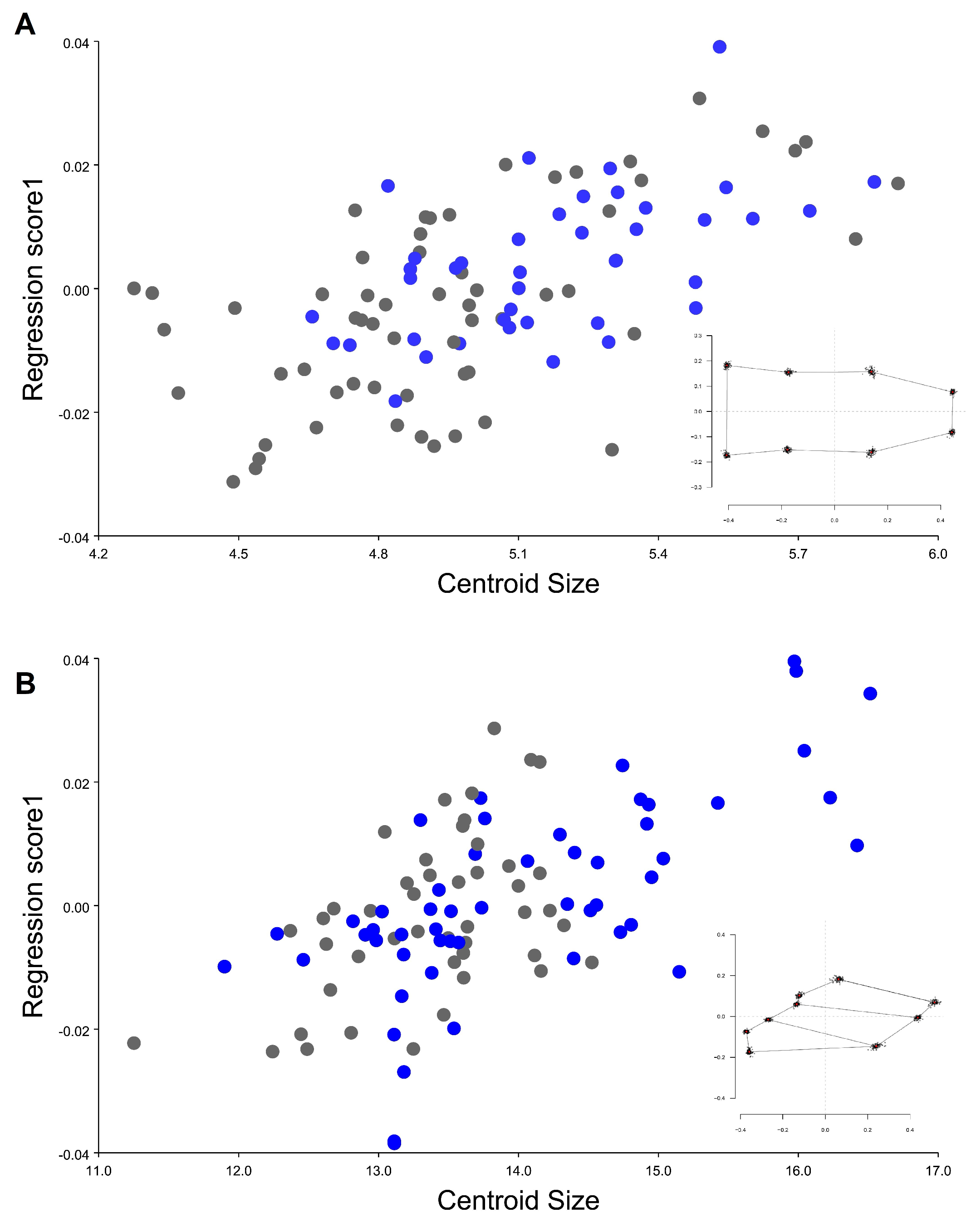
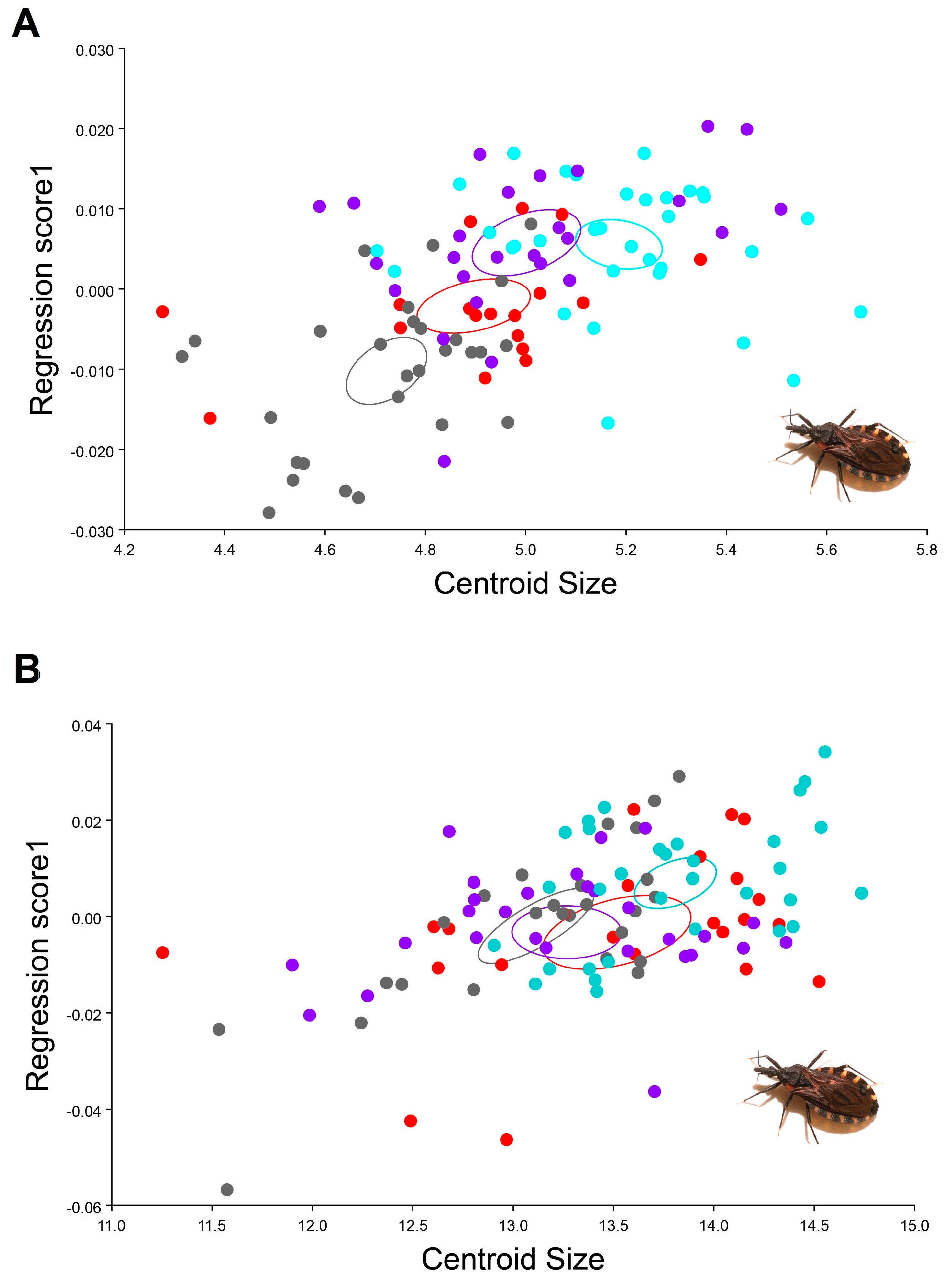
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schofield, C.J. Population dynamics and control of Triatoma infestans. Ann. de la Soc. Belg. de Med. Trop. 1985, 65, 149–164. [Google Scholar]
- Noireau, F.; Flores, R.; Gutierrez, T.; Dujardin, J.P. Detection of Sylvatic Dark Morphs of Triatoma infestans in the Bolivian Chaco. Mem. Inst. Oswaldo Cruz 1997, 92, 583–584. [Google Scholar] [CrossRef] [Green Version]
- Bermudez, H.; Balderrama, F.; Torrico, F. Identification and characterization of sylvatic foci of Triatoma infestans in Central Bolivia. Am. J. Trop. Med. Hyg. 1993, 49. [Google Scholar]
- Giordano, R.; Cortez, J.C.P.; Paulk, S.; Stevens, L. Genetic diversity of Triatoma infestans (Hemiptera: Reduviidae) in Chuquisaca, Bolivia based in the mitocondrial citocrome b gene. Mem. Inst. Oswaldo Cruz 2005, 100, 753–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Programa Chagas Chuquisaca. Informe anual de índices de infestaciónpor Triatoma infestans en el intra y peri domicilio en Chuquisaca; Ministerio de Salud y Deportes de Bolivia: Sucre, Bolivia, 2017. [Google Scholar]
- Noireau, F.; Bastrenta, B.; Catalá, S.; Dujardin, J.P.; Panzera, F.; Torres, M.; Jurberg, J. Silvatic population of Triatoma infestans from the Bolivian Chaco: From field collection to characterization. Mem. Inst. Oswaldo Cruz 2000, 95, 119–122. [Google Scholar] [CrossRef] [Green Version]
- Dujardin, J.P.; Bermudez, H.; Schofield, C. The use of morphometrics in entomological survellance of sylvatic foci of Triatoma infestans in Bolivia. Actra Trop. 1997, 66, 145–153. [Google Scholar] [CrossRef]
- Catalá, S.; Dujardin, J.P. Antennal sensilia patterns indicate geiographic an ecotopic variability among Triatoma infestans (Hemiptera Reduviidae) populations. J. Med. Entomol. 2001, 38, 423–428. [Google Scholar] [CrossRef]
- Dujardin, J.P.; Bermudez, H.; Casini, C.; Schofield, C.J.; Tibayrenc, M. Metric differences between silvatic and domestic Tria-toma infestans (Hemiptera: Reduviidae) in Bolivia. J. Med. Entomol. 1997, 34, 544–551. [Google Scholar] [CrossRef]
- Gaspe, M.S.; Schachter-Broide, J.; Gurevitz, J.M.; Kitron, U.; Gürtler, R.; Dujardin, J.P. Microgeographic spatial structuring of Triatoma infestans (Hemiptera Reduviidae) poulation using wing geometric morphometry in the Argentine Chaco. J. Med. Entomol. 2012, 49, 504–514. [Google Scholar] [CrossRef]
- Benitez, H.A. Sexual dimorphism using geometric morphometric approach. In Sexual Dimorphism; Moryama, H., Ed.; IntechOpen: London, UK, 2013; pp. 35–50. [Google Scholar] [CrossRef]
- Vilaseca, C.; Mendez, M.A.; Pinto, C.F.; Benítez, H.A. Assessment of Shape Variation Oatterns in Triatoma infestans Klug (1834) (Hemiptera: Rediviidae: Triatominae): A first Report in Populations from Bolivia. Insects 2020, 11, 274. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M. Sexual Selection; Princeton University Press: Princeton, UK, 1994. [Google Scholar]
- Fairbairn, D.J. Allometry for sexual size dimorphism: Pattern and process in the coevolution of body size in males and females. Annu. Rev. Ecol. Syst. 1997, 28, 659–687. [Google Scholar] [CrossRef]
- Teder, T.; Tammaru, T. Sexual size dimorphism within species increases with body size in insects. Oikos 2005, 108, 321–334. [Google Scholar] [CrossRef]
- Benítez, H.A.; Vargas, H.A. Sexual dimorphism and population differentiation in the Chilean Neotropical moth Macaria mirthae (Lepidoptera, Geometridae): A wing geometric morphometric example. Rev. Bras. de Èntomol. 2017, 61, 365–369. [Google Scholar] [CrossRef]
- Pigliucci, M.; Murren, C.J.; Schlichting, C.D. Phenotypic plasticity and evolution by genetic assimilation. J. Exp. Biol. 2006, 209, 2362–2367. [Google Scholar] [CrossRef] [Green Version]
- West-Eberhard, M.J. Phenotypic plasticity and the origins of diversity. Annu. Rev. Ecol. Syst. 1989, 20, 249–278. [Google Scholar] [CrossRef]
- West-Eberhard, M.J. Developmental plasticity and the origin of species differences. Proc. Natl. Acad. Sci. USA 2005, 102, 6543–6549. [Google Scholar] [CrossRef] [Green Version]
- Dujardin, J.P.; Panzera, P.; Schofield, C.J. Triatominae as a model of morphological plasticity under ecological pressure. Mem. Inst. Oswaldo Cruz 1999, 94, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Hernández, M.L.; Abrahan, L.; Dujardin, J.P.; Gorla, D.E.; Catalá, S. Phenotypic variability and population structure of per-domestic Triatoma infestans in rural areas of the aird Chaco (Western Argentina: Spatial influence of macro and microhabitats. Vector Borne Zoonotic Dis. 2011, 11, 503–513. [Google Scholar] [CrossRef]
- Dujardin, J.P.; Slice, D. Encyclopedia of Infectious Diseases; Wiley: New York, NY, USA, 2007. [Google Scholar]
- Hernández, M.L.; Amelotti, I.; Catalá, S.; Gorla, D.E. Does nutrition influence sexual dimorphism in Triatoma infestans (He-miptera: Reduviidae) of natural habitats? Rev. Soc. Entomol. Argent. 2018, 77, 1–10. [Google Scholar] [CrossRef]
- Dujardin, J.; Steindel, M.; Chavez, T.; Machane, M.; Schofield, C. Changes in the Sexual Dimorphism of Triatominae in the Transition from Natural to Artificial Habitats. Mem. Inst. Oswaldo Cruz 1999, 94, 565–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, G.; Maldonado, M. Geografía Ecológica de Bolivia; Fundación I Patiño: Cochabamba, Bolivia, 2002. [Google Scholar]
- Rolf, F.J. TPSdig v.2.27; State University Brook: Stony Brook, NY, USA, 2013. [Google Scholar]
- Rohlf, F.J.; Slice, D. Extensions of the Procustes methods for the optimal superimposition of landmarks. Syst. Zool. 1990, 39, 40–59. [Google Scholar] [CrossRef] [Green Version]
- Fruciano, C. Measurement error in geometric morphometrics. Dev. Genes Evol. 2016, 226, 139–158. [Google Scholar] [CrossRef]
- Klingenberg, J.C. MorphoJ: An integrated sofware package for geometric morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar] [CrossRef]
- Campbell, N.A.; Atchley, W.R. The Geometry of Canonical Variate Analysis. Syst. Zool. 1981, 30, 268–280. [Google Scholar] [CrossRef]
- Jolliffe, I.T. Principal Componet Analysis, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Dos Reis, S.F.; Pessoa, L.M.; Strauss, R.E. Apllication of size-free canonical discriminant analysis to studies of geographic differentiation. Braz. J. Gen. 1990, 13, 509–520. [Google Scholar]
- Bonhomme, V.; Picq, S.; Gaucherel, C.; Claude, J. Momocs: Outline Analysis Using R. J. Stat. Softw. 2014, 56, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Martinez, A.; Olmedo, R.A.; Carvallo, R.U. Una nueva subespecie Argentina de Triatoma infestans. Chagas 1987, 4, 479–480. [Google Scholar]
- Noireau, F.; Flores, R.; Gutierrez, T.; Abad-Franch, F.; Flores, E.; Vargas, F. Natural ecotopes of Triatoma infestans dark morph and other sylvatic Triatomines in the Bolivian Chaco. Trans. R. Soc. Trop. Med. Hyg. 2000, 94, 23–27. [Google Scholar] [CrossRef]
- Noireau, F.; Cortez, M.G.; Monteiro, F.A.; Jansen, A.M.; Torrico, F. Can wild Triatoma infestans foci in Bolivia jeopardize Chagas disease control effors? Trends Parasitol. 2005, 21, 7–10. [Google Scholar] [CrossRef]
- Bonduriansky, R. Convergent evolution of sexual shape dimorphism in Diptera. J. Morphol. 2006, 267, 602–611. [Google Scholar] [CrossRef] [PubMed]
- Gidaszewski, N.A.; Baylac, M.; Klingenberg, C.P. Evolution of sexual dimorphism of wing shape in the Drosophila melanogaster subgroup. BMC Evol. Biol. 2009, 9, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsteller, S.; Adams, D.C.; Collyer, M.L.; Condon, M. Six cryptic species on a single species of host plant: Morphometric evidence for possible reproductive character displacement. Ecol. Entomol. 2009, 34, 66–73. [Google Scholar] [CrossRef]
- Benítez, H.; Parra, L.E.; Sepulveda, E.; Sanzana, M.J. Geometric perspectives of sexual dimorphism in the wing shape of Lepidoptera: The case of Synneuria sp. (Lepidoptera: Geometridae). J. Entomol. Res. Soc. 2011, 13, 53–60. [Google Scholar]
- Benítez, H.A.; Bravi, R.; Parra, L.E.; Sanzana, M.J.; Sepúlveda-Zúniga, E. Allometric and non-allometric pattern in sexual dimorphism discrimination of wing shape in the parasitoid, Ophion intricatum: Do two male morphotypes coexist? J. Insect Sci. 2013, 13, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemic, D.; Benítez, H.; Bazok, R. Intercontinental effect on sexual shape dimorphism and allometric relationships in the beetle pest Diabrotica virgifera virgifera LeConte (Coleoptera: Chrysomelidae). Zool. Anz. Comp. Zool. 2014, 253, 203–206. [Google Scholar] [CrossRef]
- Lemic, D.; Benitez, H.A.; Bjeliš, M.; Ordenes-Claveria, R.; Ninčević, P.; Mikac, K.M.; Pajač Živković, I. Agroecological effect and sexual shape dimorphism in medfly Ceratitis capitata (Diptera: Tephritidae) an example in Croatian populations. Zool. Anz. Comp. Zool. 2020, 288, 118–124. [Google Scholar] [CrossRef]
- Fairbairn, D.J. Evolutionary Studies of Sexual Size Dimorphism; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Cox, R.M.; Skelly, S.L.; John-Alder, H.B. A comparative test of adaptative hypothesis for sexual size dimorphism in lizards. Evolution 2003, 57, 1653–1669. [Google Scholar] [CrossRef]
- Rebinovich, J.E.; Kitron, U.D.; Obed, Y.; Yoshioka, M.; Gottdenker, N.; Chaves, L.F. Ecological patterns of blood-feeding by kissing-bugs (Hemiptera: Redubiidae: Triatominae). Mem. Inst. Oswaldo Cruz 2011, 106, 479–494. [Google Scholar] [CrossRef] [Green Version]
- Gúrtler, R.E.; Fernández, M.P.; Cecere, M.C.; Cohen, J.E. Body size and host of Triatoma infestans population affect the size and blodmeal contens and female fecundity in rural nortwestern Argentina. PLoS Neg. Trop. Dis. 2017, 11, e0006097. [Google Scholar] [CrossRef]
- Hernández, M.L.; Dujardin, J.P.; Gorla, D.E.; Catalá, S.S. Potential sources of Triatoma infestans reinfesting peridomiciles iden-tified by morphological characterization in Los Llanos, La Rioja, Argentina. Mem. Inst. Oswaldo Cruz 2013, 108, 91–97. [Google Scholar] [CrossRef] [Green Version]
- Mikac, K.M.; Douglas, J.; Spencer, J. Wing Shape and Size of the Western Corn Rootworm (Coleoptera: Chrysomelidae) is Related to Sex and Resistance to Soybean-Maize Crop Rotation. J. Econ. Entomol. 2013, 106, 1517–1524. [Google Scholar] [CrossRef] [PubMed]
- Dingle, H. Geographic Variation and Behavioral Flexibility in Milkweed Bug Life Histories. Insect Life History Patterns; Springer: New York, NY, USA, 1981; pp. 57–73. [Google Scholar]
- Dudley, R.; Srygley, R. Flight physiology of neotropical butterflies: Allometry of airspeeds during natural free flight. J. Exp. Biol. 1994, 191, 125–139. [Google Scholar]
- Altizer, S.; Davis, A.K. Populations of monarch butterflies with different migratory behaviors show divergence in wing morphology. Evolution 2010, 64, 1018–1028. [Google Scholar] [CrossRef]
- Lazzari, C.R.; Pereira, M.H.; Lorenzo, M.G. Biología del comportamiento de los vectores de la enfermedad de Chagas. Mem. Inst. Oswaldo Cruz 2013, 108, 34–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinchera-Donoso, D.; Tregenza, T. Fecundity Selection and the Evolution of Reproductive Ouput and Sex-Specific Body Size in the Liolaemus Lizard Adaptive Radiation. Evol. Biol. 2011, 38, 197–207. [Google Scholar] [CrossRef]
- Darwin, C. The Descent of Man and Selection in Relation to Sex; Humboldt: New York, NY, USA, 1894. [Google Scholar]
- Lehane, M.J.; McEwen, P.K.; Whitaker, C.J.; Schofield, C.J. The role of temperature and nutritional status in flight initiation by Triatona infestans. Acta Trop. 1992, 52, 27–38. [Google Scholar] [CrossRef]
- Papadopoulos, N.T.; Carey, J.R.; Katsoyannos, B.I.; Kouloussis, N.A. Overwintering of the Mediterranean Fruit Fly (Diptera: Tephritidae) in Northern Greece. Ann. Entomol. Soc. Am. 1996, 89, 526–534. [Google Scholar] [CrossRef]
- Papadopoulos, N.T.; Katsoyannos, B.I.; Carey, J.R.; Kouloussis, N.A. Seasonal and Annual Occurrence of the Mediterranean Fruit Fly (Diptera: Tephritidae) in Northern Greece. Ann. Entomol. Soc. Am. 2001, 94, 41–50. [Google Scholar] [CrossRef]
- Papadopoulos, N.T.; Katsoyannos, B.I.; Nestle, D. Spatial Autocorrelation Analysis of a Ceratitis capitata (Diptera: Tephritidae) Adult Population in a Mixed Deciduous Fruit Orchard in Northern Greece. Environ. Entomol. 2003, 32, 319–326. [Google Scholar] [CrossRef] [Green Version]
- Katsoyannos, B.I.; Kouloussis, N.A.; Carey, J.R. Seasonal and annual occurrence of Mediterranean fruit flies (Diptera: Tephri-tidae) on Chios island, Greece: Differences between two neighboring citrus orchards. Ann. Entomol. Soc. Am. 1998, 91, 43–51. [Google Scholar] [CrossRef]
- Navarro-Campos, C.; Martínez-Ferrer, M.T.; Campos, J.M.; Fibla, J.M.; Alcaide, J.; Bargues, L.; Marzal, C.; Garcia-Marí, F. The Influence of Host Fruit and Temperature on the Body Size of Adult Ceratitis capitata (Diptera: Tephritidae) Under Laboratory and Field Conditions. Environ. Entomol. 2011, 40, 931–938. [Google Scholar] [CrossRef]
- Gürtler, R.R.; Kitron, U.; Cecere, M.C.; Segura, E.L.; Cohen, J.E. Sustainable vector control and management of Chagas disease in the Gran Chaco, Argentina. Proc. Natl. Acad. Sci. USA 2007, 104, 16194–16199. [Google Scholar] [CrossRef] [Green Version]
- Vazquez-Prokopec, G.M.; Ceballos, L.A.; Marcet, P.L.; Cecere, M.C.; Cardinal, M.V.; Kitron, U.; Gurtler, R.E. Seasonal variations in active dispersal of natural populations of Triatoma infestans in rural north-western Argentina. Med. Vet. Entomol. 2006, 20, 273–279. [Google Scholar] [CrossRef] [Green Version]
- Schachter-Broide, J.; Gürtler, R.E.; Uriel, K.; Dujardin, J.P. Temporal Variations of Wing Size and Shape of Triatona infestans (Hemptera:Reduviidae) Populations from Northwestern Argentina Using Geometric Morphometry. J. Med. Entomol. 2009, 46, 994–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colinet, H.; Sinclair, B.J.; Vernon, P.; Renault, D. Insects in Fluctuating Thermal Environments. Annu. Rev. Entomol. 2015, 60, 123–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moraes, E.M.; Manfrin, M.H.; Laus, A.C.; Rosada, R.S.; Bomfin, S.C.; Sene, F.M. Wing shape heritability and morphological divergence of the sibling species Drosophila mercatorum and Drosophila paranaensis. Heredity 2004, 92, 466–473. [Google Scholar] [CrossRef]
- Dujardin, J.-P. Morphometrics applied to medical entomology. Infect. Genet. Evol. 2008, 8, 875–890. [Google Scholar] [CrossRef] [PubMed]
- Natero, J.; Malerba, R.; Rodriguez, C.S.; Crocco, I. Phenotypic plasticity in response to food sourse in Triatoma infestans (Kug 1834) (Hemiptera Rdiviidae; Triatominae). Infect. Genet. Evol. 2013, 19, 38–44. [Google Scholar] [CrossRef]
- Lunardi, R.R.; Benítez, H.A.; Cámara, T.P.; Gomez, L.P.; Waagner, A.S. Head shape variation in response to diet in Triatoma williami (Hemiptera, Rduviidae; Triatominae), a possible Chagas disease vector of legal Amazonia. Zool. Anz. 2017, 267, 87–193. [Google Scholar] [CrossRef]
- Marcet, P.; Mora, M.; Cutrera, A.; Jones, L.; Gürtler, R.; Kitron, U.; Dotson, E.M. Genetic structure of Triatoma infestans populations in rural communities of Santiago del Estero, northern Argentina. Infect. Genet. Evol. 2008, 8, 835–846. [Google Scholar] [CrossRef] [Green Version]
- Schlichting, C.D. The role of phenotypic plasticity in diversification. In Phenotypic Plasticity: Functional and Conceptual Approaches; deWitt, T.J., Scheiner, S.M., Eds.; Oxford University Press: Oxford, UK, 2004; pp. 191–200. [Google Scholar]
- Murren, C.J.; Denning, W.; Pigliucci, M. Relationships between Vegetative and Life History Traits and Fitness in a Novel Field Environment: Impacts of Herbivores. Evol. Ecol. 2005, 19, 583–601. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| View | Region | Mahalanobis Distance | Procrustes Distance | ||||
|---|---|---|---|---|---|---|---|
| F/Chaco | F/Valley | M/Chaco | F/Chaco | F/Valley | M/Chaco | ||
| Head dorsal | F/Valley | 1.81138 * | 0.0123 | ||||
| M/Chaco | 1.8254 * | 2.2565 * | 0.0132 | 0.0168 * | |||
| M/Valley | 1.7989 * | 1.3489 | 2.2707 * | 0.0131 | 0.0073 | 0.0177* | |
| Wings | F/Valley | 2.5087 * | 0.0235* | ||||
| M/Chaco | 1.3489 | 2.2660 * | 0.0107 | 0.212 | |||
| M/Valley | 2.3459 * | 1.5397 * | 2.1843 * | 0.0191 | 0.0187 | 0.0193 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vilaseca, C.; Méndez, M.A.; Pinto, C.F.; Lemic, D.; Benítez, H.A. Unraveling the Morphological Variation of Triatoma infestans in the Peridomestic Habitats of Chuquisaca Bolivia: A Geometric Morphometric Approach. Insects 2021, 12, 185. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020185
Vilaseca C, Méndez MA, Pinto CF, Lemic D, Benítez HA. Unraveling the Morphological Variation of Triatoma infestans in the Peridomestic Habitats of Chuquisaca Bolivia: A Geometric Morphometric Approach. Insects. 2021; 12(2):185. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020185
Chicago/Turabian StyleVilaseca, Carolina, Marco A. Méndez, Carlos F. Pinto, Darija Lemic, and Hugo A. Benítez. 2021. "Unraveling the Morphological Variation of Triatoma infestans in the Peridomestic Habitats of Chuquisaca Bolivia: A Geometric Morphometric Approach" Insects 12, no. 2: 185. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020185