
First Multi-Target Application of Exclusion Net in Nectarine Orchards: Effectiveness against Pests and Impact on Beneficial Arthropods, Postharvest Rots and Fruit Quality

 ,
,  , ,
, ,
Abstract
:Simple Summary
Abstract
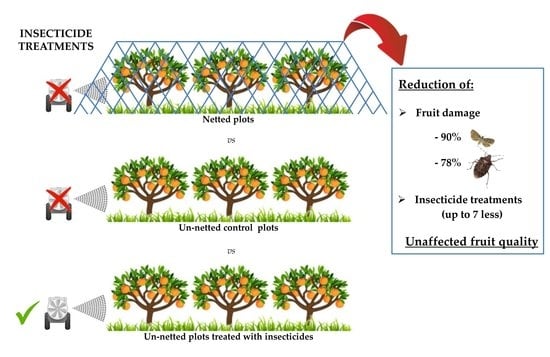
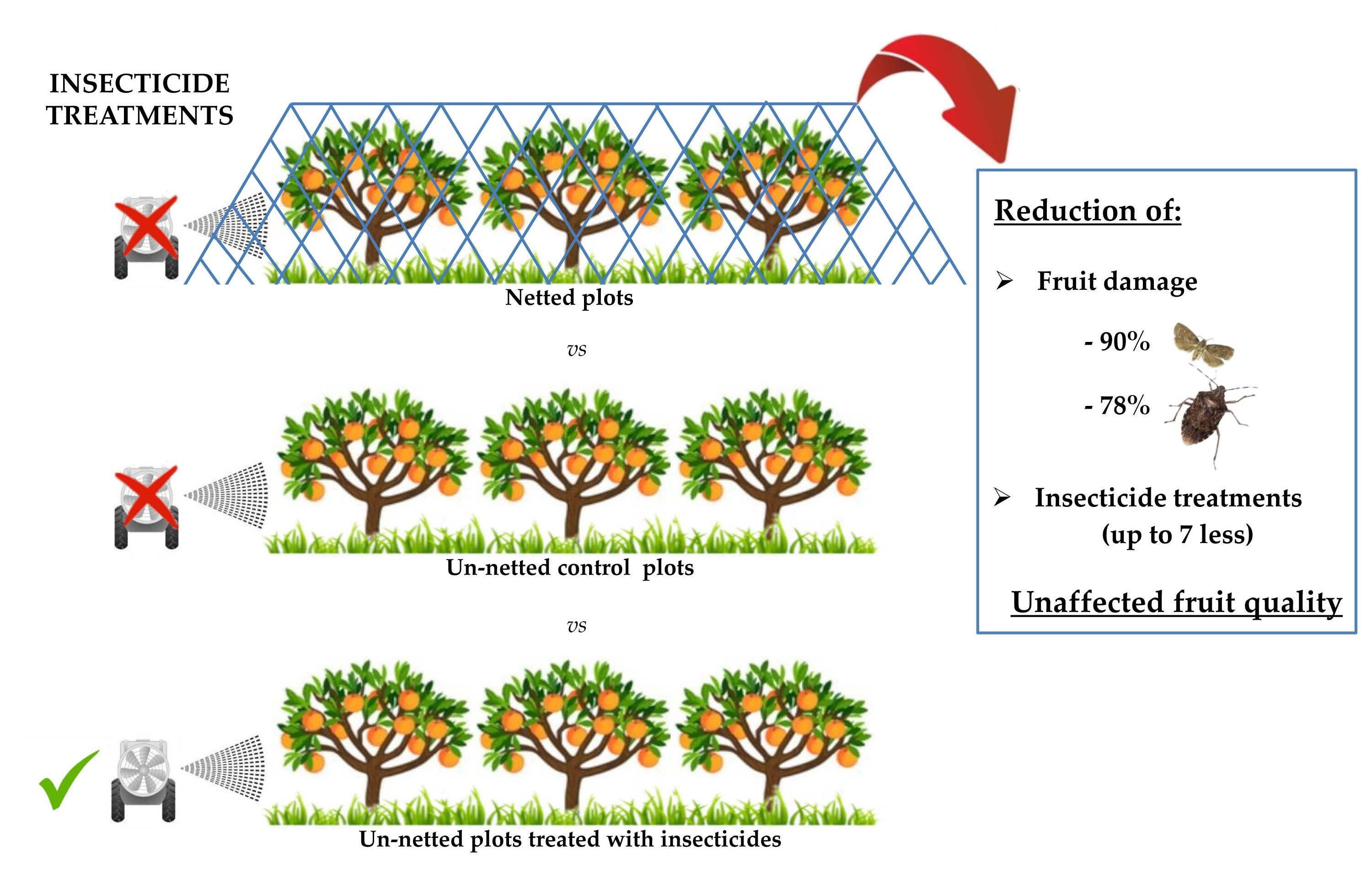
1. Introduction
2. Materials and Methods
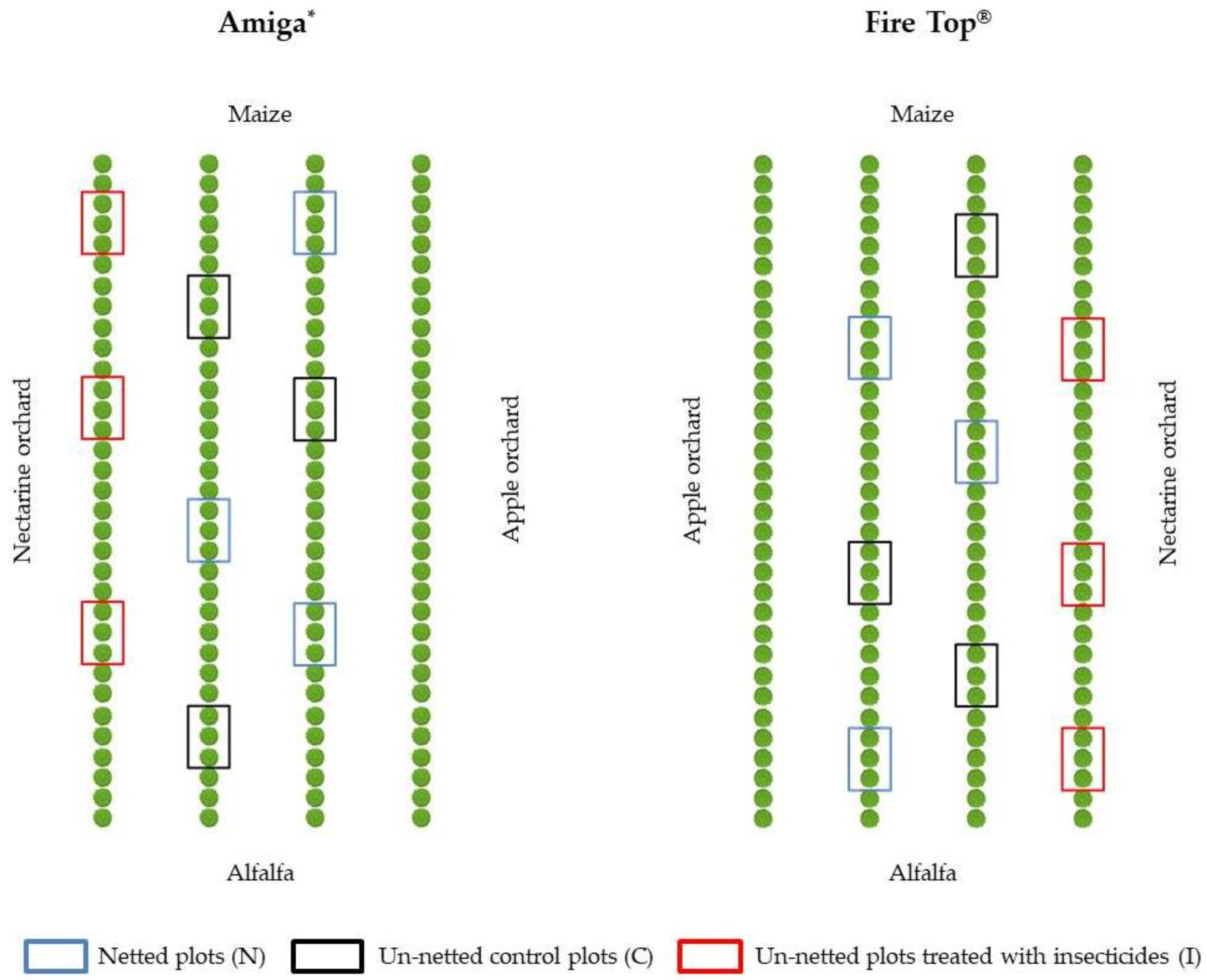
2.1. Experimental Sites
2.2. Pest and Beneficial Insect Monitoring
2.2.1. Fruit Moths
2.2.2. Halyomorpha halys
2.2.3. Drosophila suzukii
2.2.4. Other Arthropods
2.3. Evaluation of Fruit Damage Caused by Pests
2.4. Evaluation of Postharvest Rots
2.5. Fruit Quality and Nutraceutical Analyses
2.6. Statistical Analyses
3. Results
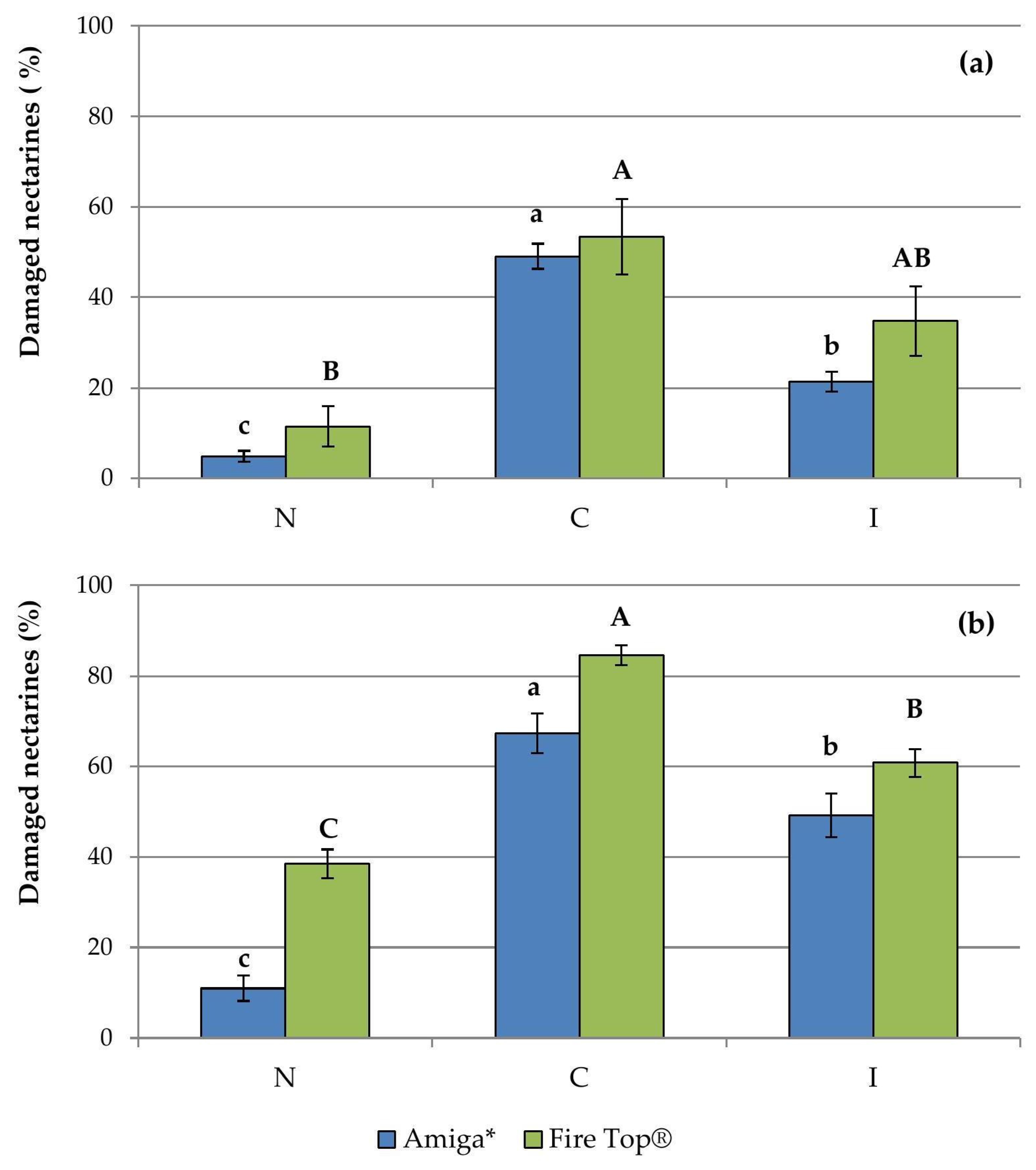
3.1. Monitoring of and Damage Caused by Grapholita molesta and Anarsia lineatella
3.2. Monitoring of and Damage Caused by Halyomorpha halys
3.3. Monitoring of Drosophila suzukii
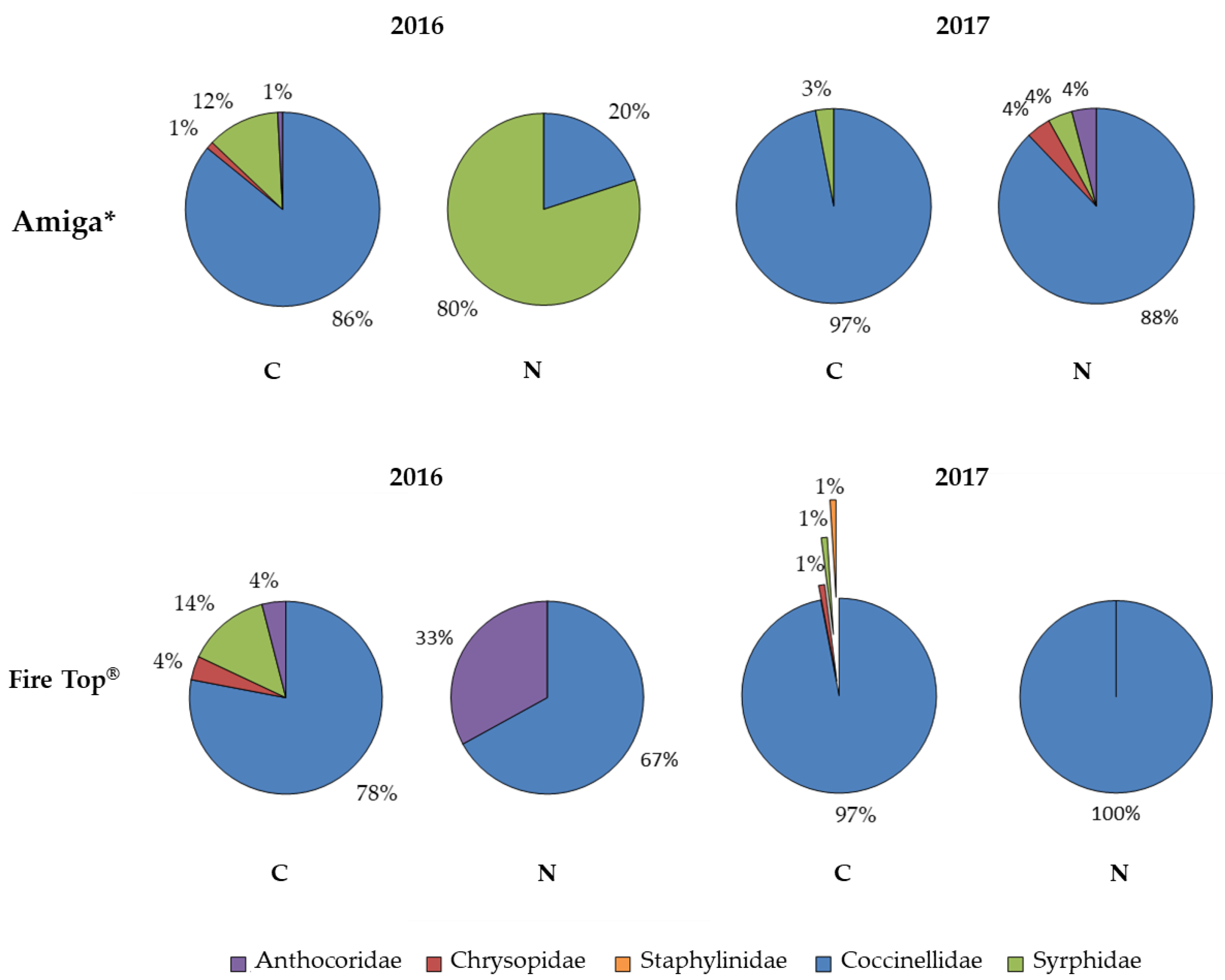
3.4. Monitoring of Beneficial Insects
3.5. Postharvest Rots
3.6. Fruit Quality and Nutraceutical Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ben-Yakir, D.; Antignus, Y.; Offir, Y.; Shahak, Y. Photoselective nets and screens can reduce insect pests and diseases in agricultural crops. Acta Hortic. 2014, 1015, 95–102. [Google Scholar] [CrossRef]
- Pree, D.J.; Whitty, K.J.; Van Driel, L.; Walker, G.M. Resistance to insecticides in Oriental fruit moth populations (Grapholita molesta) from the Niagara peninsula of Ontario. Can. Entomol. 1998, 130, 245–256. [Google Scholar] [CrossRef]
- Arthurs, S.P.; Lacey, L.A.; Miliczky, E.R. Evaluation of the codling moth granulovirus and spinosad for codling moth control and impact on non-target species in pear orchards. Biol. Control 2007, 41, 99–109. [Google Scholar] [CrossRef]
- Witzgall, P.; Kirsch, P.; Cork, A. Sex pheromones and their impact on pest management. J. Chem. Ecol. 2010, 36, 80–100. [Google Scholar] [CrossRef]
- Preti, M.; Knight, A.L.; Angeli, S. Improving Grapholita molesta monitoring in peach and nectarine orchards under mating disruption by using bisexual lures. IOBC/WPRS Bull. 2019, 146, 176–180. [Google Scholar]
- Il’ichev, A.L.; Williams, D.G.; Drago, A. Distribution of the oriental fruit moth Grapholita molesta Busck (Lep., Tortricidae) infestation on newly planted peaches before and during 2 years of mating disruption. J. Appl. Entomol. 2003, 127, 348–353. [Google Scholar] [CrossRef]
- Butault, J.P.; Dedryver, C.A.; Gary, C.; Guichard, L.; Jacquet, F.; Meynard, J.M.; Nicot, P.; Pitrat, M.; Reau, R.; Sauphanor, B.; et al. Quelles Voies Pour Réduire L’usage des Pesticides? Synthèse du Rapport d’Étude; INRA Editeur: Paris, France, 2010. [Google Scholar]
- Briassoulis, D.; Mistriotis, A.; Eleftherakis, D. Mechanical behaviour and properties of agricultural nets—Part I: Testing methods for agricultural nets. Polym. Test 2007, 26, 822–832. [Google Scholar] [CrossRef]
- Castellano, S.; Scarascia Mugnozza, G.; Russo, G.; Briassoulis, D.; Mistriotis, A.; Hemming, S.; Waaijenberg, D. Plastic nets in agriculture: A general review of types and applications. Appl. Eng. Agric. 2008, 24, 799–808. [Google Scholar] [CrossRef]
- Chouinard, G.; Firlej, A.; Cormier, D. Going beyond sprays and killing agents: Exclusion, sterilization and disruption for insect pest control in pome and stone fruit orchards. Sci. Hortic. 2016, 208, 13–27. [Google Scholar] [CrossRef] [Green Version]
- Tasin, M.; Demaria, D.; Ryne, C.; Cesano, A.; Galliano, A.; Anfora, G.; Ioriatti, C.; Alma, A. Effect of anti-hail nets on Cydia pomonella behavior in apple orchards. Entomol. Exp. Appl. 2008, 129, 32–36. [Google Scholar] [CrossRef]
- Pasqualini, E.; Caruso, S.; Piccinini, M.; Vergnani, S.; Salvatorelli, F.; Maini, S.; Ventura, F. La rete anti-insetto e gli effetti sulla carpocapsa in Emilia-Romagna. Rivista di Fruttic. Ortofloric. 2013, 75, 72–78. [Google Scholar]
- Dib, H.; Sauphanor, B.; Capowiez, Y. Effect of codling moth exclusion nets on the rosy apple aphid, Dysaphis plantaginea, and its control by natural enemies. Crop Prot. 2010, 29, 1502–1513. [Google Scholar] [CrossRef]
- Candian, V.; Pansa, M.G.; Briano, R.; Peano, C.; Tedeschi, R.; Tavella, L. Exclusion nets: A promising tool to prevent Halyomorpha halys from damaging nectarines and apples in NW Italy. Bull. Insectology 2018, 71, 21–30. [Google Scholar]
- Candian, V.; Pansa, M.G.; Santoro, K.; Spadaro, D.; Tavella, L.; Tedeschi, R. Photoselective exclusion netting in apple orchards: Effectiveness against pests and impact on beneficial arthropods, fungal diseases and fruit quality. Pest. Manag. Sci. 2020, 76, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Kuhrt, U.; Samietz, J.; Höhn, H.; Dorn, S. Modelling the phenology of codling moth: Influence of habitat and thermoregulation. Agr. Ecosyst. Environ. 2006, 117, 29–38. [Google Scholar] [CrossRef]
- Sauphanor, B.; Severac, G.; Maugin, S.; Toubon, J.F.; Apowiez, Y. Exclusion netting may alter reproduction of the codling moth (Cydia pomonella) and prevent associated fruit damage to apple orchards. Entomol. Exp. Appl. 2012, 145, 134–142. [Google Scholar] [CrossRef]
- Chouinard, G.; Veilleux, J.; Pelletier, F.; Larose, M.; Philion, V.; Cormier, D. Impact of exclusion netting row covers on arthropod presence and crop damage to ‘Honeycrisp’ apple trees in North America: A five-year study. Crop. Prot. 2017, 98, 248–254. [Google Scholar] [CrossRef]
- Tortricids of Agricultural Importance, Interactive Keys Developed in Lucid 3.5. Available online: https://idtools.org/id/leps/tortai/Grapholita_molesta.htm (accessed on 2 April 2020).
- Bellamy, D.E.; Sisterson, M.S.; Walse, S.S. Quantifying host potentials: Indexing postharvest fresh fruits for spotted wing Drosophila, Drosophila suzukii. PLoS ONE 2013, 8, 61227. [Google Scholar] [CrossRef] [Green Version]
- Identify Drosophila suzukii. Oregon Department of Agricolture. Available online: https://www.oregon.gov/oda/shared/documents/publications/ippm/spottedwingdrosophilaidkey.pdf (accessed on 8 June 2015).
- Acebes-Doria, A.L.; Leskey, T.C.; Bergh, J.C. Injury to apples and peaches at harvest from feeding by Halyomorpha halys (Stål) (Hemiptera: Pentatomidae) nymphs early and late in the season. Crop. Prot. 2016, 89, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Franco Ortega, S.; Bustos Lopez, M.P.; Nari, L.; Boonham, N.; Gullino, M.L.; Spadaro, D. Rapid detection of Monilinia fructicola and Monilinia laxa on peaches and nectarines using loop-mediated isothermal amplification. Plant Dis. 2019, 103, 2305–2314. [Google Scholar] [CrossRef]
- Côté, M.-J.; Tardif, M.C.C.; Meldrum, A.J.; Cote, M.J.; Tardif, M.C.C.; Meldrum, A.J. Identification of Monilinia fructigena, M. fructicola, M. laxa and Monilia polystroma on inoculated and naturally infected fruit using multiplex PCR. Plant Dis. 2004, 88, 1219–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahak, Y.; Gussakovsky, E.E.; Cohen, Y.; Lurie, S.; Stern, R.; Kfir, S.; Naor, A.; Atzmon, I.; Doron, I.; Greenblatavron, Y. ColorNets: A new approach for light manipulation in fruit trees. Acta Hortic. 2004, 636, 609–616. [Google Scholar] [CrossRef]
- Rajapakse, N.C.; Shahak, Y. Light quality manipulation by horticulture industry. In Light and Plant Development; Whitelam, G., Halliday, K., Eds.; Blackwell Publishing: Oxford, UK,, 2007; pp. 290–312. [Google Scholar]
- Retamales, J.B.; Monecino, J.M.; Lobos, G.A.; Rojas, L.A. Colored shading increase yields and profitability of highbush blueberries. Acta Hortic. 2008, 770, 193–198. [Google Scholar] [CrossRef]
- Shahak, Y. Photo-selective netting for improved performance of horticultural crops: A review of ornamental and vegetable studies in Israel. Acta Hortic. 2008, 770, 161–168. [Google Scholar] [CrossRef]
- Giaccone, M.; Basile, B.; Forlani, M. Tree vigor, fruit yield and quality of nectarine trees under red photoselective anti-hail nets in southern Italy. Acta Hortic. 2009, 962, 387–393. [Google Scholar] [CrossRef]
- Basile, B.; Giaccone, M.; Cirillo, C.; Ritieni, A.; Graziani, G.; Shahak, Y.; Forlani, M. Photo-selective hail nets affect fruit size and quality in Hayward kiwifruit. Sci. Hortic. 2012, 141, 91–97. [Google Scholar] [CrossRef]
- Shahak, Y. Photoselective netting: An overview of the concept, research and development and practical implementation in agriculture. Acta Hortic. 2014, 1015, 155–162. [Google Scholar] [CrossRef]
- Mashabela, M.N.; Selahle, K.M.; Soundy, P.; Crosby, K.M.; Sivakumar, D. Bioactive compounds and fruit quality of green sweet pepper grown under different colored shade netting during postharvest storage. J. Food Sci. 2015, 80, 2612–2618. [Google Scholar] [CrossRef] [PubMed]
- Morrison, W.R.; Milonas, P.; Kapantaidaki, D.E.; Cesari, M.; Di Bella, E.; Guidetti, R.; Haye, T.; Maistrello, L.; Moraglio, S.T.; Piemontese, L.; et al. Attraction of Halyomorpha halys (Hemiptera: Pentatomidae) haplotypes in North America and Europe to baited traps. Sci. Rep. 2017, 7, 16941. [Google Scholar] [CrossRef]
- Morrison, W.R.; Lee, D.H.; Short, B.D.; Khrimian, A.; Leskey, T.C. Establishing the behavioral basis for an attract-and-kill strategy to manage the invasive Halyomorpha halys in apple orchards. J. Pest. Sci. 2016, 89, 81–96. [Google Scholar] [CrossRef]
- Leskey, T.C.; Short, B.D.; Butler, B.R.; Wright, S.E. Impact of the invasive brown marmorated stink bug, Halyomorpha halys (Stål), in mid-Atlantic tree fruit orchards in the United States: Case studies of commercial management. Psyche 2012, 2012, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Shahak, Y.; Ratner, K.; Zur, N.; Offir, Y.; Matan, E.; Yehezkel, H.; Messika, Y.; Posalski, I.; Ben-Yakir, D. Photoselective netting: An emerging approach in protected agriculture. Acta Hortic. 2009, 807, 79–84. [Google Scholar] [CrossRef]
- Ben-Yakir, D.; Hadar, M.D.; Offir, Y.; Chen, M.; Tregerman, M. Protecting crops from pests using OptiNet® and ChromatiNet® shading nets. Acta Hortic. 2008, 770, 205–212. [Google Scholar] [CrossRef]
- Romet, L.; Severac, G.; Warlop, F. Overview of ALT’CARPO concept and its development in France. In Proceedings of the Ecofruit-14th International Conference on Cultivation Technique and Phytopathological Problems in Organic Fruit-Growing, Hohenheim, Germany, 22–24 February 2010; Fördergemeinschaft Ökologischer Obstbau eV: Weinsberg, Germany, 2010; pp. 176–182. [Google Scholar]
- Doukas, D.; Payne, C.C. Greenhouse whitefly (Homoptera: Aleyrodidae) dispersal under different UV-light environments. J. Econ. Entomol. 2007, 100, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Legarrea, S.; Weintraub, P.G.; Plaza, M.; Viñuela, E.; Fereres, A. Dispersal of aphids, whiteflies and their natural enemies under photoselective nets. Biocontrol 2012, 57, 523–532. [Google Scholar] [CrossRef] [Green Version]
- Raviv, M.; Antignus, Y. UV radiation effects on pathogens and insect pests of greenhouse-grown crops. Photochem. Photobiol. 2004, 79, 219–226. [Google Scholar] [CrossRef]
- Caruso, S.; Cera, M.C. Control strategies for the cherry fruit fly (Rhagoletis cerasi) in organic farming. OILB SROP Bull. 2004, 27, 99–104. [Google Scholar]
- Charlot, G.; Weydert, C.; Millan, M.; Brachet, M.L.; Warlop, F. Nets and covers to protect cherry trees from rain and insects. In Proceedings of the 16th International Conference on Organic-Fruit Growing, Hohenheim, Germany, 17–19 February 2014; pp. 222–227. [Google Scholar]
- Salvador, R.; Fideghelli, C. La coltura protetta del pesco nell’Italia centro-meridionale, come tecnica per la coltivazione biologica. Italus. Hortus 2005, 12, 93–95. [Google Scholar]
- Arnegger, T.; Buchleither, S.; Mayr, U. Impact of a rain-roof-covering-system on the incidence of fungal diseases, quality parameters and solar radiation in organic apple production. In Proceedings of the 18th International Conference on Organic Fruit-Growing, Hohenheim, Germany, 19–21 February 2018; pp. 57–63. [Google Scholar]
- Mari, M.; Spadaro, D.; Casals, C.; Collina, M.; De Cal, A.; Usall, J. Postharvest Diseases of Stone Fruits. In Postharvest Pathology of Fresh Horticultural Produce; Palou, L., Smilanick, J.L., Eds.; CRC Press: Boca Raton, FL, USA, 2019; pp. 111–140. ISBN 9781138630833. [Google Scholar]
- Hoffmann, A.A.; Sgro, C.M. Climate change and evolutionary adaptation. Nature 2011, 470, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Roy, D.B.; Sparks, T.H. Phenology of British butterflies and climate change. Glob. Change Biol. 2000, 6, 407–416. [Google Scholar] [CrossRef]
- Forister, M.L.; Shapiro, A.M. Climatic trends and advancing spring flight of butterflies in lowland California. Glob. Change Biol. 2003, 9, 1130–1135. [Google Scholar] [CrossRef]
- Carroll, A.L.; Taylor, S.W.; Régnière, J.; Safranyik, L. Effects of climate change on range expansion by the mountain pine beetle in British Columbia. In Information Report-Pacific Forestry Centre; Canadian forest Service: Victoria, BC, Canada, 2004; pp. 223–232, BC-X-399. [Google Scholar]
- Battisti, A.; Stastny, M.; Buffo, E.; Larsson, S. A rapid altitudinal range expansion in the pine processionary moth produced by the 2003 climatic anomaly. Glob. Change Biol. 2006, 12, 662–671. [Google Scholar] [CrossRef]
- Gordo, O.; Sanz, J.J. Temporal trends in phenology of the honey bee Apis mellifera (L.) and the small white Pieris rapae (L.) in the Iberian Peninsula (1952–2004). Ecol. Entomol. 2006, 31, 261–268. [Google Scholar] [CrossRef]
- Harrington, R.; Clark, S.J.; Welham, S.J.; Verrier, P.J.; Denholm, C.H.; Hullé, M.; Maurice, D.; Rounsevell, M.D.; Cocu, N. Environmental change and the phenology of European aphids. Glob. Change Biol. 2007, 13, 1550–1564. [Google Scholar] [CrossRef]
- Dingemanse, N.J.; Kalkman, V.J. Changing temperature regimes have advanced the phenology of Odonata in the Netherlands. Ecol. Entomol. 2008, 33, 394–402. [Google Scholar] [CrossRef]
- Mukherjee, A.; Knoch, S.; Chouinard, G.; Tavares, J.R.; Dumont, M.J. Use of bio-based polymers in agricultural exclusion nets: A perspective. Biosyst. Eng. 2019, 180, 121–145. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Applied on | Active Ingredient | Trade Name | Year | No. of Treatments |
|---|---|---|---|---|---|
| Grapholita molesta | I | Chlorpyrifos methyl | ReldanTM | 2016 | 1 |
| I | Etofenprox | Trebon® UP | 2016 | 1 | |
| I | Phosmet | Spada® | 2017 | 1 | |
| I | Chlorpyrifos methyl | Pyrinex® | 2017 | 2▲ | |
| I | Etofenprox | Trebon® UP | 2017 | 2 | |
| I | Chlorpyrifos methyl | Runner® LO | 2017 | 1 | |
| Halyomorpha halys | I | Deltamethrin | Decis® | 2017 | 1 |
| Monilia spp. | N, C, I | Sulfur | Tiovit® | 2016 | 1 |
| N, C, I | Tebuconazole, sulfur | Tebusip® | 2016 | 2 | |
| N, C, I | Tebuconazole, sulfur | Tebusip® | 2017 | 1 | |
| N, C, I | Tebuconazole | Dedalus® 25 WDG | 2017 | 1 | |
| Monilia spp., Podosphaera pannosa | N, C, I | Sulfur | Tiovit® | 2017 | 2 |
| Cultivar | Treatment | Grapholita molesta (no.) | Damaged Nectarines (%) | ||
|---|---|---|---|---|---|
| 2016 | 2017 | 2016 | 2017 | ||
| Amiga* | N | 5.0 ± 2.1 b | 39.0 ± 9.2 b | 0.4 ± 0.2 | 0.6 ± 0.3 c |
| C | 22.7 ± 1.7 a | 160.3 ± 11.5 a | 4.2 ± 0.8 | 9.4 ± 0.9 a | |
| I | - | - | 2.1 ± 0.5 | 3.4 ± 0.6 b | |
| Fire Top® | N | 8.7 ± 3.3 b | 39.0 ± 4.6 b | 0.0 ± 0.0 | 0.4 ± 0.2 b |
| C | 38.3 ± 1.9 a | 174.7 ± 30.4 a | 0.1 ± 0.1 | 6.2 ± 0.9 a | |
| I | - | - | 0.1 ± 0.1 | 4.1 ± 0.7 a | |
| Cultivar | Treatment | Drosophila suzukii (no.) | Predators (no.) | ||
|---|---|---|---|---|---|
| 2016 | 2017 | 2016 | 2017 | ||
| Amiga* | N | 7.0 ± 1.0 b | 7.0 ± 2.3 b | 1.7 ± 1.2 b | 7.7 ± 2.0 b |
| C | 60.7 ± 3.4 a | 21.3 ± 1.8 a | 41.0 ± 3.0 a | 74.0 ± 2.0 a | |
| Fire Top® | N | 13.0 ± 7.6 b | 12.0 ± 2.5 b | 1.0 ± 0.6 b | 1.3 ± 0.3 b |
| C | 129.0 ± 25.2 a | 33.7 ± 6.4 a | 25.7 ± 1.7 a | 58.7 ± 9.6 a | |
| Cultivar | Treatment | Postharvest Rots Incidence (%) | |||
|---|---|---|---|---|---|
| After Storage | After Shelf Life | ||||
| 2016 | 2017 | 2016 | 2017 | ||
| Amiga* | N | 0.4 ± 0.3 | 0.0 ± 0.0 | 4.9 ± 1.6 b | 4.2 ± 1.9 b |
| C | 0.4 ± 0.5 | 0.6 ± 1.0 | 11.0 ± 4.2 a | 14.3 ± 2.5 a | |
| I | 0.0 ± 0.0 | 0.0 ± 0.0 | 3.3 ± 1.8 b | 6.8 ± 4.4 ab | |
| Fire Top® | N | 0.7 ± 0.9 | 2.2 ± 1.3 b | 15.9 ± 7.2 b | 14.1 ± 8.9 b |
| C | 4.0 ± 1.8 | 0.7 ± 1.3 b | 28.6 ± 8.4 a | 37.6 ± 2.3 a | |
| I | 1.6 ± 0.9 | 10.0 ± 4.8 a | 26.7 ± 11.3 a | 33.6 ± 15.2 ab | |
| Cultivar | Treatment | Color Index | Firmness (g cm−2) | Total Soluble Solids (°Brix) | Total Polyphenols (mgGAE 100 g−1) | Total Anthocyanins (mgC3G 100 g−1) |
|---|---|---|---|---|---|---|
| Amiga* | N | 30.8 ± 1.0 | 4.9 ± 0.1 | 8.2 ± 0.1 b | 33.8 ± 2.4 | 7.4 ± 2.0 |
| C | 30.8 ± 1.2 | 4.9 ± 0.1 | 8.6 ± 0.1 a | 31.1 ± 1.9 | 12.3 ± 1.8 | |
| I | 27.8 ± 1.1 | 5.2 ± 0.1 | 8.4 ± 0.1 ab | 29.0 ± 2.3 | 9.5 ± 2.2 | |
| Fire Top® | N | 36.7 ± 1.9 b | 4.2 ± 0.1 | 8.3 ± 0.1 c | 65.4 ± 6.9 | 9.7 ± 2.0 |
| C | 44.9 ± 1.6 a | 4.2 ± 0.1 | 8.7 ± 0.1 b | 51.5 ± 3.7 | 11.0 ± 2.3 | |
| I | 38.0 ± 1.6 b | 4.1 ± 0.1 | 8.9 ± 0.1 a | 47.9 ± 2.0 | 13.3 ± 2.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Candian, V.; Pansa, M.G.; Santoro, K.; Spadaro, D.; Briano, R.; Peano, C.; Tavella, L.; Tedeschi, R. First Multi-Target Application of Exclusion Net in Nectarine Orchards: Effectiveness against Pests and Impact on Beneficial Arthropods, Postharvest Rots and Fruit Quality. Insects 2021, 12, 210. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12030210
Candian V, Pansa MG, Santoro K, Spadaro D, Briano R, Peano C, Tavella L, Tedeschi R. First Multi-Target Application of Exclusion Net in Nectarine Orchards: Effectiveness against Pests and Impact on Beneficial Arthropods, Postharvest Rots and Fruit Quality. Insects. 2021; 12(3):210. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12030210
Chicago/Turabian StyleCandian, Valentina, Marco Giuseppe Pansa, Karin Santoro, Davide Spadaro, Rossella Briano, Cristiana Peano, Luciana Tavella, and Rosemarie Tedeschi. 2021. "First Multi-Target Application of Exclusion Net in Nectarine Orchards: Effectiveness against Pests and Impact on Beneficial Arthropods, Postharvest Rots and Fruit Quality" Insects 12, no. 3: 210. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12030210