
Neelipleona and Symphypleona (Collembola) from a Sampling in the Mesovoid Shallow Substratum of the Sierra de Guadarrama National Park (Madrid and Segovia, Spain): Taxonomy and Biogeography



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
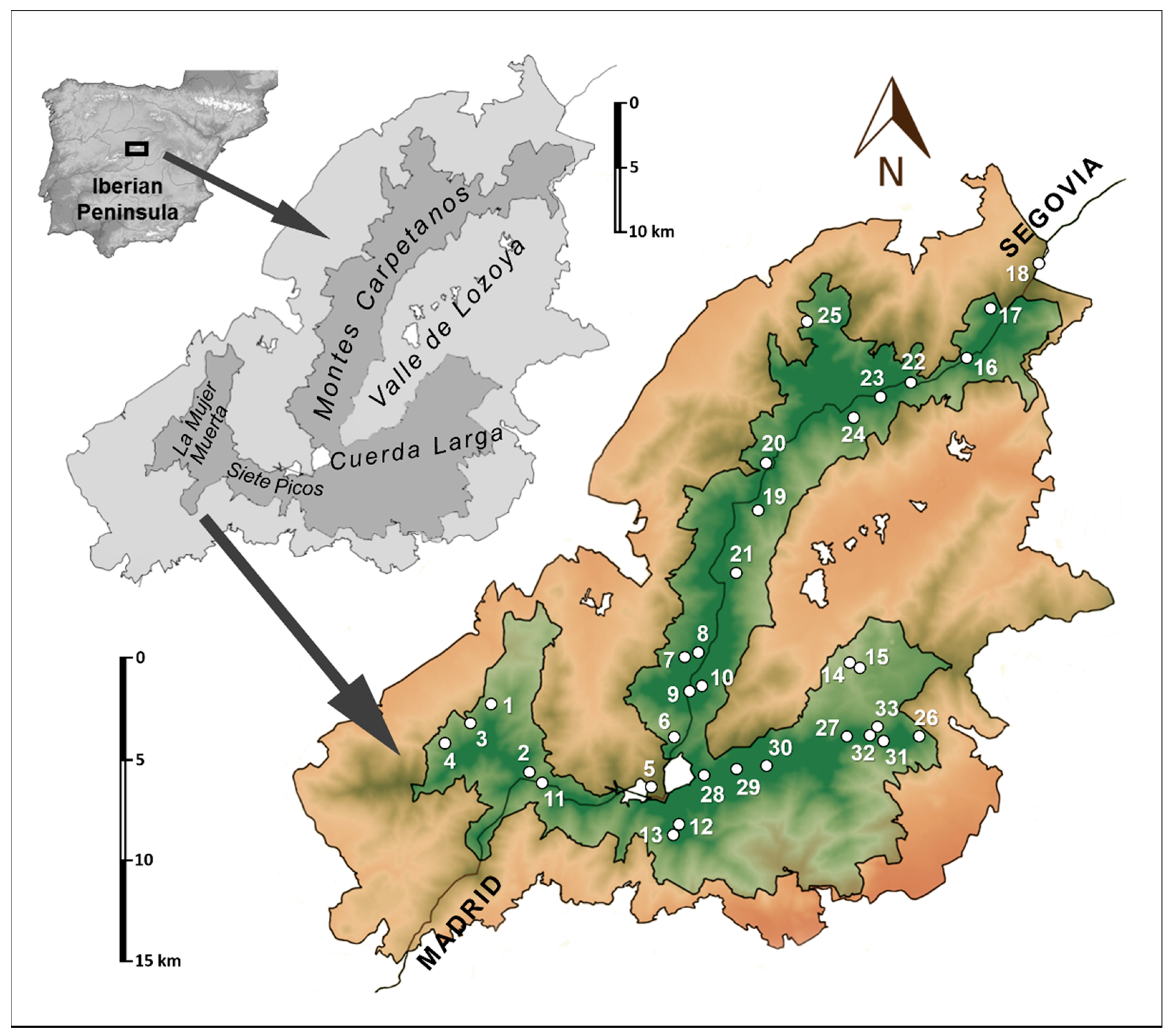
2.1. Site
2.2. Methodology
3. Results
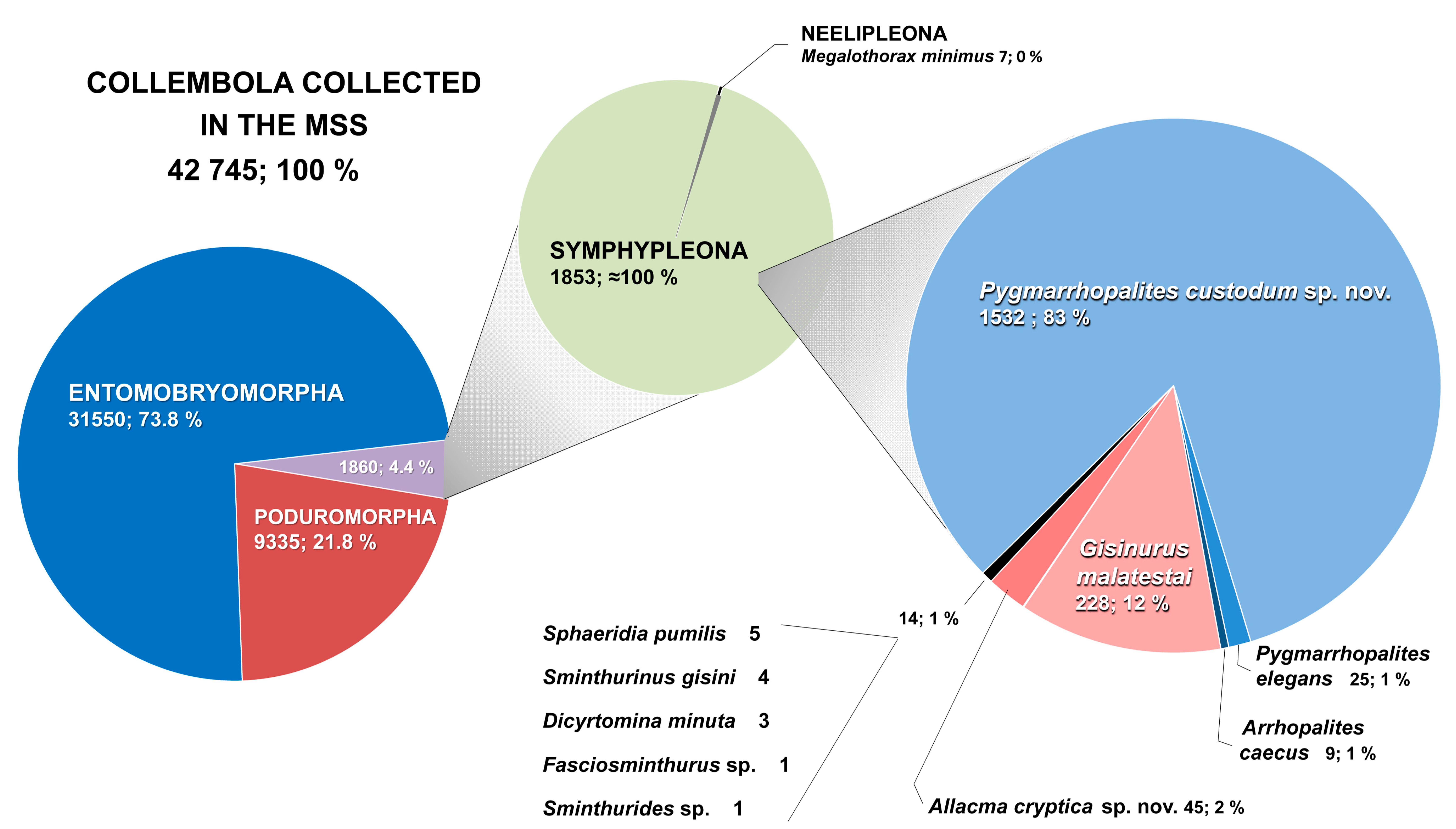
3.1. Summary
3.2. Taxonomy
3.2.1. Megalothorax minimus Willem, 1900
Material Studied
Remarks
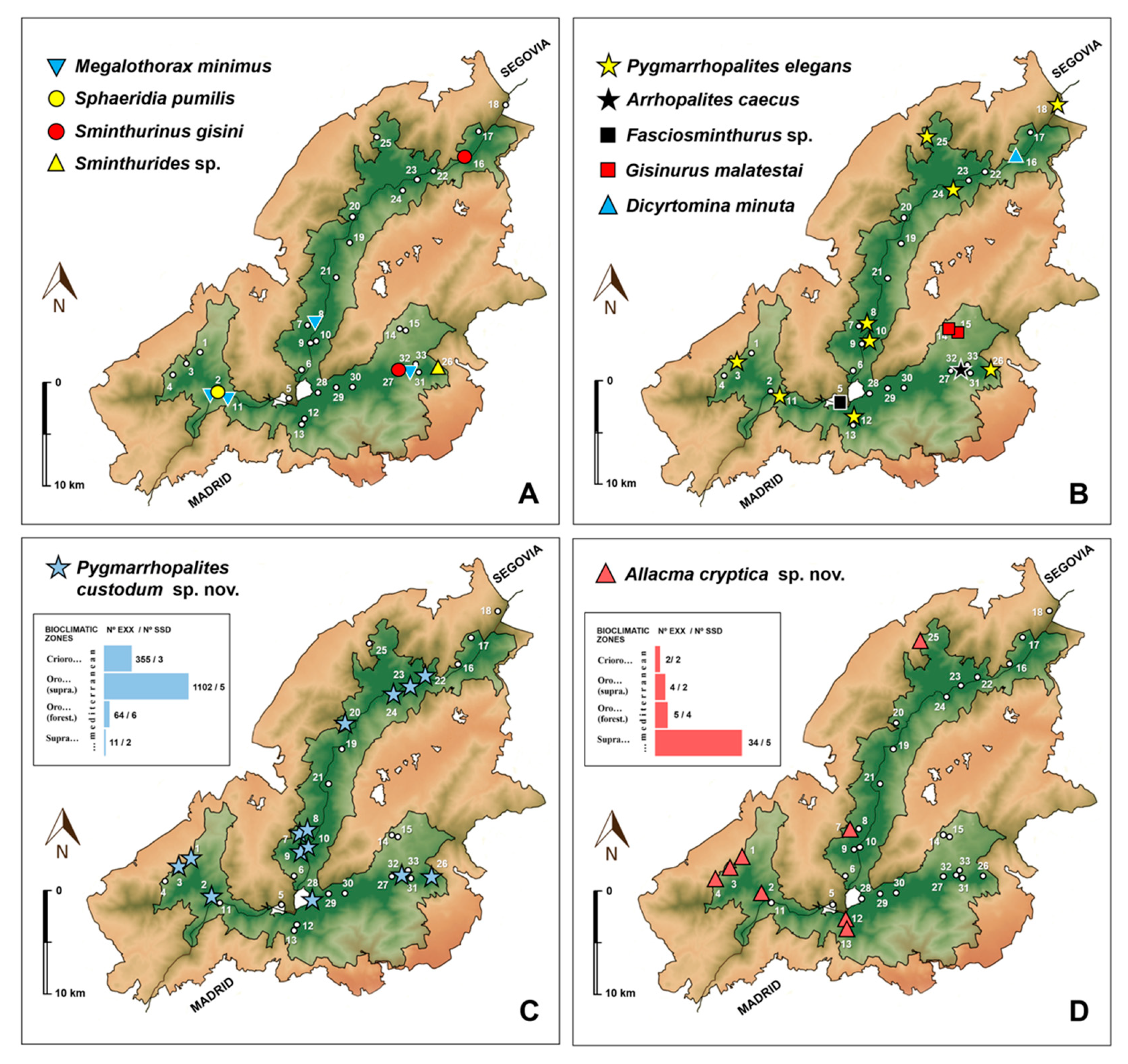
Ecology
3.2.2. Sphaeridia pumilis (Krausbauer, 1898)
Material Studied
Remarks
Ecology
3.2.3. Sminthurinus gisini Gama, 1965
Material Studied
Remarks
Ecology
3.2.4. Arrhopalites caecus (Tullberg, 1871)
Material Studied
Remarks
Ecology
3.2.5. Pygmarrhopalites elegans (Cassagnau and Delamare-Deboutteville, 1953)
Material Studied
Remarks
Ecology
3.2.6. Pygmarrhopalites custodum Baquero and Jordana sp. nov. (http://zoobank.org/1139329B-EE8F-45D2-A29D-F935DF913018, accessed on 20 March 2021)
Type Material
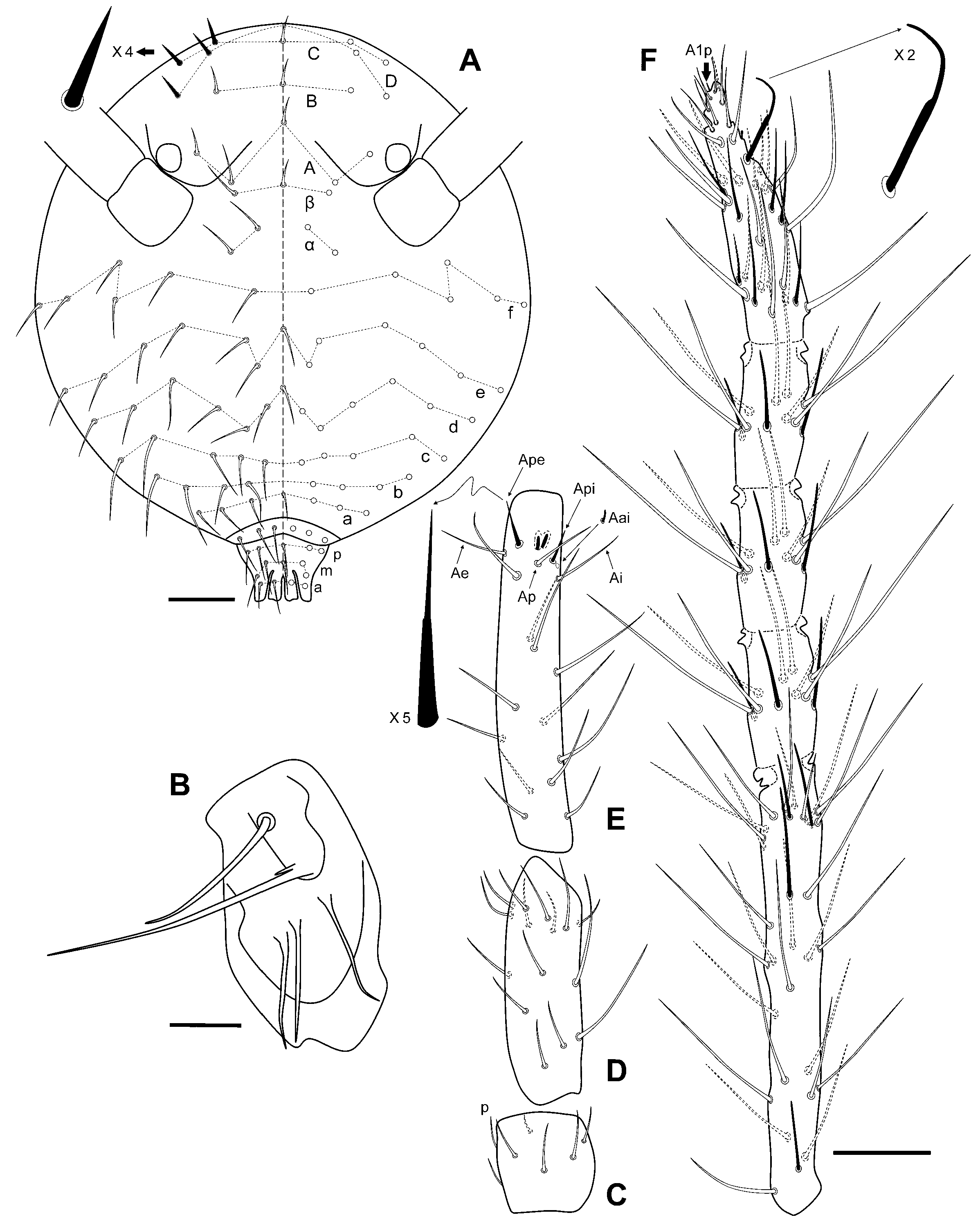
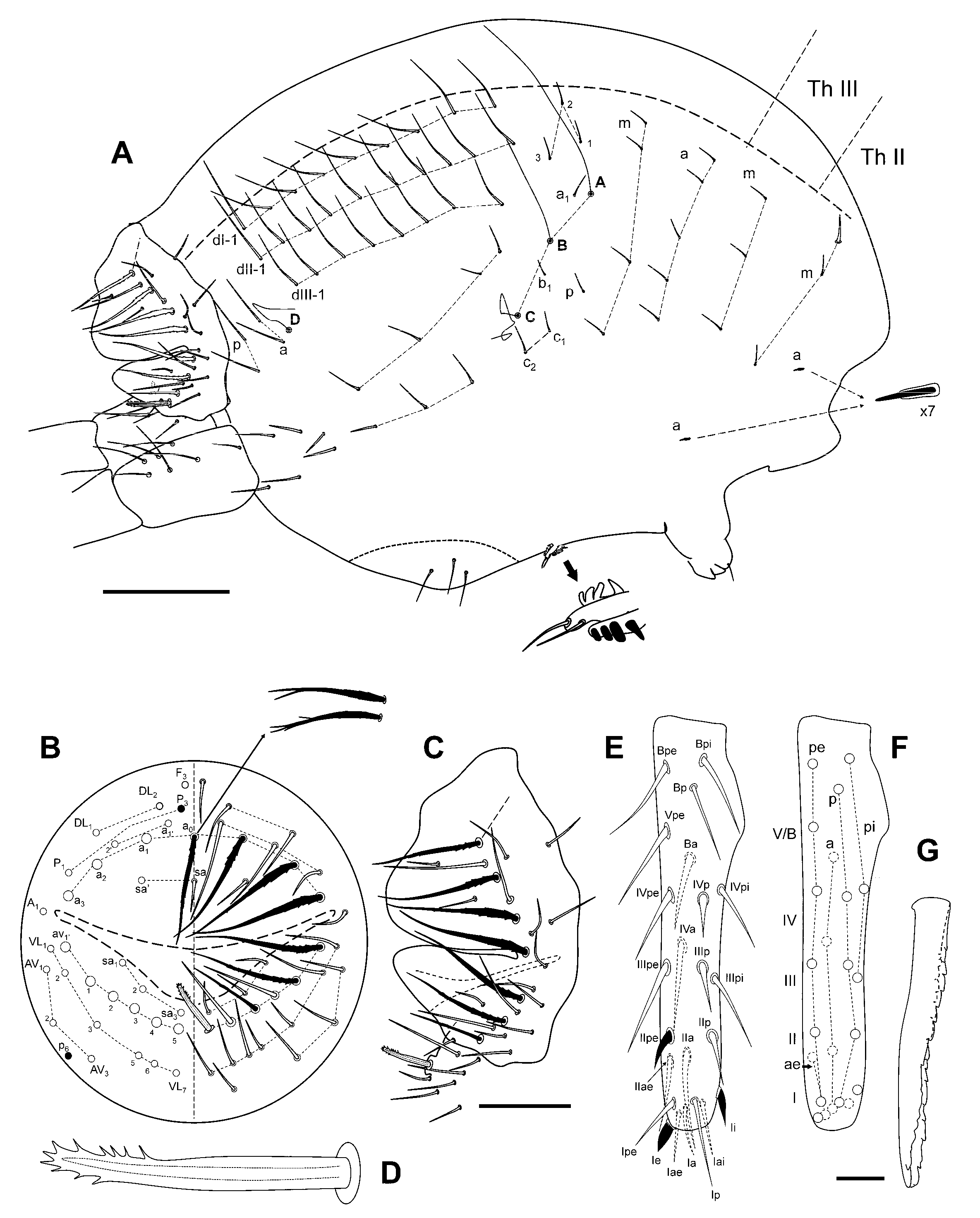
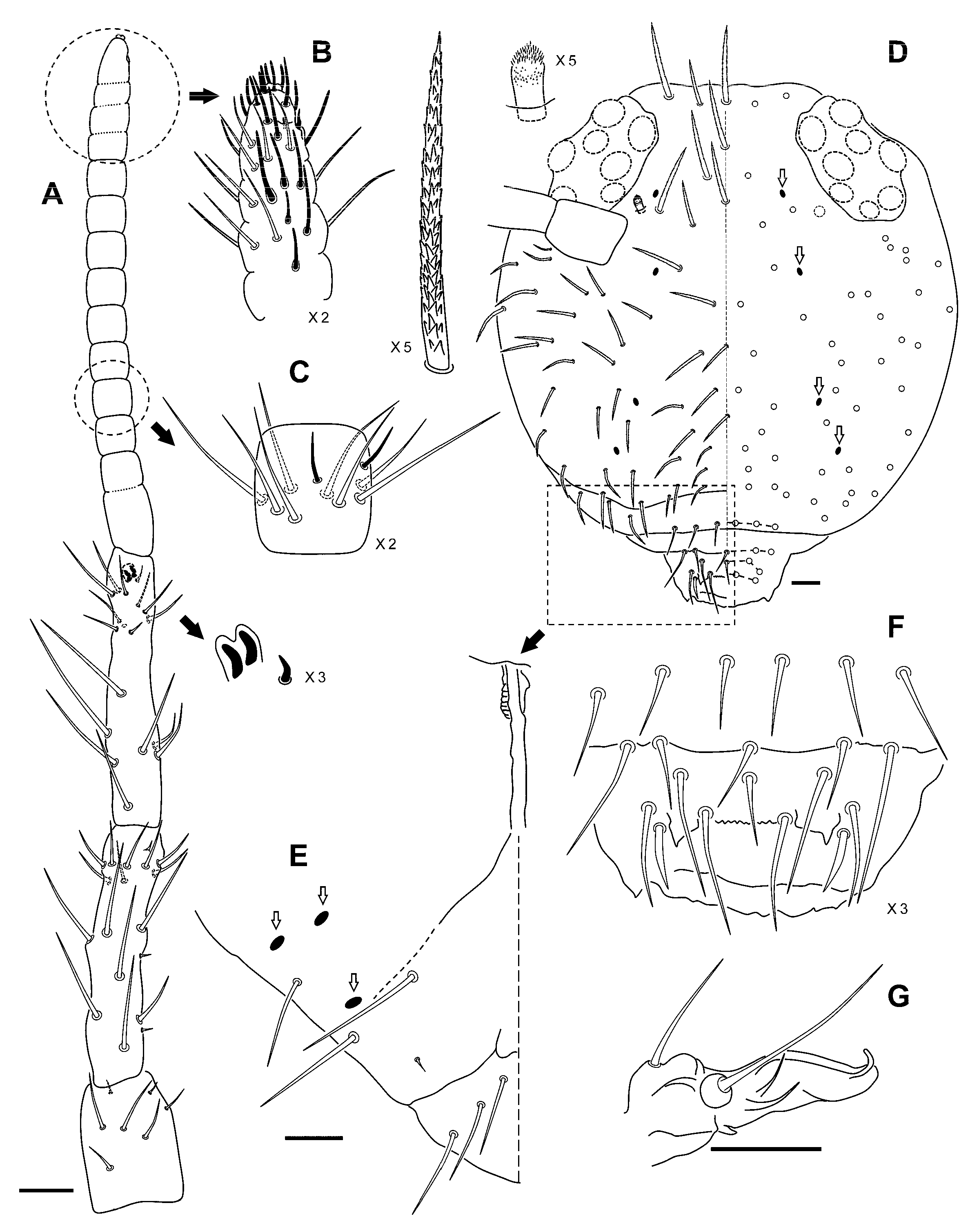
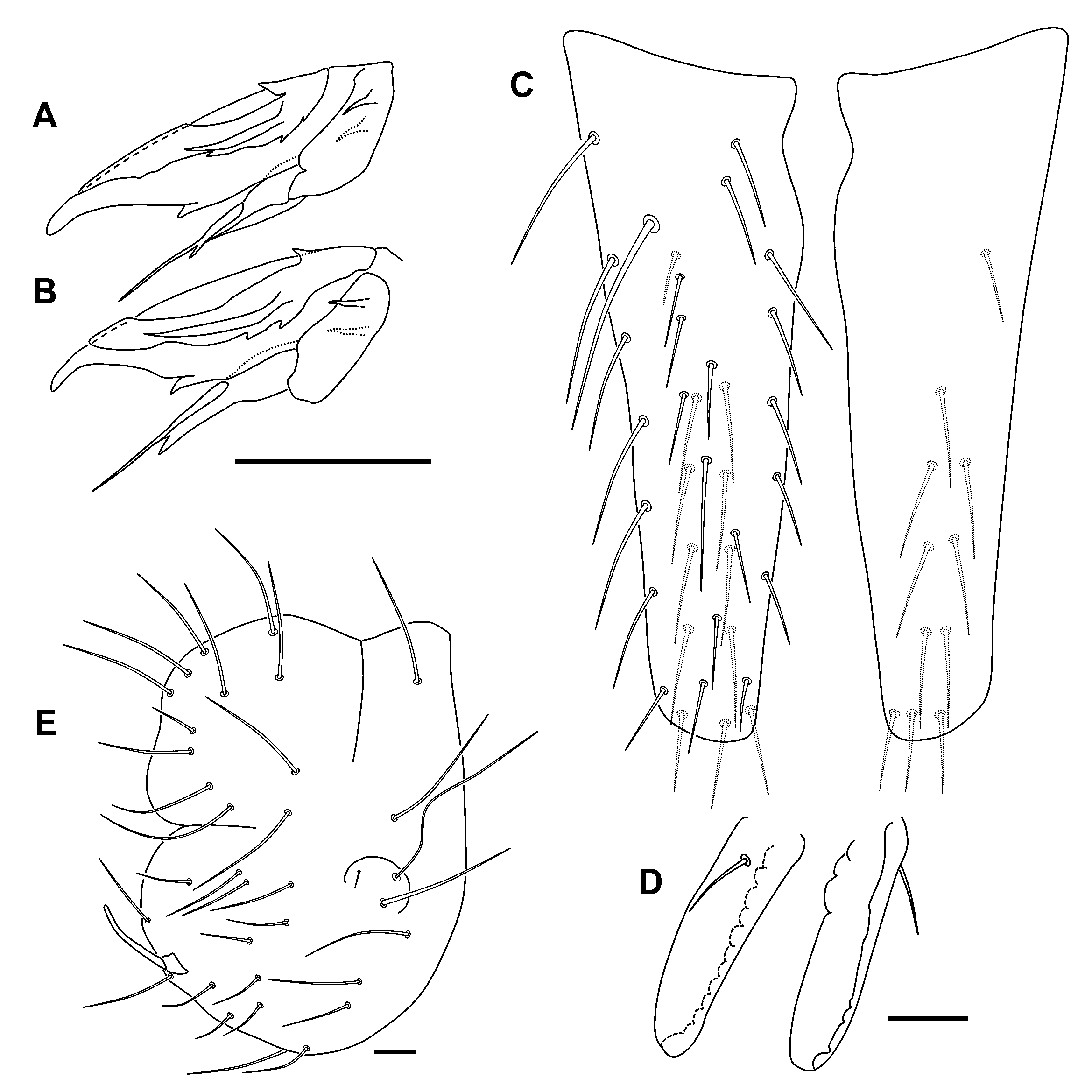
Diagnosis
Description
3.2.7. Allacma cryptica Baquero and Jordana sp. nov. (http://zoobank.org/51419F2B-F400-4AB8-B36B-FE7D1E111711, accessed on 20 March 2021)
Type Material
Diagnosis
Description
3.2.8. Gisinurus malatestai Dallai, 1970
Material Studied
Remarks
Ecology
3.2.9. Sminthurides sp.
Material Studied
Remarks
Ecology
3.2.10. Fasciosminthurus sp.
Material Studied
Remarks
Ecology
3.2.11. Dicyrtomina minuta (Fabricius, 1783)
Material Studied
Remarks
Ecology
4. General Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martínez, M.; Baquero, E.; Barranco, P.; Ariño, A.H.; Jordana, R. A new genus and species of Collembola from caves of south Iberian Peninsula (Collembola, Poduromorpha, Onychiuridae). Zootaxa 2004, 734, 1–15. [Google Scholar]
- Baquero, E.; Ledesma, E.; Gilgado, J.D.; Ortuño, V.M.; Jordana, R. Distinctive Collembola communities in the Mesovoid Shallow Substratum: First data for the Sierra de Guadarrama National Park (Central Spain) and a description of two new species of Orchesella (Entomobryidae). PLoS ONE 2017, 12, e0189205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordana, R.; Baquero, E.; Ledesma, E.; Ortuño, V.M. Poduromorpha (Collembola) from a sampling in the mesovoid shallow substratum of the Sierra de Guadarrama National Park (Madrid and Segovia, Spain): Taxonomy and Biogeography. Zool. Anz. 2020, 285, 81–96. [Google Scholar] [CrossRef]
- Baquero, E.; Jordana, R.; Ortuño, V.M. Distinctive Collembola Communities in the Mesovoid Shallow Substratum: Entomobryomorpha of the Sierra de Guadarrama National Park (Central Spain). Zoosystema 2021, 43, 37–78. [Google Scholar] [CrossRef]
- MAPAMA. Red de Parques Nacionales. Sierra de Guadarrama: Ficha Técnica; Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 2017. Available online: http://www.mapama.gob.es/es/red-parques-nacionales/nuestros-parques/guadarrama/ficha-tecnica/default.aspx (accessed on 30 April 2017).
- Vialette, Y.; Casquet, C.; Fúster, J.M.; Ibarrola, E.; Navidad, M.; Peinado, M.; Villaseca, C. Geochronological study of orthogneisses from the sierra de Guadarrama (Spanish central system). N. Jb. Miner. Mh. 1987, 10, 465–479. [Google Scholar]
- Sanz, C. Periglaciarismo en montaña: La Sierra de Guadarrama. In Atlas de Geomorfología; Martínez de Pisón, E., Ed.; Alianza Editorial: Madrid, Spain, 1986; pp. 239–254. [Google Scholar]
- Pedraza de, J.; Carrasco, R.M. El glaciarismo Pleistoceno del Sistema Central. Enseñanza Ciencias Tierra 2005, 13, 278–288. Available online: http://www.raco.cat/index.php/ECT/issue/archive (accessed on 15 January 2021).
- Rivas–Martínez, S. Pisos bioclimáticos de España. Lazaroa 1984, 5, 33–43. [Google Scholar]
- Rivas–Martínez, S.; Belmonte, D.; Cantó, P.; Fernández–González, F.; de la Fuente, V.; Moreno, J.M.; Sánchez–Mata, D.; Sancho, L.G.; Leopoldo, G. Piornales, enebrales y pinares oromediterráneos (Pino-Cytision oromediterranei) en el Sistema Central. Lazaroa 1987, 7, 93–124. [Google Scholar]
- Ortuño, V.M.; Ledesma, E.; Jiménez–Valverde, A.; Pérez‑Suárez, G. Studies of the mesovoid shallow substratum can change the accepted autecology of species: The case of ground beetles (Coleoptera, Carabidae) in the Sierra de Guadarrama National Park (Spain). Anim. Biodiv. Conserv. 2019, 42, 213–226. [Google Scholar] [CrossRef]
- Vargovitsh, R.S. New cave Arrhopalitidae (Collembola: Symphypleona) from the Crimea (Ukraine). Zootaxa 2009, 2047, 1–47. [Google Scholar] [CrossRef]
- Fjellberg, A. The maxillary outer lobe, an important systematic tool in Isotomidae (Collembola). Bull. Ann. Soc. R. Belge Entomolog. 1984, 114, 83–88. [Google Scholar]
- Nayrolles, P. La chetotaxie antennaire des Collemboles Symphypleones. Trav. Lab. Ecobiol. Arth. Edaph. 1991, 6, 1–94. [Google Scholar]
- Bretfeld, G. Synopses on Palaearctic Collembola, Volume 2. Symphypleona. Abh. Ber. Naturkundemus. Görlitz 1999, 71, 1–318. [Google Scholar]
- Christiansen, K. The genus Arrhopalites (Collembola: Sminthuridae) in the United States and Canada. Int. J. Speleol. 1966, 2, 43–73. [Google Scholar] [CrossRef] [Green Version]
- Christiansen, K.; Bellinger, P.F. The Collembola of North America, North of the Rio Grande, Part 4, Families Neelidae and Sminthuridae, 2nd ed.; Publications of the Grinnell College: Grinnell, IA, USA, 1998; pp. 1175–1520. [Google Scholar]
- Börner, C. Das System der Collembolen nebst Beschreibung neuer Collembolen des Hamburger Naturhistorischen Museums. Mit. Natur. Mus. Hamburg 1906, 23, 147–188. [Google Scholar]
- Dallai, R. Ricerche sui Collemboli. XIV. Le Alpi Apuane. Lav. Soc. Ital. Biogeogr. 1970, 1, 433–482. [Google Scholar] [CrossRef] [Green Version]
- Lubbock, J. Notes on the Thysanura, Part III. Trans. Linn. Soc. Lond. 1870, 26, 295–304. [Google Scholar] [CrossRef]
- Massoud, Z. Contribution à la connaissance morphologique et systématique des Collemboles Neelidae. Rev. Ecol. Biol. Sol. 1971, 8, 195–198. [Google Scholar]
- Folsom, J.W. Neelus Murinus, representing a new thysanuran family. Psyche 1896, 7, 391–392. [Google Scholar] [CrossRef] [Green Version]
- Willem, V. Un type nouveau de Sminthuride: Megalothorax. Ann. Soc. Ent. Belg. 1900, 44, 7–10. [Google Scholar]
- Selga, D. Catálogo de los colémbolos de la Península Ibérica. Graellsia 1971, 24, 133–283. [Google Scholar]
- Börner, C. Collembola of Khumbu Himal. Ergeb. Forsch. Nepal 1901, 4, 80–130. [Google Scholar]
- Betsch, J.M.; Massoud, Z. Etudes sur les Insectes Collemboles. I.-Systematique, ultrastructure externe et ecologie du genre Jeannenotia Stach,1956 (Symphypleones, Sminthurididae n. comb.). Description de deux Collemboles nouveaux (Proisotoma et Sminthurides). Rev. Ecol. Biol. Sol 1970, 7, 153–225. [Google Scholar]
- Krausbauer, T. Neue Collembola aus der Umgebung von Weilburg an der Lahn. Zool. Anz. 1898, 21, 495–499, 501–504. [Google Scholar]
- Gama, M.M. Contribuçao para o estudio dos Colêmbolos do Arquipélago da Madeira. Mem. Estud. Mus. Zool. Univ. Coimbra 1959, 257, 1–42. [Google Scholar]
- Selga, D. Proisotoma (Collembola) de los suelos de Guadarrama con descripción de una nueva especie del mismo género. Bol. Soc. Esp. Hist. Nat. 1962, 60, 69–76. [Google Scholar]
- Börner, C. Die Familien der Collembolen. Zool. Anz. 1913, 41, 315–322. [Google Scholar]
- Gama, M.M. Sminthurinus gisini n. sp. provenant du Portugal (Insecta, Collembola). Rev. Ecol. Biol. Sol 1965, 2, 277–279. [Google Scholar]
- Rusek, J. Eine neue Sminthurinus-Art (Collembola) aus der Mittel-Slowakei. Acta Entomol. Bohemoslov. 1966, 63, 25–26. [Google Scholar]
- Dunger, W. Beitrag zur Collembolenfauna des Bruby Jesenik-Gebriges (Altvatergebirge). Acta Mus. Sil. 1970, 19, 35–44. [Google Scholar]
- Weiner, W.M. Collembola of the Pieniny National Park in Poland. Acta Zool. Cracov. 1981, 25, 417–500. [Google Scholar]
- Bauer, R. Zur Zönologie und Phänologie granitblockbesiedelnder Collembolen. Zool. Anz. 1993, 5/6, 237–248. [Google Scholar]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of This Group of Insects. Family: Sminthuridae; Polska Akademia Nauk: Krakowie, Poland, 1956; pp. 1–287. [Google Scholar]
- Tullberg, T.F. Forteckning öfver Svenska Podurider. Öfversigt af. Kongl. Vetenskaps-Akademiens. Förhandlingar 1871, 28, 143–155. [Google Scholar]
- Cassagnau, P.; Delamare-Deboutteville, C. Les Arrhopalites et Pararrhopalites d’Europe. (Collemboles Symphypleones Cavernicoles). Notes Biospeol. 1953, 8, 133–147. [Google Scholar]
- Yoshii, R. Höhlencollembolen Japans I. Kontyu 1954, 20, 62–70. [Google Scholar]
- Vargovitsh, R.S. New troglomorphic Arrhopalitidae (Collembola: Symphypleona) from the Western Caucasus. Zootaxa 2012, 3174, 1–21. [Google Scholar] [CrossRef]
- Vargovitsh, R.S. Arrhopalites potapovi sp. nov. (Collembola, Symphypleona) from Russia. Zootaxa 2015, 3955, 101–112. [Google Scholar] [CrossRef] [Green Version]
- Jordana, R.; Fadrique, F.; Baquero, E. The collembolan fauna of Maestrazgo caves (Teruel, Spain) with description of three new species. Zootaxa 2012, 3502, 49–71. [Google Scholar] [CrossRef] [Green Version]
- Jordana, R.; Barranco, P.; Amezcua, A.; Baquero, E. Two new species of Collembola (Hexapoda) from Saliente Cave (Almería, Spain)—Deux nouvelles espèces de collemboles (Hexapoda) de grotte du Saliente (Almería, Espagne). Zoosystema 2017, 39, 103–115. [Google Scholar] [CrossRef]
- Vargovitsh, R.S. Two new troglobiont Pygmarrhopalites species of the principalis group (Collembola: Arrhopalitidae) from the West Caucasus. Zootaxa 2017, 4250, 23–42. [Google Scholar] [CrossRef]
- Loksa, I.; Rubio, I. Angaben zu den Kenntnissen über die Collembolenfauna des Bakony-Gebirges. Opusc. Zool. 1966, 6, 139–156. [Google Scholar]
- Vargovitch, R.S. Arrhopalites kristiani sp. n. (Collembola, Arrhopalitidae) from a cave in Eastern Carpathians. Vestn. Zool. 2005, 39, 67–73. [Google Scholar]
- Park, K.H.; Kang, J.S. Three new species of Arrhopalites (Collembola, Symphypleona, Arrhopalitidae) from Korea. Entomol. Res. 2007, 37, 157–162. [Google Scholar] [CrossRef]
- Arbea, J. Los Colémbolos (Hexapoda, Collembola) subterráneos de Jaén. In Los Invertebrados de Hábitats Subterráneos de Jaén; Pérez-Fernández, T., Pérez Ruiz, A., Eds.; Grupo de Espeleología de Villacarrillo (G.E.V.): Jaén, Spain, 2013; pp. 104–186. [Google Scholar]
- Linnaniemi, W.M. Die Apterygotenfauna Finlands II. Specieller Teil. Acta Soc. Sci. Fenn. 1912, 40, 1–361. [Google Scholar]
- Soto-Adames, F.N.; Taylor, S.J. New species and new records of springtails (Hexapoda: Collembola) from caves in the Salem Plateau of Illinois, USA. J. Cave Karst Stud. 2013, 75, 146–175. [Google Scholar] [CrossRef]
- Curcic, B.P.M.; Lucic, L.R. Arrhopalites zloti, a new species of cave springtails (Sminthuridae, Collembola) from Serbia, Yugoslavia. Arch. Biol. Sci. 1997, 49, 31–32. [Google Scholar]
- Lubbock, J. Notes on the Thysanura, Part I: Smynthuridae. Trans. Linn. Soc. Lond. 1862, 23, 429–448. [Google Scholar] [CrossRef]
- Deharveng, L. Recent advances in Collembola systematics. In Proceedings of the 6th International Seminar on Apterygota, Siena, Italy, 13–17 September 2002; pp. 415–433. [Google Scholar]
- Linnæus, C. Systema Naturæ per Regna tria Naturæ, Secundum Classes, Ordines, Genera, Species, cum Characteribus, Differentis, Synonymis, Locis; Tomus I, Editio Decima, Reformata, Holmiæ, (Laurentii Salvii); Lugduni: Apud J. B. Delamolliere: Lyon, France, 1758; pp. 1–824. [Google Scholar]
- Carl, J.H. Über die Collembola der Schweiz. Rev. Suisse Zool. 1899, 6, 264–362. [Google Scholar]
- Yoshii, R.; Lee, C. On some Collembola of Korea with notes on the genus Ptenothrix. Contrib. Biol. Lab. Kyoto Univ. 1963, 15, 1–37. [Google Scholar]
- Betsch, J.M. Eléments pour une monographie des Collemboles Symphypléones (Hexapodes, Aptérigotes). Mém. Mus. natl. Hist. Nat. 1980, 116, 1–227. [Google Scholar]
- Nayrolles, P. A standardized description of European Sminthuridae (Collembola, Symphypleona), 1: Genera Lipothrix, Gisinurus, and Caparainea. Bijdr. Dierkd. 1993, 63, 43–60. [Google Scholar] [CrossRef] [Green Version]
- Gama, M.M. Colémbolos das Canárias (Insectos, Apterigotas). In Proceedings of the Actas III Congreso Ibérico de Entomología, Granada, Spain, October 1987; pp. 73–89. [Google Scholar]
- Bretfeld, G. Sturmius epiphytus n. gen. n. spec. from Colombia, a taxon of the Symphypleona (Insecta, Collembola) with an unexpected character combination. Z. Zool. Syst. Evol. 1994, 32, 264–281. [Google Scholar] [CrossRef]
- Fabricius, O. Beskrivelse over nogle lidet bekiendte podurer, og en besonderlig Loppe. Nye Saml. K. Dansk.Vid. Selsk. Skr. 1783, 2, 296–311. [Google Scholar]
- Arbea, J.I.; Jordana, R. Estudio ecológico de la colembofauna de los suelos del Macizo de Quinto Real (Pirineos Occidentales) y descripción de dos especies nuevas: Anurida flagellata sp. n. y Onychiurus subedinensis sp. n. (Insecta, Collembola). Bol. Est. Cent. Ecología 1985, 28, 57–80. [Google Scholar]
- Jordana, R.; Arbea, J.I.; Ariño, A.H. Catálogo de colémbolos ibéricos. Base de datos. Pub. Biol. Univ. Navarra 1990, 21, 1–231. [Google Scholar]
- Gama, M.M.; Lopes, C.M.; Nogueira, A. Etude comparée de populations de Collemboles de plusieurs biotopes de l’Algarve (Portugal). In Proceedings of the 3rd International Seminar on Apterygota, Siena, Italy, 21–26 August 1989; pp. 363–370. [Google Scholar]
- Massoud, Z.; Thibaud, J.M. Essai de classification des Collemboles «cavernicoles» européens. In Proceedings of the 6 Congrès International de Spéléologie, Olomouc, Czechoslovakia, 31 August–18 September 1973; Volume 5, pp. 141–157. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mountain Areas of the Sierra de Guadarrama | Code | UTM Coordinates (100 × 100 m) | Altitude (m a. s. l.) | Toponymy/Province | Date Installation of Traps | Date of Trap Recovery | Orientation |
|---|---|---|---|---|---|---|---|
| Siete Picos—La Mujer Muerta | SSD-1 | 30 T 4081 45204 | 1606 | Cancho del Río Peces/Segovia | 20 May 2015 | 17 September 2015 | North |
| SSD-2 | 30 T 4100 45166 | 1818 | Corrales de la Majada Minguete/Segovia | 20 May 2015 | 17 September 2015 | Northeast | |
| SSD-3 | 30 T 4068 45192 | 1622 | Umbría de la Mujer Muerta/Segovia | 21 May 2015 | 17 September 2015 | North | |
| SSD-4 | 30 T 4056 45181 | 1685 | Majada Conejo/Segovia | 21 May 2015 | 17 September 2015 | Northwest | |
| SSD-11 | 30 T 4108 45161 | 1876 | Cerro Ventoso/Madrid | 9 June 2015 | 17 September 2015 | East | |
| Puerto de los Cotos—Puerto de Navacerrada | SSD-5 | 30 T 4166 45159 | 1923 | Arroyo Seco/Segovia | 27 May 2015 | 22 September 2015 | Northwest |
| Montes Carpetanos | SSD-7 | 30 T 4185 45229 | 1994 | Majada Hambrienta/Segovia | 2 June 2015 | 17 September 2015 | Northwest |
| SSD-8 | 30 T 4190 45231 | 2071 | Majada Aranguez/Segovia | 2 June 2015 | 17 September 2015 | Northwest | |
| SSD-9 | 30 T 4187 45211 | 2208 | Dos Hermanas/Madrid | 3 June 2015 | 5 October 2015 | East | |
| SSD-10 | 30 T 4191 45213 | 2049 | Hoya de la Laguna Grande/Madrid | 3 June 2015 | 5 October 2015 | East | |
| SSD-16 | 30 T 4334 45389 | 1956 | Las Revueltas—Los Horcos/Segovia | 23 June 2015 | 7 October 2015 | West | |
| SSD-18 | 30 T 4373 45438 | 1885 | Los Loberos/Segovia | 23 June 2015 | 7 October 2015 | Northwest | |
| SSD-20 | 30 T 4226 45332 | 1937 | Cerro de Navahonda/Segovia | 24 June 2015 | 6 October 2015 | Northeast | |
| SSD-22 | 30 T 4304 45376 | 1995 | Alto del Puerto/Segovia | 24 June 2015 | 22 September 2015 | North | |
| SSD-23 | 30 T 4288 45367 | 2144 | Circo del Pico Nevero/Madrid | 25 June 2015 | 6 October 2015 | Southeast | |
| SSD-24 | 30 T 4274 45357 | 2042 | Peñacabra/Madrid | 25 June 2015 | 22 October 2015 | East | |
| SSD-25 | 30 T 4249 45407 | 1731 | Arroyo del Charco (La Cepa)/Segovia | 2 July 2015 | 22 October 2015 | Northwest | |
| Cuerda Larga and Associated Mountainous complex | SSD-12 | 30 T 4180 45138 | 2102 | Collado del Piornal/Madrid | 9 June 2015 | 22 September 2015 | North |
| SSD-13 | 30 T 4179 45135 | 2113 | Los Almorchones—Las Buitreras/Madrid | 10 June 2015 | 22 September 2015 | Southwest | |
| SSD-14 | 30 T 4274 45224 | 1406 | El Purgatorio/Madrid | 18 June 2015 | 5 October 2015 | West | |
| SSD-15 | 30 T 4273 45224 | 1375 | Hueco de los Ángeles/Madrid | 18 June 2015 | 5 October 2015 | Northeast | |
| SSD-26 | 30 T 4309 45186 | 1890 | La Najarra—Cuatro Calles/Madrid | 2 July 2015 | 30 October 2015 | East | |
| SSD-27 | 30 T 4270 45185 | 2101 | Bailaderos/Madrid | 2 July 2015 | 30 October 2015 | North | |
| SSD-28 | 30 T 4193 45164 | 2156 | Collado de Valdemartín/Madrid | 3 July 2015 | 6 November 2015 | North | |
| SSD-32 | 30 T 4285 45187 | 1948 | Arroyo de La Najarra/Madrid | 9 July 2015 | 22 October 2015 | Northeast |
| Species/Character | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | D |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A. antrobius | 1 | 8 | U | 5 | 1 | 1 | 2 | 1 | 1 | 2 | 1 | 1 | 1 | 0 * | 2 | 0 | 5 * | 1 * | 1 * | 1 | 1 * | 1 * | 3 * | 1 * | 1 * | 1 * | 6 * | 0 * | 2 * | 2 * | 2 * | 2 * | 2 * | 2 * | 18 |
| A. macronyx | 1 | 0 * | 0 | 0 * | 0 * | 0 * | 0 * | 0 * | 0 * | 0 * | 0 * | 1 | 0 * | 0 * | 1–2 | 0 | 0 * | 0 * | 0 | 0 * | 0 | 0 | 0 * | 0 * | 0 | 0 | 9 * | 0 | 3 | 3 | 3 | 3 | 3 | 1 * | 18 |
| A. potapovi | 1 | 13 * | 1 * | 0 * | 1 | 1 | 2 | 1 | 1 | 2 | 1 | 0–1 | 1 | 0 * | 1 | 1 * | 5 * | 1 * | 1 * | 1 | 1 * | 1 * | 3 * | 1 * | 1 * | 1 * | 5 | 0 | 3 | 2 * | 3 | 3 | 3 | 3 * | 16 |
| P. cantavetulae | 1 | 0 * | 0 | 5 | 1 | 1 | 2 | 1 | 1 | 2 | 1 | 1 | 1 | 1 | 2 | 0 | 2 | 2 | 0 | 1 | 0 | 0 | 3 * | 2 | 1 * | 1 * | 5 | 0 | 3 | 3 | 3 | 2 * | 2 * | 0 | 6 |
| P. crepidinis | 1 | 0 * | 0 | 5 | 1 | 1 | 2 | 1 | 1 | 2 | 1 | 1 | 1 | 1 | 2 | 0 | 2 | 2 | 0 | 0–1 | 0 | 0 | 0(1) | 0(1) * | 0 | 0 | 5 | 0 | 2 * | 2 * | 2(4) * | 2 * | 1 * | 1 * | 8 |
| P. dbari | 1 | 11 * | U | 7 * | 1 | 1 | 1 * | 1 | 1 | 0 * | 1 | 0 | 0 * | 1 | 2 | 0 | 2 | 2 | 0 | 2 * | 0 | 0 | 1 | 1 * | 0 | 0 | 8 * | 0 | 0 * | 1 * | 1 * | 1 * | 3 * | 0 | 13 |
| P. dudichi | 1 | 8 | 0 | 6 * | 0 * | 1 | 2 | 0 * | 1 | 2 | 0 * | 1 | 1 | 1 | U | 0 | 2 | 2 | 0 | 1 | 0 | 0 | U | U | U | U | 0 * | U | U | U | U | U | U | U | 5 |
| P. kovali | 1 | 8 | 0 | 5 | 1 | 1 | 1 * | 1 | 1 | 1 * | 1 | 1 | 1 | 1 | 2 | 0 | 2 | 2 | 0 | 2 * | 0 | 0 | 3 * | 1 * | 1 * | 1 * | 8 * | 0 | 1 * | 1 * | 1 * | 1 * | 1 * | 0 | 13 |
| P. kristiani | 1 | 0 * | 0 | 5 | 0 * | 0 * | 2 | 0 * | 0 * | 1 * | 0 * | 1 | 1 | 1–2 | 1 | 0 | 1 * | 2 | 0 | 0 * | 0 | 0 | 0 * | 0 * | 0 | 0 | U | 0 | 3 | 3 | 2 * | 2 * | 1 * | 0 | 14 |
| P. nigripes | 2? | 9 * | 0 | 7 * | 1 | 1 | 2 | 1 | 1 | 2 | 1 | 1 | 1 | 1 | 2 | 0 | 4 * | 2 | 0 | 1 | 1 * | 1 * | 3 * | 1 * | 1 * | 1 * | 2 * | 0 | 3 | 3 | 3 | 3 | 3 | 3 * | 11 |
| P. perezi | 1 | 8 | 0 | 5 | 1 | 1 | 2 | 1 | 1 | 2 | 1 | 1 | 1 | 1 | 2 | 0 | 3 * | 2 | 1 * | 1 | 0 | 0 | 3 * | 1 * | 1 * | 1 * | 1 * | 0 | 3 | 3 | 3 | 2 * | 2 * | 2 * | 10 |
| P. principalis pallida | 1 | 8 | 0 | 6 * | 0 * | 1 | 2 | 1 | 1 | 2 | 1 | 1 | 1 | 1 | 2 | U | 3 * | 2 | 1 * | 1 | 0 | 0 | 3 * | 1 * | 1 * | 1 * | 2 * | 1 * | 1 * | 1 * | 4 * | 4 * | 1 * | 1 * | 16 |
| P. pseudoprincipalis | 1 | 8 | U | 5 | 1 | 1 | 2 | 1 | 1 | 2 | 1 | 0 | 1 | 1 | 2 | U | 4 * | 2 | 0 | 2 * | 1 * | 1 * | 3 * | 1 * | 1 * | 1 * | 2 * | 0 | 1 * | 1 * | 1 * | 1 * | 1 * | 1 * | 15 |
| P. salemensis | 1 | 0 * | 0 | 5 | 1 | 1 | 2 | 1 | 1 | 2 | 1 | 0–1 | 1 | U | 2 | 0 | 2 | 2 | 0 | 2 * | 0 | 0 | 3 * | 2 | 1 * | 1 * | 2 * | 0 | 3 | 3 | 3 | 3 | 3 | 1 * | 7 |
| P. zloti | 1 | 13 * | 0 | 5 | 0 * | U | 2 | 1 | U | 0 * | 1 | U | 0 * | 1 | U | 0 | 2 | 2 | 0 | 1 | 0 | 0 | 1 | 1 * | 0 | 0 | U | U | U | U | U | U | U | U | 5 |
| P. custodum sp. nov. | 1 | 8 | 0 | 5 | 1 | 1 | 2 | 1 | 1 | 2 | 1 | 0–1 | 1 | 1 | 1–2 | 0 | 2 | 2 | 0 | 1 | 0 | 0 | 1 | 2 | 0 | 0 | 5 | 0 or 2 | 3 | 3 | 3 | 3 | 3 | 0 |
| Specie/Character | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | D |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A. fusca | 8 | 1 * | 3 | 5 | 5 | 15 | 1 * | 1 | 0 * | 1 | 1 * | 1 * | 1 * | 1 * | 2 | 6–8 | 2 * | 2 | 2 | 2 | 1 | 0 | 1 | 1 * | 3 * | 10 |
| A. gallica | 8 | 2 | 3 | 3 * | 5 | 13 | 2 | 0 * | 0 * | U | 0–1 | 0–1 | 1 * | 0 | U | 6–9 | 2 * | 2 | 2 | 2 | 2 * | 1 * | 1 | 0 | 3 * | 8 |
| A. koreana | 8 | 3 * | U | 4 | 4 | 12 * | 1 * | 1 | 0–1 | 1 | 1 * | 1 * | 1 * | 0 | 3 | 7 * | 2 * | 2 | 2 | 2 | 1 | 0 | 1 | 0 | 4 * | 9 |
| A. cryptica sp. nov. | 8 | 2 | 2–3 | 4–5 | 4–5 | 13–15 | 2 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 1–3 | 6 | 3 | 2 | 2 | 2 | 1 | 0 | 1 | 0 | 1–2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baquero, E.; Jordana, R.; Ortuño, V.M. Neelipleona and Symphypleona (Collembola) from a Sampling in the Mesovoid Shallow Substratum of the Sierra de Guadarrama National Park (Madrid and Segovia, Spain): Taxonomy and Biogeography. Insects 2021, 12, 266. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12030266
Baquero E, Jordana R, Ortuño VM. Neelipleona and Symphypleona (Collembola) from a Sampling in the Mesovoid Shallow Substratum of the Sierra de Guadarrama National Park (Madrid and Segovia, Spain): Taxonomy and Biogeography. Insects. 2021; 12(3):266. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12030266
Chicago/Turabian StyleBaquero, Enrique, Rafael Jordana, and Vicente M. Ortuño. 2021. "Neelipleona and Symphypleona (Collembola) from a Sampling in the Mesovoid Shallow Substratum of the Sierra de Guadarrama National Park (Madrid and Segovia, Spain): Taxonomy and Biogeography" Insects 12, no. 3: 266. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12030266