
Lessons from Drosophila: Engineering Genetic Sexing Strains with Temperature-Sensitive Lethality for Sterile Insect Technique Applications



Abstract
:Simple Summary
Abstract
1. Introduction
1.1. The Sterile Insect Technique (SIT) and the Challenge of Sex Separation
1.2. Removal of Females, from Potential to Possible
2. Lessons from Drosophila melanogaster
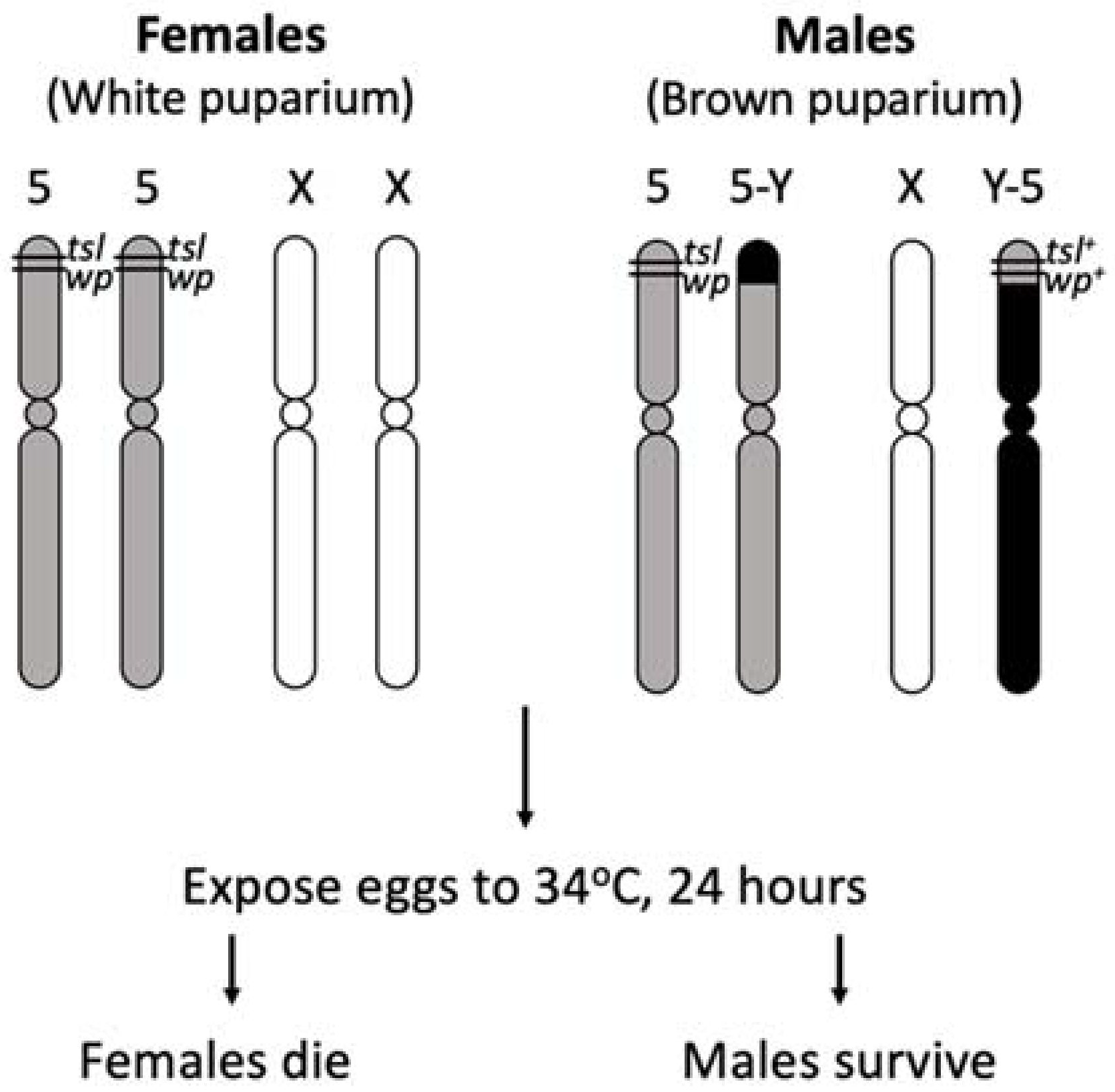
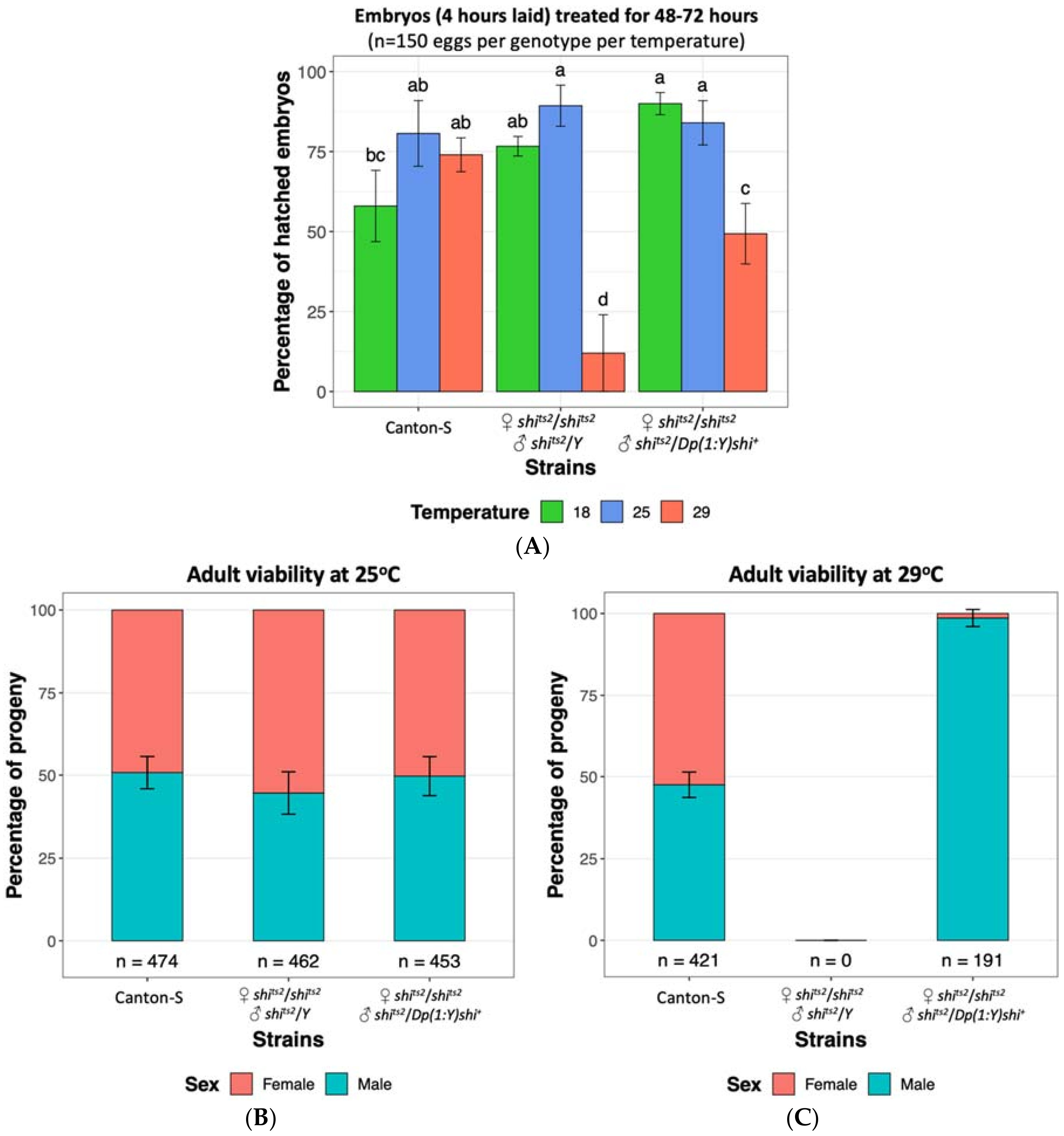
2.1. Shibire
2.2. Notch
2.2.1. Recessive Temperature Sensitive Lethal Notch Mutations
2.2.2. Dominant Temperature Sensitive Lethal Notch Mutations
2.3. RpII215
2.4. Pale
2.5. Transformer-2
2.6. Dsor1
2.7. CK2α
3. What’s Still to Come—Efficient Translocation Methods
4. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Robinson, A.S. Genetic Basis of the Sterile Insect Technique. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 95–114. [Google Scholar] [CrossRef]
- Klassen, W. Area-Wide Integrated Pest Management and the Sterile Insect Technique. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 39–68. [Google Scholar]
- Knipling, E. Possibilities of insect control or eradication through use of sexually sterile males. J. Econ. Entomol. 1955, 48, 459–462. [Google Scholar] [CrossRef]
- Beketov, M.A.; Kefford, B.J.; Schäfer, R.B.; Liess, M. Pesticides reduce regional biodiversity of stream invertebrates. Proc. Natl. Acad. Sci. USA 2013, 110, 11039–11043. [Google Scholar] [CrossRef] [Green Version]
- Nicolopoulou-Stamati, P.; Maipas, S.; Kotampasi, C.; Stamatis, P.; Hens, L. Chemical pesticides and human health: The urgent need for a new concept in agriculture. Front. Public Health 2016, 4, 148. [Google Scholar] [CrossRef] [Green Version]
- Wyss, J.H. Screwworm Eradication in the Americas. Ann. N. Y. Acad. Sci. 2000, 916, 186–193. [Google Scholar] [CrossRef]
- Enkerlin, W.; Gutiérrez-Ruelas, J.M.; Cortes, A.V.; Roldan, E.C.; Midgarden, D.; Lira, E.; López, J.L.Z.; Hendrichs, J.; Liedo, P.; Arriaga, F.J.T. Area Freedom in Mexico from Mediterranean Fruit Fly (Diptera: Tephritidae): A Review of Over 30 Years of a Successful Containment Program Using an Integrated Area-Wide SIT Approach. Fla. Entomol. 2015, 98, 665–681. [Google Scholar] [CrossRef] [Green Version]
- Marec, F.; Vreysen, M.J.B. Advances and Challenges of Using the Sterile Insect Technique for the Management of Pest Lepidoptera. Insects 2019, 10, 371. [Google Scholar] [CrossRef] [Green Version]
- Bourtzis, K.; Lees, R.S.; Hendrichs, J.; Vreysen, M.J.B. More than one rabbit out of the hat: Radiation, transgenic and symbiont-based approaches for sustainable management of mosquito and tsetse fly populations. Acta Trop. 2016, 157, 115–130. [Google Scholar] [CrossRef]
- Lees, R.S.; Gilles, J.R.L.; Hendrichs, J.; Vreysen, M.J.B.; Bourtzis, K. Back to the future: The sterile insect technique against mosquito disease vectors. Curr. Opin. Insect Sci. 2015, 10, 156–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sassu, F.; Nikolouli, K.; Caravantes, S.; Taret, G.; Pereira, R.; Vreysen, M.J.B.; Stauffer, C.; Caceres, C. Mass-Rearing of Drosophila suzukii for Sterile Insect Technique Application: Evaluation of Two Oviposition Systems. Insects 2019, 10, 448. [Google Scholar] [CrossRef] [Green Version]
- Suckling, D.M.; Conlong, D.E.; Carpenter, J.E.; Bloem, K.A.; Rendon, P.; Vreysen, M.J.B. Global range expansion of pest Lepidoptera requires socially acceptable solutions. Biol. Invasions 2017, 19, 1107–1119. [Google Scholar] [CrossRef] [Green Version]
- Parker, A.G. Mass-Rearing for Sterile Insect Release. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 209–232. [Google Scholar] [CrossRef]
- Cayol, J.P.; Causse, R.; Louis, C.; Barthes, J. Medfly Ceratitis capitata Wiedemann (Dipt., Trypetidae) as a rot vector in laboratory conditions. J. Appl. Entomol. 1994, 117, 338–343. [Google Scholar] [CrossRef]
- McInnis, D.O.; Tam, S.; Grace, C.; Miyashita, D. Population suppression and sterility rates induced by variable sex ratio, Sterile Insect Releases of Ceratitis capitata (Diptera: Tephritidae) in Hawaii. Ann. Entomol. Soc. Am. 1994, 87, 231–240. [Google Scholar] [CrossRef]
- Rendón, P.; McInnis, D.; Lance, D.; Stewart, J. Medfly (Diptera:Tephritidae) Genetic Sexing: Large-scale field comparison of males-only and bisexual sterile fly releases in Guatemala. J. Econ. Entomol. 2004, 97, 1547–1553. [Google Scholar] [CrossRef]
- Caceres, C. Mass rearing of temperature sensitive genetic sexing strains in the Mediterranean fruit fly (Ceratitis capitata). Genetica 2002, 116, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Robinson, A.S. Genetic Sexing Strains in Medfly, Ceratitis Capitata, Sterile Insect Technique Programmes. Genetica 2002, 116, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Bakri, A.; Mehta, K.; Lance, D.; Dyck, V.; Hendrichs, J.; Robinson, A. Sterilizing Insects with Ionizing Radiation. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 233–268. [Google Scholar] [CrossRef]
- Bushland, R.C.; Hopkins, D.E. Experiments with Screw-Worm Flies Sterilized by X-Rays. J. Econ. Entomol. 1951, 44, 725–731. [Google Scholar] [CrossRef]
- Bushland, R.C. Male sterilization for the control of insects. In Advances in Pest Control Research; Metcalf, R.L., Ed.; Interscience Publishers: New York, NY, USA, 1960; pp. 1–25. [Google Scholar]
- Papathanos, P.A.; Bourtzis, K.; Tripet, F.; Bossin, H.; Virginio, J.F.; Capurro, M.L.; Pedrosa, M.C.; Guindo, A.; Sylla, L.; Coulibaly, M.B.; et al. A perspective on the need and current status of efficient sex separation methods for mosquito genetic control. Parasites Vectors 2018, 11, 654. [Google Scholar] [CrossRef]
- Zheng, X.; Zhang, D.; Li, Y.; Yang, C.; Wu, Y.; Liang, X.; Liang, Y.; Pan, X.; Hu, L.; Sun, Q.; et al. Incompatible and sterile insect techniques combined eliminate mosquitoes. Nature 2019, 572, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Bellini, R.; Medici, A.; Puggioli, A.; Balestrino, F.; Carrieri, M. Pilot Field Trials With Aedes albopictus Irradiated Sterile Males in Italian Urban Areas. J. Med. Entomol. 2013, 50, 317–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seck, M.T.; Pagabeleguem, S.; Bassene, M.D.; Fall, A.G.; Diouf, T.A.R.; Sall, B.; Vreysen, M.J.B.; Rayaissé, J.-B.; Takac, P.; Sidibé, I.; et al. Quality of Sterile Male Tsetse after Long Distance Transport as Chilled, Irradiated Pupae. PLoS Negl. Trop. Dis. 2015, 9, e0004229. [Google Scholar] [CrossRef] [Green Version]
- Franz, G. Genetic Sexing Strains in Mediterranean Fruit Fly, an Example for Other Species Amenable to Large-Scale Rearing for the Sterile Insect Technique. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 427–451. [Google Scholar] [CrossRef]
- McCombs, S.D.; Saul, S.H. Linkage Analysis of Five New Genetic Markers of the Oriental Fruit Fly, Bactrocera dorsalis (Diptera: Tephritidae). J. Hered. 1992, 83, 199–203. [Google Scholar] [CrossRef] [PubMed]
- McInnis, D.O.; Tam, S.; Lim, R.; Komatsu, J.; Kurashima, R.; Albrecht, C. Development of a Pupal Color-Based Genetic Sexing Strain of the Melon Fly, Bactrocera cucurbitae (Coquillett) (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 2004, 97, 1026–1033. [Google Scholar] [CrossRef] [Green Version]
- Zepeda-Cisneros, C.S.; Meza-Hernandez, J.S.; Garcia-Martinez, V.; Ibanez-Palacios, J.; Zacharopoulou, A.; Franz, G. Development, genetic and cytogenetic analyses of genetic sexing strains of the Mexican fruit fly, Anastrepha ludens Loew (Diptera: Tephritidae). BMC Genet. 2014, 15, S1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Augustinos, A.A.; Misbah-ul-Haq, M.; Carvalho, D.O.; de la Fuente, L.D.; Koskinioti, P.; Bourtzis, K. Irradiation induced inversions suppress recombination between the M locus and morphological markers in Aedes aegypti. BMC Genet. 2020, 21, 142. [Google Scholar] [CrossRef]
- Koskinioti, P.; Augustinos, A.A.; Carvalho, D.O.; Misbah-ul-Haq, M.; Pillwax, G.; de la Fuente, L.D.; Salvador-Herranz, G.; Herrero, R.A.; Bourtzis, K. Genetic sexing strains for the population suppression of the mosquito vector Aedes aegypti. Philos. Trans. R. Soc. B Biol. Sci. 2021, 376, 20190808. [Google Scholar] [CrossRef]
- Rössler, Y. The Genetics of the Mediterranean Fruit Fly: A “White Pupae” Mutant. Ann. Entomol. Soc. Am. 1979, 72, 583–585. [Google Scholar] [CrossRef]
- Caceres, C.; Cayol, J.-P.; Enkerlin, W.R.; Franz, G.; Hendrichs, J.; Robinson, A.S. Comparison of Mediterranean fruit fly (Ceratitis capitata) (Tephritidae) bisexual and genetic sexing strains: Development, evaluation and economics. In Proceedings of the 6th International Symposium on Fruit Flies of Economic Importance, Stellenbosch, South Africa, 6–10 May 2002; pp. 367–381. [Google Scholar]
- Franz, G.; Gencheva, E.; Kerremans, P. Improved stability of genetic sex-separation strains for the Mediterranean fruit fly, Ceratitis capitata. Genome 1994, 37, 72–82. [Google Scholar] [CrossRef]
- Franz, G. Development and application of genetic sexing systems for the Mediterranean fruit fly based on a temperature sensitive lethal. In Fruit Fly Pest, A World Assessment of Their Biology and Management; McPheron, B.A., Steck, G.J., Eds.; St. Lucia Press: Miami, FL, USA, 1996; pp. 185–191. [Google Scholar]
- Augustinos, A.A.; Targovska, A.; Cancio-Martinez, E.; Schorn, E.; Franz, G.; Cáceres, C.; Zacharopoulou, A.; Bourtzis, K. Ceratitis capitata genetic sexing strains: Laboratory evaluation of strains from mass-rearing facilities worldwide. Entomol. Exp. Appl. 2017, 164, 305–317. [Google Scholar] [CrossRef] [Green Version]
- Kerremans, P.; Franz, G. Isolation and cytogenetic analyses of genetic sexing strains for the medfly, Ceratitis capitata. Theor. Appl. Genet. 1995, 91, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Franz, G. Recombination Between Homologous Autosomes in Medfly (Ceratitis Capitata) Males: Type-1 Recombination and the Implications for the Stability of Genetic Sexing Strains. Genetica 2002, 116, 73–84. [Google Scholar] [CrossRef]
- Sim, S.B.; Ruiz-Arce, R.; Barr, N.B.; Geib, S.M. A New Diagnostic Resource for Ceratitis capitata Strain Identification Based on QTL Mapping. G3 Genes Genomes Genet. 2017, 7, 3637–3647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, C.M.; Aumann, R.A.; Whitehead, M.A.; Nikolouli, K.; Leveque, G.; Gouvi, G.; Fung, E.; Reiling, S.J.; Djambazian, H.; Hughes, M.A.; et al. White pupae phenotype of tephritids is caused by parallel mutations of a MFS transporter. Nat. Commun. 2021, 12, 491. [Google Scholar] [CrossRef] [PubMed]
- Aumann, R.A.; Häcker, I.; Schetelig, M.F. Female-to-male sex conversion in Ceratitis capitata by CRISPR/Cas9 HDR-induced point mutations in the sex determination gene transformer-2. Sci. Rep. 2020, 10, 18611. [Google Scholar] [CrossRef] [PubMed]
- Choo, A.; Fung, E.; Chen, I.Y.; Saint, R.; Crisp, P.; Baxter, S.W. Precise single base substitution in the shibire gene by CRISPR/Cas9-mediated homology directed repair in Bactrocera tryoni. BMC Genet. 2020, 21, 127. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Handler, A.M. Temperature-dependent sex-reversal by a transformer-2 gene-edited mutation in the spotted wing drosophila, Drosophila suzukii. Sci. Rep. 2017, 7, 12363. [Google Scholar] [CrossRef] [PubMed]
- Robinson, A.S. Mutations and their use in insect control. Mutat. Res. 2002, 511, 113–132. [Google Scholar] [CrossRef]
- Heinrich, J.C.; Scott, M.J. A repressible female-specific lethal genetic system for making transgenic insect strains suitable for a sterile-release program. Proc. Natl. Acad. Sci. USA 2000, 97, 8229–8232. [Google Scholar] [CrossRef] [Green Version]
- Horn, C.; Wimmer, E.A. A transgene-based, embryo-specific lethality system for insect pest management. Nat. Biotechnol. 2003, 21, 64–70. [Google Scholar] [CrossRef]
- Thomas, D.D.; Donnelly, C.A.; Wood, R.J.; Alphey, L.S. Insect Population Control Using a Dominant, Repressible, Lethal Genetic System. Science 2000, 287, 2474–2476. [Google Scholar] [CrossRef]
- Ogaugwu, C.E.; Schetelig, M.F.; Wimmer, E.A. Transgenic sexing system for Ceratitis capitata (Diptera: Tephritidae) based on female-specific embryonic lethality. Insect Biochem. Mol. Biol. 2013, 43, 1–8. [Google Scholar] [CrossRef]
- Schetelig, M.F.; Handler, A.M. A transgenic embryonic sexing system for Anastrepha suspensa (Diptera: Tephritidae). Insect Biochem. Mol. Biol. 2012, 42, 790–795. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Scott, M.J. A transgenic embryonic sexing system for the Australian sheep blow fly Lucilia cuprina. Sci. Rep. 2015, 5, 16090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, K.P.; Teo, T.M.; Ho, T.X.; Le, V.S. Mechanisms of sex determination and transmission ratio distortion in Aedes aegypti. Parasites Vectors 2016, 9, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galizi, R.; Doyle, L.A.; Menichelli, M.; Bernardini, F.; Deredec, A.; Burt, A.; Stoddard, B.L.; Windbichler, N.; Crisanti, A. A synthetic sex ratio distortion system for the control of the human malaria mosquito. Nat. Commun. 2014, 5, 3977. [Google Scholar] [CrossRef] [Green Version]
- Bernardini, F.; Galizi, R.; Menichelli, M.; Papathanos, P.-A.; Dritsou, V.; Marois, E.; Crisanti, A.; Windbichler, N. Site-specific genetic engineering of the Anopheles gambiae Y chromosome. Proc. Natl. Acad. Sci. USA 2014, 111, 7600. [Google Scholar] [CrossRef] [Green Version]
- Catteruccia, F.; Benton, J.P.; Crisanti, A. An Anopheles transgenic sexing strain for vector control. Nat. Biotechnol. 2005, 23, 1414–1417. [Google Scholar] [CrossRef]
- Condon, K.C.; Condon, G.C.; Dafa’alla, T.H.; Fu, G.; Phillips, C.E.; Jin, L.; Gong, P.; Alphey, L. Genetic sexing through the use of Y-linked transgenes. Insect Biochem. Mol. Biol. 2007, 37, 1168–1176. [Google Scholar] [CrossRef] [PubMed]
- Meza, J.S.; Schetelig, M.F.; Zepeda-Cisneros, C.S.; Handler, A.M. Male-specific Y-linked transgene markers to enhance biologically-based control of the Mexican fruit fly, Anastrepha ludens (Diptera: Tephritidae). BMC Genet. 2014, 15, S4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schetelig, M.F.; Handler, A.M. Y-linked markers for improved population control of the tephritid fruit fly pest, Anastrepha suspensa. Adv. Biochem. Eng. Biotechnol. 2013, 136, 123–133. [Google Scholar] [CrossRef]
- Yan, Y.; Scott, M.J. Building a transgenic sexing strain for genetic control of the Australian sheep blow fly Lucilia cuprina using two lethal effectors. BMC Genom. Data 2020, 21, 141. [Google Scholar] [CrossRef]
- Fu, G.; Lees, R.S.; Nimmo, D.; Aw, D.; Jin, L.; Gray, P.; Berendonk, T.U.; White-Cooper, H.; Scaife, S.; Kim Phuc, H.; et al. Female-specific flightless phenotype for mosquito control. Proc. Natl. Acad. Sci. USA 2010, 107, 4550–4554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panjwani, A.; Wilson, A. What Is Stopping the Use of Genetically Modified Insects for Disease Control? PLoS Pathog. 2016, 12, e1005830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, S.R.; Maselko, M.; Upadhyay, A.; Smanski, M.J. Genetic engineering of sex chromosomes for batch cultivation of non-transgenic, sex-sorted males. PLoS Genet. 2020, 16, e1009180. [Google Scholar] [CrossRef] [PubMed]
- Sahara, K.; Yoshido, A.; Traut, W. Sex chromosome evolution in moths and butterflies. Chromosome Res. 2012, 20, 83–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marec, F.; Neven, L.G.; Robinson, A.S.; Vreysen, M.; Goldsmith, M.R.; Nagaraju, J.; Franz, G. Development of Genetic Sexing Strains in Lepidoptera: From Traditional to Transgenic Approaches. J. Econ. Entomol. 2005, 98, 248–259. [Google Scholar] [CrossRef]
- Ma, S.; Wang, X.; Fei, J.; Liu, Y.; Duan, J.; Wang, F.; Xu, H.; Zhao, P.; Xia, Q. Genetic marking of sex using a W chromosome-linked transgene. Insect. Biochem. Mol. Biol. 2013, 43, 1079–1086. [Google Scholar] [CrossRef] [PubMed]
- Tamura, T.; Thibert, C.; Royer, C.; Kanda, T.; Abraham, E.; Kamba, M.; Komoto, N.; Thomas, J.L.; Mauchamp, B.; Chavancy, G.; et al. Germline transformation of the silkworm Bombyx mori L. using a piggyBac transposon-derived vector. Nat. Biotechnol. 2000, 18, 81–84. [Google Scholar] [CrossRef] [PubMed]
- Marec, F.; Neven, L.G.; Fukova, I. Developing Transgenic Sexing Strains for the Release of Non-Transgenic Sterile Male Codling Moths Cydia pomonella. In Area-Wide Control of Insect Pests; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, The Netherland, 2007. [Google Scholar] [CrossRef]
- Bai, X.; Zeng, T.; Ni, X.Y.; Su, H.A.; Huang, J.; Ye, G.Y.; Lu, Y.Y.; Qi, Y.X. CRISPR/Cas9-mediated knockout of the eye pigmentation gene white leads to alterations in color of head spots in the oriental fruit fly, Bactrocera dorsalis. Insect Mol. Biol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Choo, A.; Crisp, P.; Saint, R.; O’Keefe, L.V.; Baxter, S.W. CRISPR/Cas9-mediated mutagenesis of the white gene in the tephritid pest Bactrocera tryoni. J. Appl. Entomol. 2018, 142, 52–58. [Google Scholar] [CrossRef]
- Kistler, K.E.; Vosshall, L.B.; Matthews, B.J. Genome engineering with CRISPR-Cas9 in the mosquito Aedes aegypti. Cell. Rep. 2015, 11, 51–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Akbari, O.S.; White, B.J. Highly Efficient Site-Specific Mutagenesis in Malaria Mosquitoes Using CRISPR. G3 Genes Genomes Genet. 2018, 8, 653–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulo, D.F.; Williamson, M.E.; Arp, A.P.; Li, F.; Sagel, A.; Skoda, S.R.; Sanchez-Gallego, J.; Vasquez, M.; Quintero, G.; Perez de Leon, A.A.; et al. Specific Gene Disruption in the Major Livestock Pests Cochliomyia hominivorax and Lucilia cuprina Using CRISPR/Cas9. G3 Genes Genomes Genet. 2019, 9, 3045–3055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, E.B.; Bacher, F. Methods of feeding ethyl methane sulphonate (EMS) to Drosophila males. Drosoph. Inf. Serv. 1968, 43, 193. [Google Scholar]
- Suzuki, D.T. Temperature-Sensitive Mutations in Drosophila melanogaster. Science 1970, 170, 695–706. [Google Scholar] [CrossRef]
- Suzuki, D.T.; Grigliatti, T.; Williamson, R. Temperature-Sensitive Mutations in Drosophila melanogaster, VII. A Mutation (parats) Causing Reversible Adult Paralysis. Proc. Natl. Acad. Sci. USA 1971, 68, 890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grigliatti, T.A.; Hall, L.; Rosenbluth, R.; Suzuki, D.T. Temperature-sensitive mutations in Drosophila melanogaster. Mol. Gen. Genet. 1973, 120, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Hanna, D.E.; Rethinaswamy, A.; Glover, C.V. Casein kinase II is required for cell cycle progression during G1 and G2/M in Saccharomyces cerevisiae. J. Biol. Chem. 1995, 270, 25905–25914. [Google Scholar] [CrossRef] [Green Version]
- Kuntamalla, P.P.; Kunttas-Tatli, E.; Karandikar, U.; Bishop, C.P.; Bidwai, A.P. Drosophila protein kinase CK2 is rendered temperature-sensitive by mutations of highly conserved residues flanking the activation segment. Mol. Cell. Biochem. 2009, 323, 49–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poodry, C.A.; Hall, L.; Suzuki, D.T. Developmental properties of shibirets1: A pleiotropic mutation affecting larval and adult locomotion and development. Dev. Biol. 1973, 32, 373–386. [Google Scholar] [CrossRef]
- Kim, H.J.; Lee, H.J.; Kim, H.; Cho, S.W.; Kim, J.-S. Targeted genome editing in human cells with zinc finger nucleases constructed via modular assembly. Genome Res. 2009, 19, 1279–1288. [Google Scholar] [CrossRef] [Green Version]
- Ramaswami, M.; Rao, S.; van der Bliek, A.; Kelly, R.B.; Krishnan, K.S. Genetic Studies on Dynamin Function in Drosophila. J. Neurogenet. 1993, 9, 73–87. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, K.S.; Rikhy, R.; Rao, S.; Shivalkar, M.; Mosko, M.; Narayanan, R.; Etter, P.; Estes, P.S.; Ramaswami, M. Nucleoside diphosphate kinase, a source of GTP, is required for dynamin-dependent synaptic vesicle recycling. Neuron 2001, 30, 197–210. [Google Scholar] [CrossRef] [Green Version]
- Narayanan, R.; Leonard, M.; Song, B.D.; Schmid, S.L.; Ramaswami, M. An internal GAP domain negatively regulates presynaptic dynamin in vivo: A two-step model for dynamin function. J. Cell. Biol. 2005, 169, 117–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shellenbarger, D.L.; Mohler, J.D. Temperature-sensitive periods and autonomy of pleiotropic effects of l(1)Nts1, a conditional notch lethal in Drosophila. Dev. Biol. 1978, 62, 432–446. [Google Scholar] [CrossRef]
- Ge, X.; Hannan, F.; Xie, Z.; Feng, C.; Tully, T.; Zhou, H.; Xie, Z.; Zhong, Y. Notch signaling in Drosophila long-term memory formation. Proc. Natl. Acad. Sci. USA 2004, 101, 10172–10176. [Google Scholar] [CrossRef] [Green Version]
- Portin, P.; Sirén, M. Timing of temperature-sensitive period for lethality of Abruptex mutations of Drosophila melanogaster. Hereditas 1976, 84, 109–116. [Google Scholar] [CrossRef]
- Welshons, W.J.; von Halle, E.S. Pseudoallelism at the notch locus in drosophila. Genetics 1962, 47, 743–759. [Google Scholar] [CrossRef] [PubMed]
- Foster, G.G. Temperature-sensitive mutations in Drosophila melanogaster: XIII. Temperature-sensitive periods of the lethal and morphological phenotypes of selected combinations of notch-locus mutations. Dev. Biol. 1973, 32, 282–296. [Google Scholar] [CrossRef]
- Fryxell, K.J.; Miller, T.A. Autocidal Biological Control: A General Strategy for Insect Control Based on Genetic Transformation with a Highly Conserved Gene. J. Econ. Entomol. 1995, 88, 1221–1232. [Google Scholar] [CrossRef]
- Lyman, D.; Young, M.W. Further evidence for function of the Drosophila Notch protein as a transmembrane receptor. Proc. Natl. Acad. Sci. USA 1993, 90, 10395–10399. [Google Scholar] [CrossRef] [Green Version]
- Mortin, M.A.; Kaufman, T.C. Development effects of a temperature-sensitive RNA polymerase II mutation in Drosophila melanogaster. Dev. Biol. 1984, 103, 343–354. [Google Scholar] [CrossRef]
- Pendleton, R.G.; Rasheed, A.; Sardina, T.; Tully, T.; Hillman, R. Effects of tyrosine hydroxylase mutants on locomotor activity in Drosophila: A study in functional genomics. Behav. Genet. 2002, 32, 89–94. [Google Scholar] [CrossRef]
- Belote, J.M.; Lucchesi, J.C. Control of X chromosome transcription by the maleless gene in Drosophila. Nature 1980, 285, 573–575. [Google Scholar] [CrossRef]
- Hsu, J.C.; Perrimon, N. A temperature-sensitive MEK mutation demonstrates the conservation of the signaling pathways activated by receptor tyrosine kinases. Genes Dev. 1994, 8, 2176–2187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.S.; Obar, R.A.; Schroeder, C.C.; Austin, T.W.; Poodry, C.A.; Wadsworth, S.C.; Vallee, R.B. Multiple forms of dynamin are encoded by shibire, a Drosophila gene involved in endocytosis. Nature 1991, 351, 583–586. [Google Scholar] [CrossRef] [PubMed]
- van der Bliek, A.M.; Meyerowitz, E.M. Dynamin-like protein encoded by the Drosophila Shibire gene associated with vesicular traffic. Nature 1991, 351, 411–414. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.G.; Shurland, D.-L.; Meyerowitz, E.M.; Bargmann, C.I.; van der Bliek, A.M. A dynamin GTPase mutation causes a rapid and reversible temperature-inducible locomotion defect in C.elegans. Proc. Natl. Acad. Sci. USA 1997, 94, 10438–10443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damke, H.; Baba, T.; van der Bliek, A.M.; Schmid, S.L. Clathrin-independent pinocytosis is induced in cells overexpressing a temperature-sensitive mutant of dynamin. J. Cell Biol. 1995, 131, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Grant, D.; Unadkat, S.; Katzen, A.; Krishnan, K.S.; Ramaswami, M. Probable mechanisms underlying interallelic complementation and temperature-sensitivity of mutations at the shibire locus of Drosophila melanogaster. Genetics 1998, 149, 1019–1030. [Google Scholar]
- Poodry, C.A. Shibire, a neurogenic mutant of Drosophila. Dev. Biol. 1990, 138, 464–472. [Google Scholar] [CrossRef]
- Kim, Y.-T.; Wu, C.-F. Allelic Interactions at the Shibire Locus of Drosophila: Effects on Behavior. J. Neurogenet. 1990, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Wharton, K.A.; Johansen, K.M.; Xu, T.; Artavanis-Tsakonas, S. Nucleotide sequence from the neurogenic locus Notch implies a gene product that shares homology with proteins containing EGF-like repeats. Cell 1985, 43, 567–581. [Google Scholar] [CrossRef]
- Kidd, S.; Kelley, M.R.; Young, M.W. Sequence of the notch locus of Drosophila melanogaster: Relationship of the encoded protein to mammalian clotting and growth factors. Mol. Cell. Biol. 1986, 6, 3094–3108. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Adam, J.C.; Montell, D. Spatially localized Kuzbanian required for specific activation of Notch during border cell migration. Dev. Biol. 2007, 301, 532–540. [Google Scholar] [CrossRef] [Green Version]
- Brennan, C.A.; Moses, K. Determination of Drosophila photoreceptors: Timing is everything. Cell. Mol. Life Sci. CMLS 2000, 57, 195–214. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Reyes, A.; St Johnston, D. Patterning of the follicle cell epithelium along the anterior-posterior axis during Drosophila oogenesis. Development 1998, 125, 2837–2846. [Google Scholar] [PubMed]
- Ward, E.J.; Zhou, X.; Riddiford, L.M.; Berg, C.A.; Ruohola-Baker, H. Border of Notch activity establishes a boundary between the two dorsal appendage tube cell types. Dev. Biol. 2006, 297, 461–470. [Google Scholar] [CrossRef] [Green Version]
- Shellenbarger, D.L.; Mohler, J.D. Temperature-sensitive mutations of the notch locus in Drosophila melanogaster. Genetics 1975, 81, 143–162. [Google Scholar] [CrossRef]
- de Celis, J.F.; Garcia-Bellido, A. Modifications of the notch function by Abruptex mutations in Drosophila melanogaster. Genetics 1994, 136, 183–194. [Google Scholar] [PubMed]
- Xu, T.; Caron, L.A.; Fehon, R.G.; Artavanis-Tsakonas, S. The involvement of the Notch locus in Drosophila oogenesis. Development 1992, 115, 913. [Google Scholar]
- Kelley, M.R.; Kidd, S.; Deutsch, W.A.; Young, M.W. Mutations altering the structure of epidermal growth factor-like coding sequences at the Drosophila Notch locus. Cell 1987, 51, 539–548. [Google Scholar] [CrossRef]
- Gordon, W.R.; Arnett, K.L.; Blacklow, S.C. The molecular logic of Notch signaling—A structural and biochemical perspective. J. Cell Sci. 2008, 121, 3109–3119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, G.G.; Suzuki, D.T. Temperature-sensitive mutations in Drosophila melanogaster. IV. A mutation affecting eye facet arrangement in a polarized manner. Proc. Natl. Acad. Sci. USA 1970, 67, 738–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortin, M.A.; Kaufman, T.C. Developmental genetics of a temperature sensitive RNA polymerase II mutation in Drosophila melanogaster. Mol. Gen. Genet. 1982, 187, 120–125. [Google Scholar] [CrossRef]
- Ingles, C.J.; Biggs, J.; Wong, J.K.; Weeks, J.R.; Greenleaf, A.L. Identification of a structural gene for a RNA polymerase II polypeptide in Drosophila melanogaster and mammalian species. Proc. Natl. Acad. Sci. USA 1983, 80, 3396–3400. [Google Scholar] [CrossRef] [Green Version]
- Krasnoselskaya, I.; Huang, J.; Jones, T.; Dezan, C.; Mortin, M.A. Selection and analysis of rare second-site suppressors of Drosophila RNA polymerase II mutations. Mol. Gen. Genet. 1998, 258, 457–465. [Google Scholar] [CrossRef]
- Severinov, K.; Mustaev, A.; Kukarin, A.; Muzzin, O.; Bass, I.; Darst, S.A.; Goldfarb, A. Structural Modules of the Large Subunits of RNA Polymerase: Introducing archaebacterial and chloroplast split sites in the β and β′ subunits of Escherichia coli RNA polymerase. J. Biol. Chem. 1996, 271, 27969–27974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cramer, P.; Bushnell, D.A.; Kornberg, R.D. Structural Basis of Transcription: RNA Polymerase II at 2.8 Ångstrom Resolution. Science 2001, 292, 1863. [Google Scholar] [CrossRef] [Green Version]
- Neckameyer, W.S.; White, K. Drosophila tyrosine hydroxylase is encoded by the pale locus. J. Neurogenet. 1993, 8, 189–199. [Google Scholar] [CrossRef]
- Swinderen, B.V.; Andretic, R. Dopamine in Drosophila: Setting arousal thresholds in a miniature brain. Proc. R Soc. B Biol. Sci. 2011, 278, 906–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Dartevelle, L.; Yuan, C.; Wei, H.; Wang, Y.; Ferveur, J.-F.; Guo, A. Reduction of Dopamine Level Enhances the Attractiveness of Male Drosophila to Other Males. PLoS ONE 2009, 4, e4574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jürgens, G.; Wieschaus, E.; Nüsslein-Volhard, C.; Kluding, H. Mutations affecting the pattern of the larval cuticle in Drosophila melanogaster: II. Zygotic loci on the third chromosome. Wilehm Roux Arch. Dev. Biol. 1984, 193, 283–295. [Google Scholar] [CrossRef]
- Bachtrog, D.; Mank, J.E.; Peichel, C.L.; Kirkpatrick, M.; Otto, S.P.; Ashman, T.-L.; Hahn, M.W.; Kitano, J.; Mayrose, I.; Ming, R.; et al. Sex Determination: Why So Many Ways of Doing It? PLoS Biol. 2014, 12, e1001899. [Google Scholar] [CrossRef] [Green Version]
- Tian, M.; Maniatis, T. A splicing enhancer complex controls alternative splicing of doublesex pre-mRNA. Cell 1993, 74, 105–114. [Google Scholar] [CrossRef]
- Amrein, H.; Gorman, M.; Nöthiger, R. The sex-determining gene tra-2 of Drosophila encodes a putative RNA binding protein. Cell 1988, 55, 1025–1035. [Google Scholar] [CrossRef]
- Hedley, M.L.; Maniatis, T. Sex-specific splicing and polyadenylation of dsx pre-mRNA requires a sequence that binds specifically to tra-2 protein in vitro. Cell 1991, 65, 579–586. [Google Scholar] [CrossRef]
- Hoshijima, K.; Inoue, K.; Higuchi, I.; Sakamoto, H.; Shimura, Y. Control of doublesex alternative splicing by transformer and transformer-2 in Drosophila. Science 1991, 252, 833. [Google Scholar] [CrossRef]
- Liu, G.; Wu, Q.; Li, J.; Zhang, G.; Wan, F. RNAi-Mediated Knock-Down of transformer and transformer 2 to Generate Male-Only Progeny in the Oriental Fruit Fly, Bactrocera dorsalis (Hendel). PLoS ONE 2015, 10, e0128892. [Google Scholar] [CrossRef] [Green Version]
- Thongsaiklaing, T.; Nipitwattanaphon, M.; Ngernsiri, L. The transformer2 gene of the pumpkin fruit fly, Bactrocera tau (Walker), functions in sex determination, male fertility and testis development. Insect Mol. Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Salvemini, M.; Robertson, M.; Aronson, B.; Atkinson, P.; Polito, C.; Saccone, G. Ceratitis capitata transformer- 2 gene is required to establish and maintain the auto-regulation of Cctra, the master gene for female sex determination. Int. J. Dev. Biol. 2009, 53, 109–120. [Google Scholar] [CrossRef] [Green Version]
- Schetelig, M.F.; Milano, A.; Saccone, G.; Handler, A.M. Male only progeny in Anastrepha suspensa by RNAi-induced sex reversion of chromosomal females. Insect Biochem. Mol. Biol. 2012, 42, 51–57. [Google Scholar] [CrossRef]
- Belote, J.M.; Baker, B.S. Sex determination in Drosophila melanogaster: Analysis of transformer-2, a sex-transforming locus. Proc. Natl. Acad. Sci. USA 1982, 79, 1568. [Google Scholar] [CrossRef] [Green Version]
- Belote, J.M.; Lucchesi, J.C. Male-specific lethal mutations of Drosophila melanogaster. Genetics 1980, 96, 165–186. [Google Scholar] [PubMed]
- Amrein, H.; Maniatis, T.; Nothiger, R. Alternatively spliced transcripts of the sex determinaing gene tra-2 of Drosophila encode functional proteins of different size. EMBO 1990, 9, 3619–3629. [Google Scholar] [CrossRef]
- Belote, J.M.; Handler, A.M.; Wolfner, M.F.; Livak, K.J.; Baker, B.S. Sex-specific regulation of yolk protein gene expression in Drosophila. Cell 1985, 40, 339–348. [Google Scholar] [CrossRef]
- Bandziulis, R.J.; Swanson, M.S.; Dreyfuss, G. RNA-binding proteins as developmental regulators. Genes Dev. 1989, 3, 431–437. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, L.; Inoue, Y.H.; Yoo, M.A.; Mizuno, M.; Hata, M.; Lim, Y.M.; Adachi-Yamada, T.; Ryo, H.; Masamune, Y.; Nishida, Y. A protein kinase similar to MAP kinase activator acts downstream of the raf kinase in Drosophila. Cell 1993, 72, 407–414. [Google Scholar] [CrossRef]
- Carr, A.M.; MacNeill, S.A.; Hayles, J.; Nurse, P. Molecular cloning and sequence analysis of mutant alleles of the fission yeast cdc2 protein kinase gene: Implications for cdc2+ protein structure and function. Mol. Gen. Genet. MGG 1989, 218, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, M.; Arbet, S.; Bishop, C.P.; Bidwai, A.P. Drosophila protein kinase CK2: Genetics, regulatory complexity and emerging roles during development. Pharmaceuticals 2017, 20, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nirmala, X.; Zimowska, G.J.; Handler, A.M. Characterization of the proteasomebeta2 subunit gene and its mutant allele in the tephritid fruit fly pest, Anastrepha suspensa. Insect. Mol. Biol. 2009, 18, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Zacharopoulou, A.; Augustinos, A.A.; Drosopoulou, E.; Tsoumani, K.T.; Gariou-Papalexiou, A.; Franz, G.; Mathiopoulos, K.D.; Bourtzis, K.; Mavragani-Tsipidou, P. A review of more than 30 years of cytogenetic studies of Tephritidae in support of sterile insect technique and global trade. Entomol. Exp. Appl. 2017, 164, 204–225. [Google Scholar] [CrossRef] [Green Version]
- Fisher, K.; Caceres, C. A filter rearing system for mass reared genetic sexing strains of Mediterranean fruit fly (Diptera: Tephritidae). In Area-Wide Control of Fruit Flies and Other Insect Pests Joint Proceedings of the International Conference on Area-Wide Control of Insect Pests and the Fifth International Symposium on Fruit Flies of Economic Importance; Tan, K.-H., Ed.; Penerbit Universiti Sains: Penang, Malaysia, 2000; pp. 543–550. [Google Scholar]
- Choi, P.S.; Meyerson, M. Targeted genomic rearrangements using CRISPR/Cas technology. Nat. Commun. 2014, 5, 3728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, R.; Martin, M.C.; Garcia, A.; Cigudosa, J.C.; Ramirez, J.C.; Rodriguez-Perales, S. Engineering human tumour-associated chromosomal translocations with the RNA-guided CRISPR–Cas9 system. Nat. Commun. 2014, 5, 3964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagutina, I.V.; Valentine, V.; Picchione, F.; Harwood, F.; Valentine, M.B.; Villarejo-Balcells, B.; Carvajal, J.J.; Grosveld, G.C. Modeling of the human alveolar rhabdomyosarcoma Pax3-Foxo1 chromosome translocation in mouse myoblasts using CRISPR-Cas9 nuclease. PLoS Genet. 2015, 11, e1004951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Li, M.; Feng, X.; Guang, S. Targeted Chromosomal Translocations and Essential Gene Knockout Using CRISPR/Cas9 Technology in Caenorhabditis elegans. Genetics 2015, 201, 1295–1306. [Google Scholar] [CrossRef] [Green Version]
- Buchman, A.B.; Ivy, T.; Marshall, J.M.; Akbari, O.S.; Hay, B.A. Engineered Reciprocal Chromosome Translocations Drive High Threshold, Reversible Population Replacement in Drosophila. ACS Synthet. Biol. 2018, 7, 1359–1370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beying, N.; Schmidt, C.; Pacher, M.; Houben, A.; Puchta, H. CRISPR–Cas9-mediated induction of heritable chromosomal translocations in Arabidopsis. Nat. Plants 2020, 6, 638–645. [Google Scholar] [CrossRef]
- Champer, J.; Buchman, A.; Akbari, O.S. Cheating evolution: Engineering gene drives to manipulate the fate of wild populations. Nat. Rev. Genet. 2016, 17, 146–159. [Google Scholar] [CrossRef] [Green Version]
- Reiter, L.T.; Potocki, L.; Chien, S.; Gribskov, M.; Bier, E. A systematic analysis of human disease-associated gene sequences in Drosophila melanogaster. Genome Res. 2001, 11, 1114–1125. [Google Scholar] [CrossRef] [Green Version]
- Bier, E. Drosophila, the golden bug, emerges as a tool for human genetics. Nat. Rev. Genet. 2005, 6, 9–23. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Name | Symbol | Chromosomal Location 1 | Mutation 2 | Amino Acid Substitution | Permissive Temperature | Restrictive Temperature | Phenotype (Restrictive Temperature) | References |
|---|---|---|---|---|---|---|---|---|
| shibire | shi | Chromosome X 15,892,116 to 15,906,716 [+] | shits1 | G268D | 22 °C | 27–29 °C | Embryonic lethal | Grigliatti, et al. [75] Poodry, et al. [78] |
| shits2 | G141S | 22 °C | 27–29 °C | Semi-embryonic lethal | ||||
| shits4 | P171S | 22 °C | 27–29 °C | Recessive, adult paralysis | Grigliatti, et al. [75] Kim, et al. [79] | |||
| shiSHY | T749I | 22 °C | 34.5 °C | Adult paralysis | Ramaswami, et al. [80] Krishnan, et al. [81] Narayanan, et al. [82] | |||
| Notch | N | Chromosome X 3,134,870 to 3,172,221 [+] | Nts1 | G1272D | 18 °C | 29 °C | Embryonic, larval (2nd and 3rd instars) and pupal lethality reported | Shellenbarger and Mohler [83] |
| Nts2 | - | 18 °C | 30 °C | Embryonic lethal | Shellenbarger and Mohler [83] Ge, et al. [84] | |||
| NAx-ts1 | - | 18 °C | 29 °C | Embryonic lethal | Shellenbarger and Mohler [83] | |||
| NAx28 | N986I | 25 °C | 29 °C | Lethal beginning at pupal stage | Portin and Sirén [85] | |||
| N60g11 (Dominant) | S2257G and 16 bp deletion removing T15442 to A15457 inclusive | Above 26 °C | Below 20 °C | Embryonic lethal, cold sensitive allele | Welshons and Von Halle [86] Foster [87] Fryxell and Miller [88] Lyman and Young [89] | |||
| RNA polymerase II 215kD subunit | RpII215 | Chromosome X 11,562,800 to 11,570,326 [−] | RpII215ts | R977C | 22 °C | 29 °C | Semi-embryonic lethal and completely lethal if continue treat at RT until first instar larvae | Mortin and Kaufman [90] |
| pale (tyrosine hydroxylase) | ple | Chromosome 3L 6,713,356 to 6,719,525 [−] | plets1 | C415R | 18 °C | 29 °C | Embryonic lethal | Pendleton, et al. [91] |
| transformer-2 | tra-2 | Chromosome 2R 14,602,004 to 14,604,337 [−] | tra-2ts1 | A151V | - | 16 °C, 29 °C | Sterility, pseudo XX males | Belote and Lucchesi [92] |
| tra-2ts2 | P181S | 16 °C | 29 °C | Sterility, pseudo XX males | ||||
| Downstream of raf1 | Dsor1 | Chromosome X 9,247,342 to 9,250,037 [+] | D-mekts1 | P209S | 20 °C | 25 °C | Lethal (unreported stage of development). Temperature sensitivity observed during oogenesis. | Hsu and Perrimon [93] |
| Saccharomyces cerevisiae Hanna, et al. [76] | Drosophila melanogaster Kuntamalla, et al. [77] | |||
|---|---|---|---|---|
| Temperature sensitive Allele | Amino Acid Substitution | Nucleotide Substitution | Temperature sensitive Allele | Amino Acid Substitution |
| ckα2-7 | A190T | GCG → ACG | YKP7 | A177T |
| T336M | ACG → ATG | |||
| ckα2-8 | E51K | GAA → AAA | ||
| G102D | GGC → GAC | YKP10 | G89D | |
| ckα2-11 | D225N | GAC → AAC | ||
| E299K | GAG → AAG | |||
| ckα2-12 | A190V | GCG → GTG | YKP8 | A177V |
| H294Y | CAC → TAC | |||
| ckα2-13 | D225N | GAC → AAC | YKP9 | D212N |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, T.N.M.; Choo, A.; Baxter, S.W. Lessons from Drosophila: Engineering Genetic Sexing Strains with Temperature-Sensitive Lethality for Sterile Insect Technique Applications. Insects 2021, 12, 243. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12030243
Nguyen TNM, Choo A, Baxter SW. Lessons from Drosophila: Engineering Genetic Sexing Strains with Temperature-Sensitive Lethality for Sterile Insect Technique Applications. Insects. 2021; 12(3):243. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12030243
Chicago/Turabian StyleNguyen, Thu N. M., Amanda Choo, and Simon W. Baxter. 2021. "Lessons from Drosophila: Engineering Genetic Sexing Strains with Temperature-Sensitive Lethality for Sterile Insect Technique Applications" Insects 12, no. 3: 243. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12030243