
Synthesis of the Brazilian Poduromorpha (Collembola: Hexapoda) with Special Emphasis on the Equatorial Oceanic Islands

 , ,
, ,
Abstract
:Simple Summary
Abstract
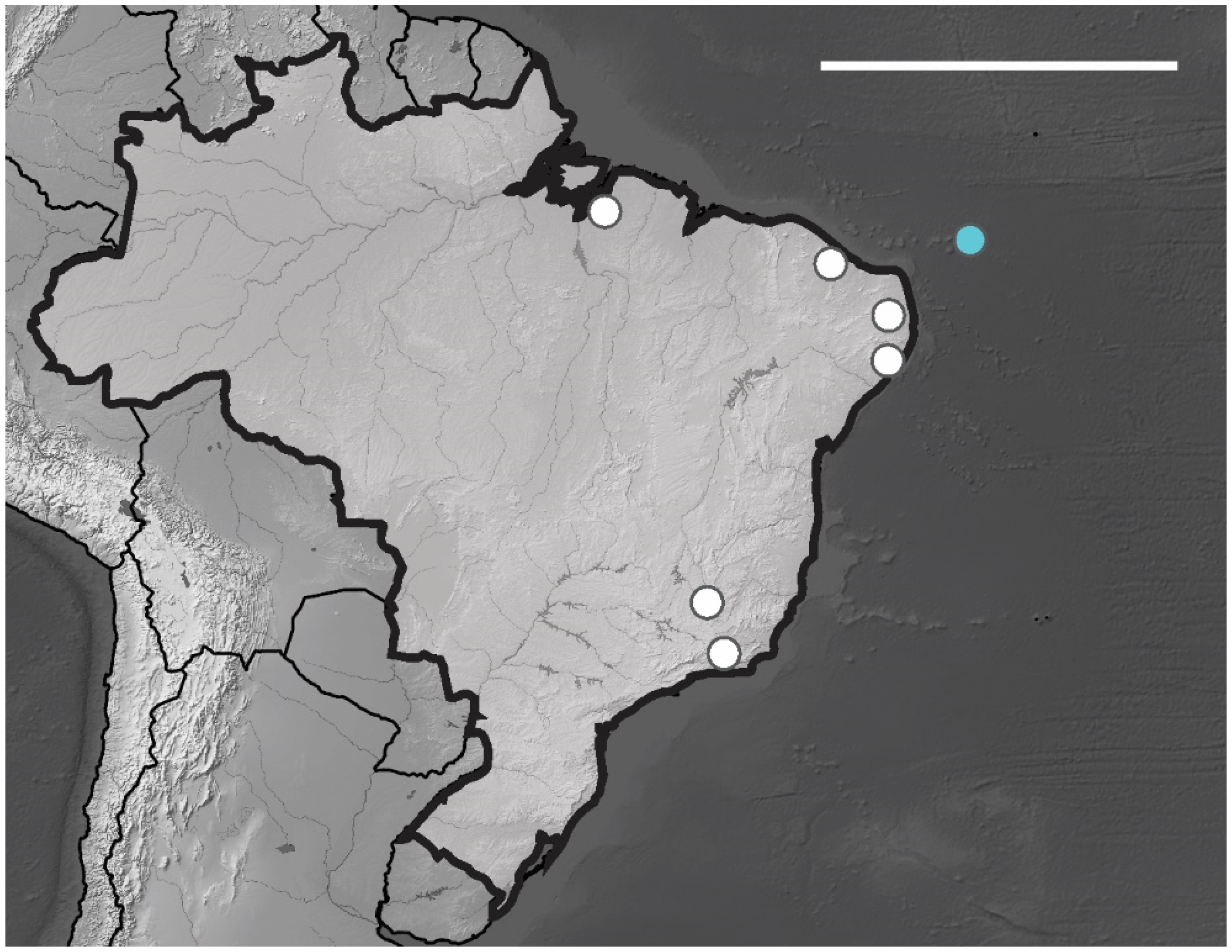
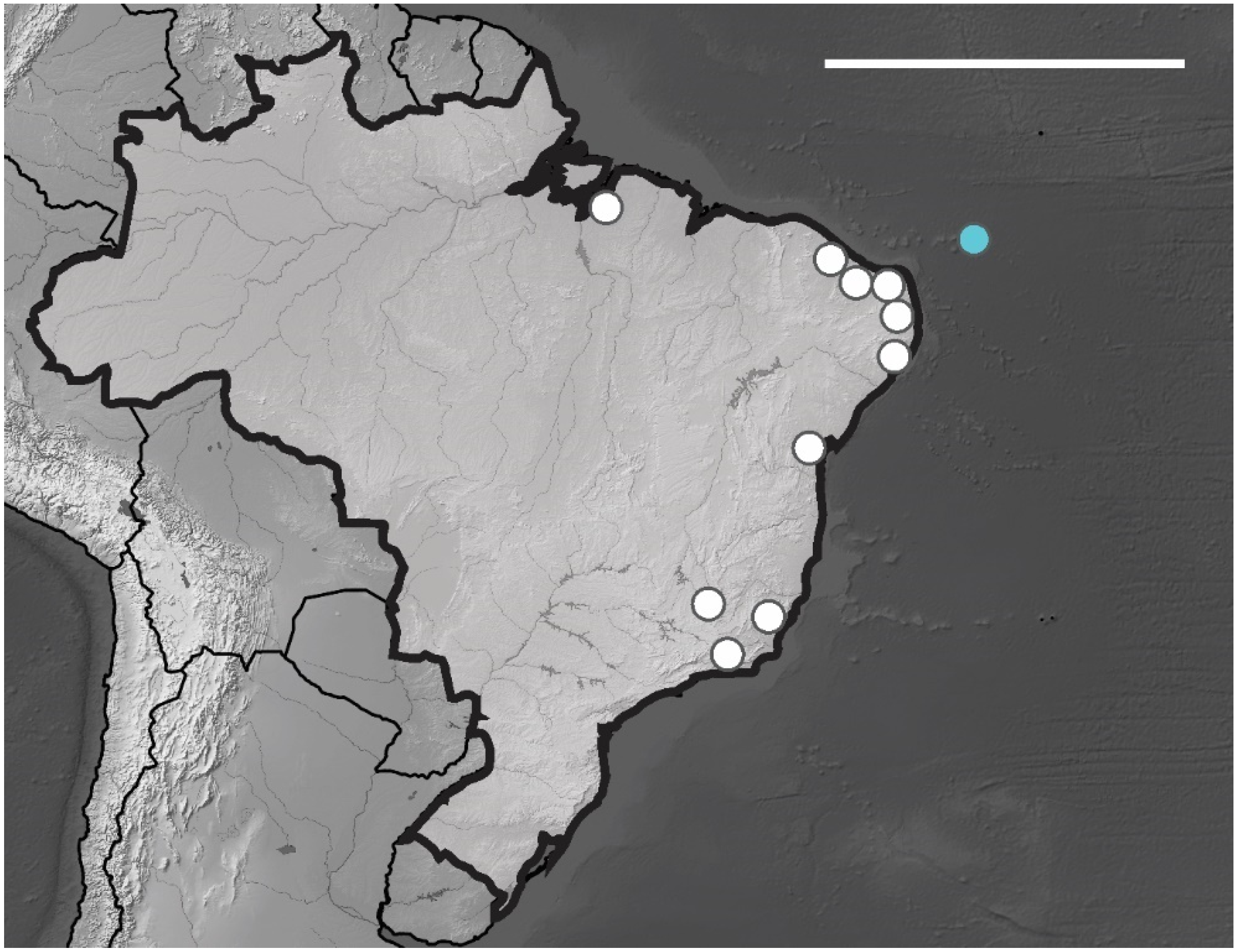
1. Introduction
2. Materials and Methods
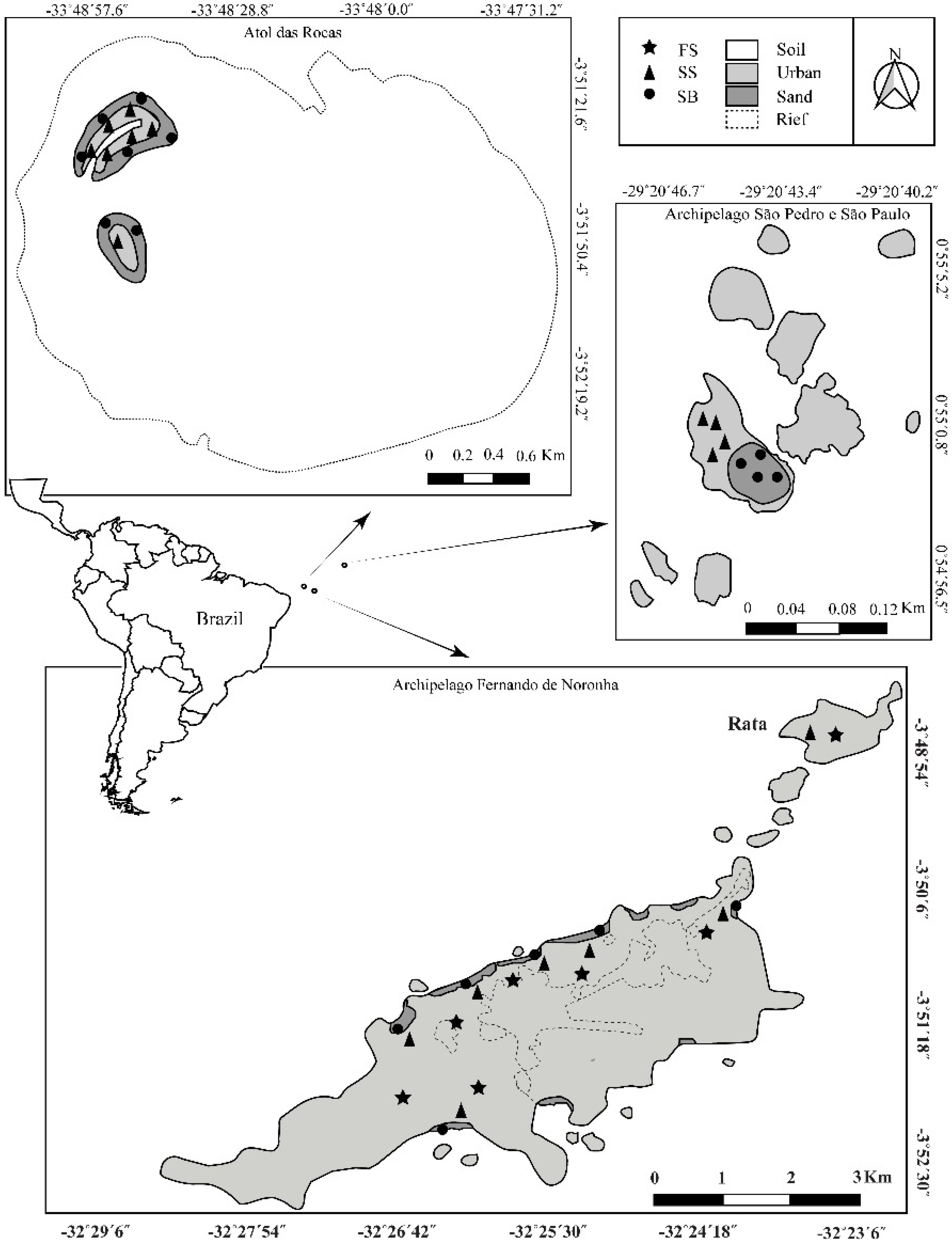
2.1. Poduromorpha Survey of the Brazilian Oceanic Islands
2.2. Sample Procedure
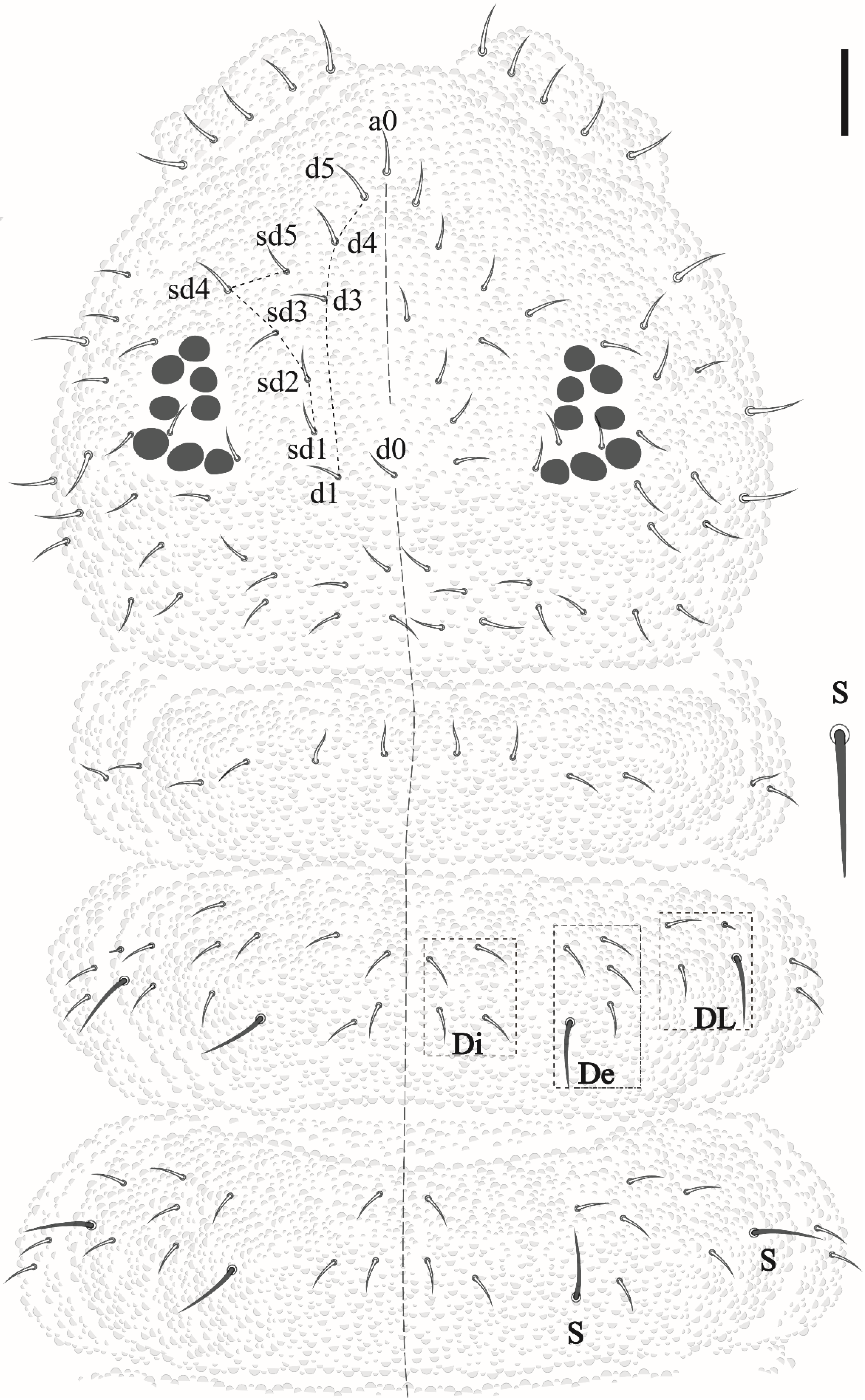
2.3. Taxonomic Procedure
2.4. Species List of Brazilian Poduromorpha
2.5. Abbreviations
3. Results
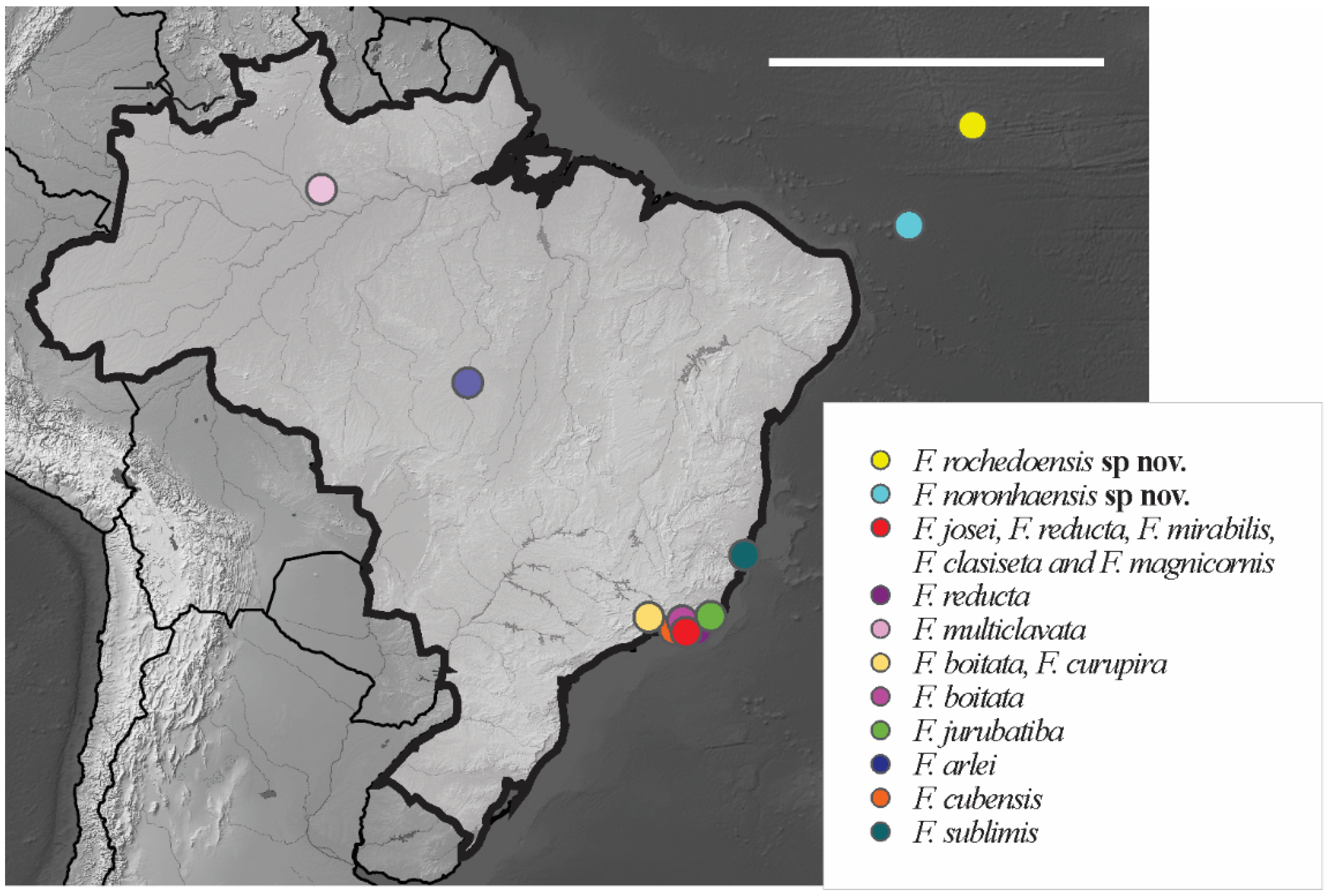
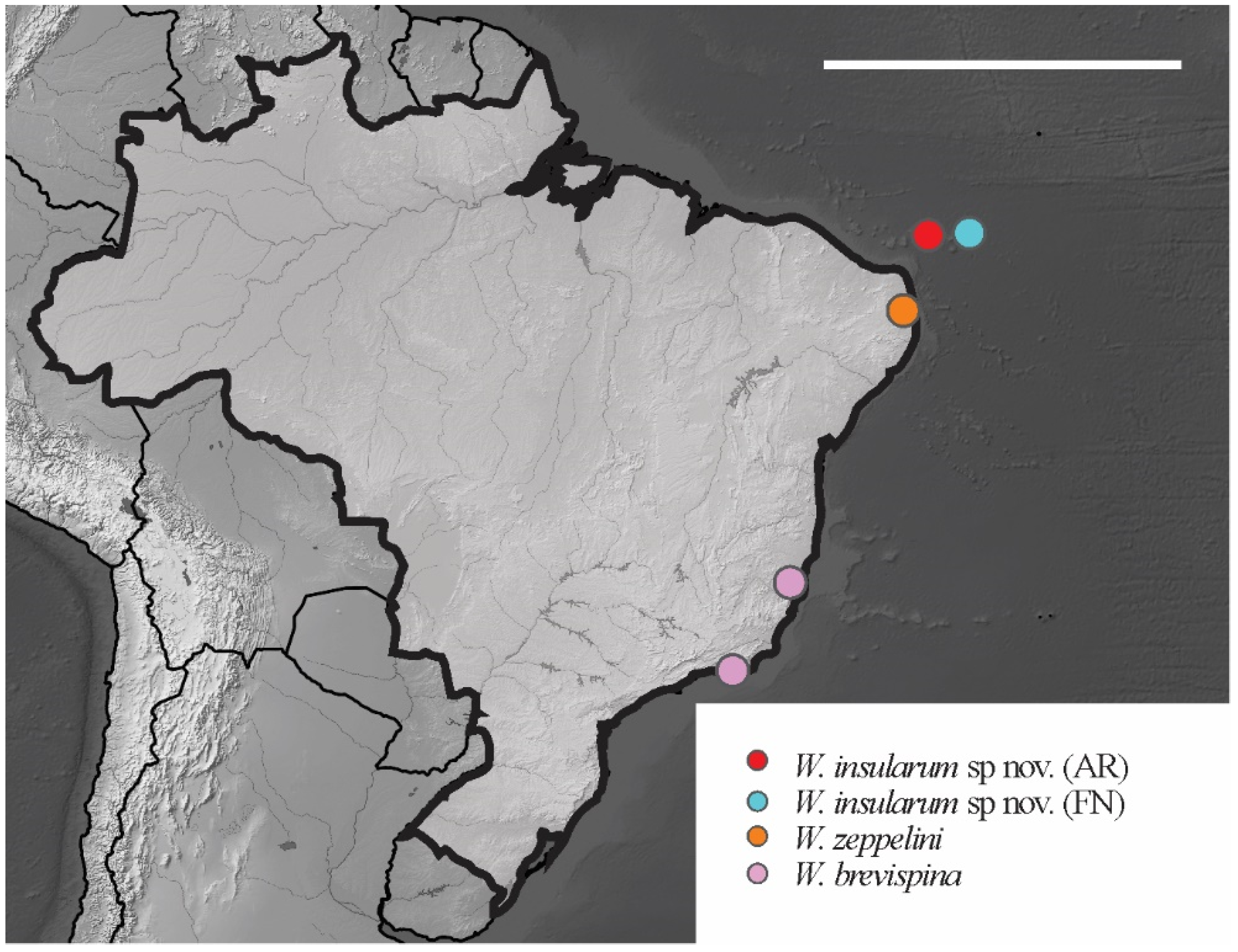
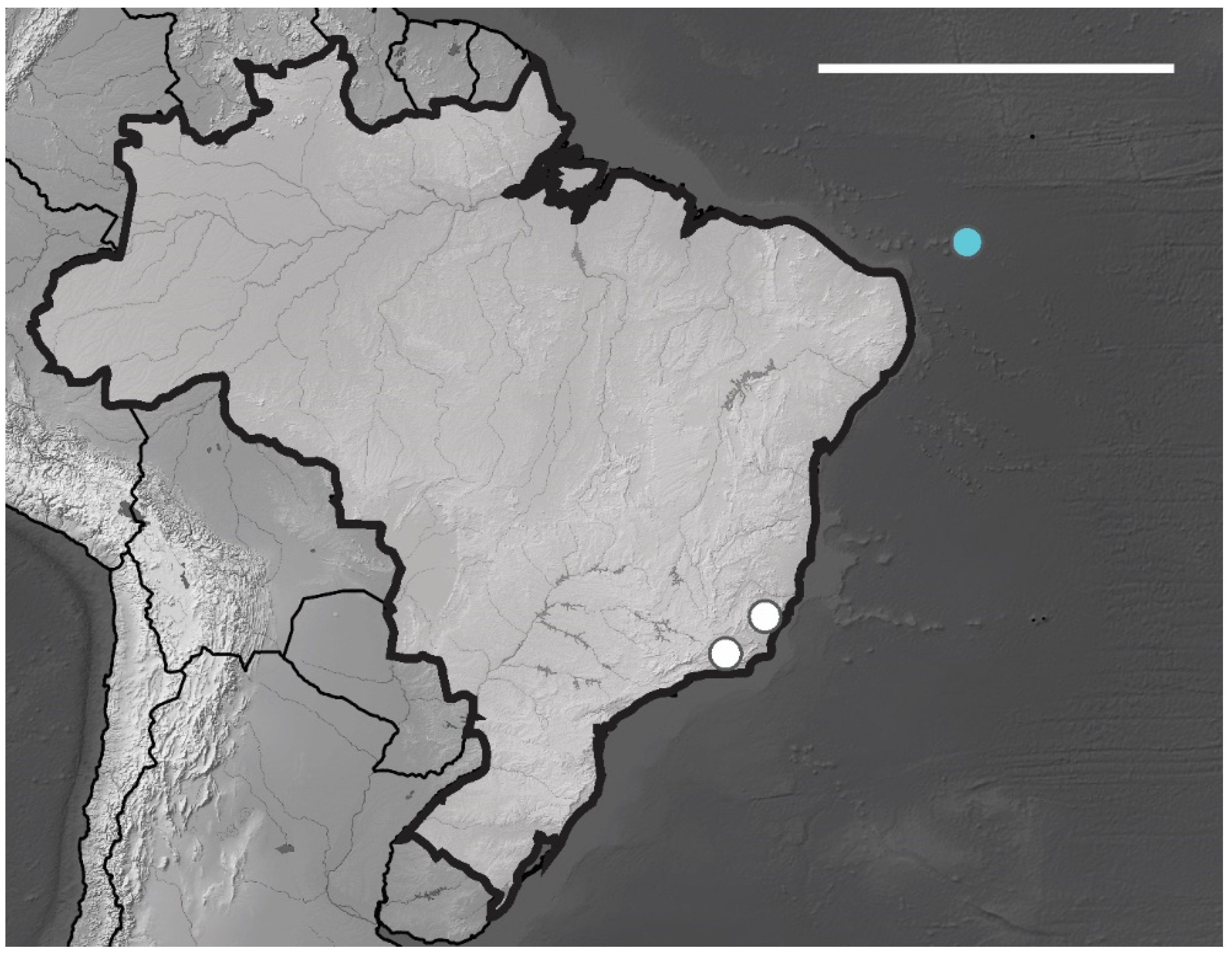
3.1. Poduromorpha Survey of the Brazilian Equatorial Oceanic Islands
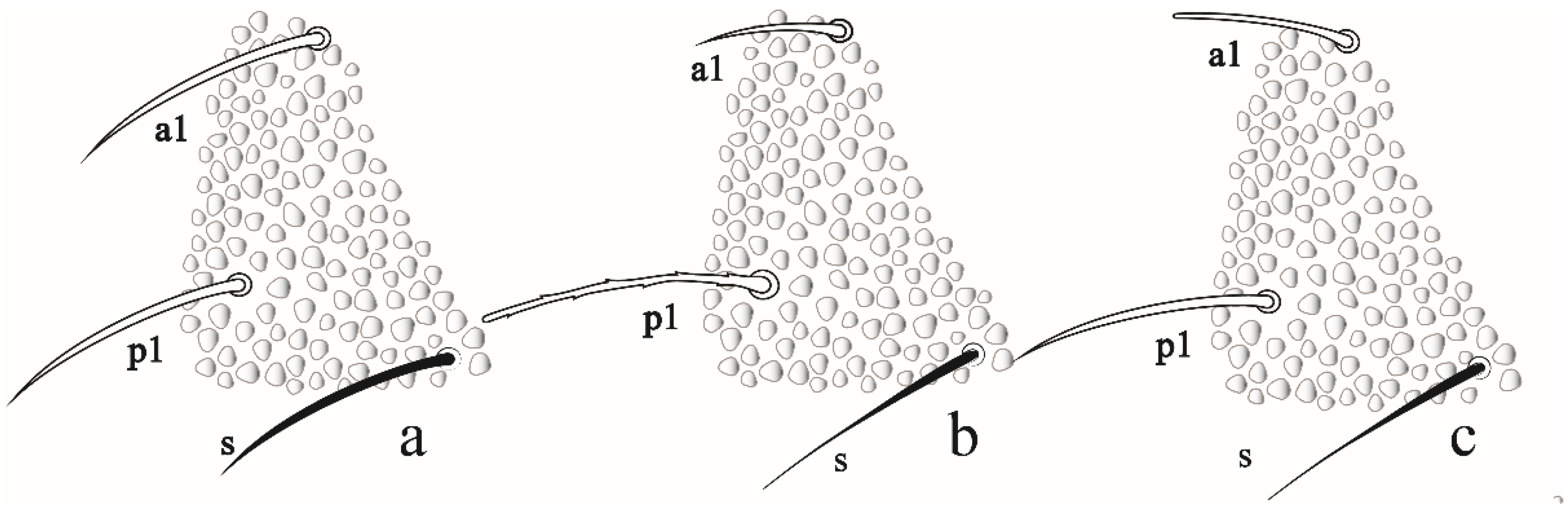
3.1.1. Neanuridae Börner, 1901
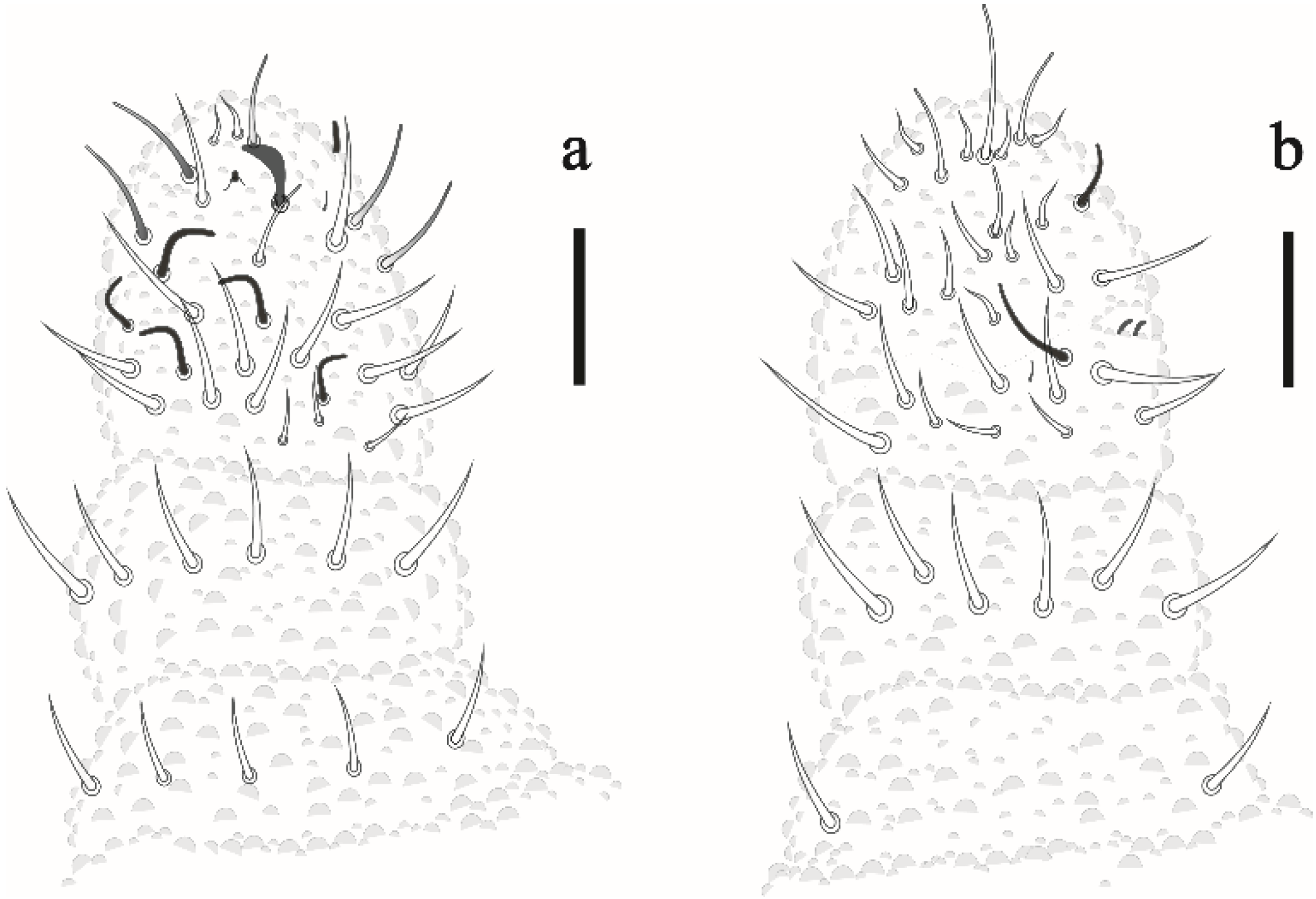
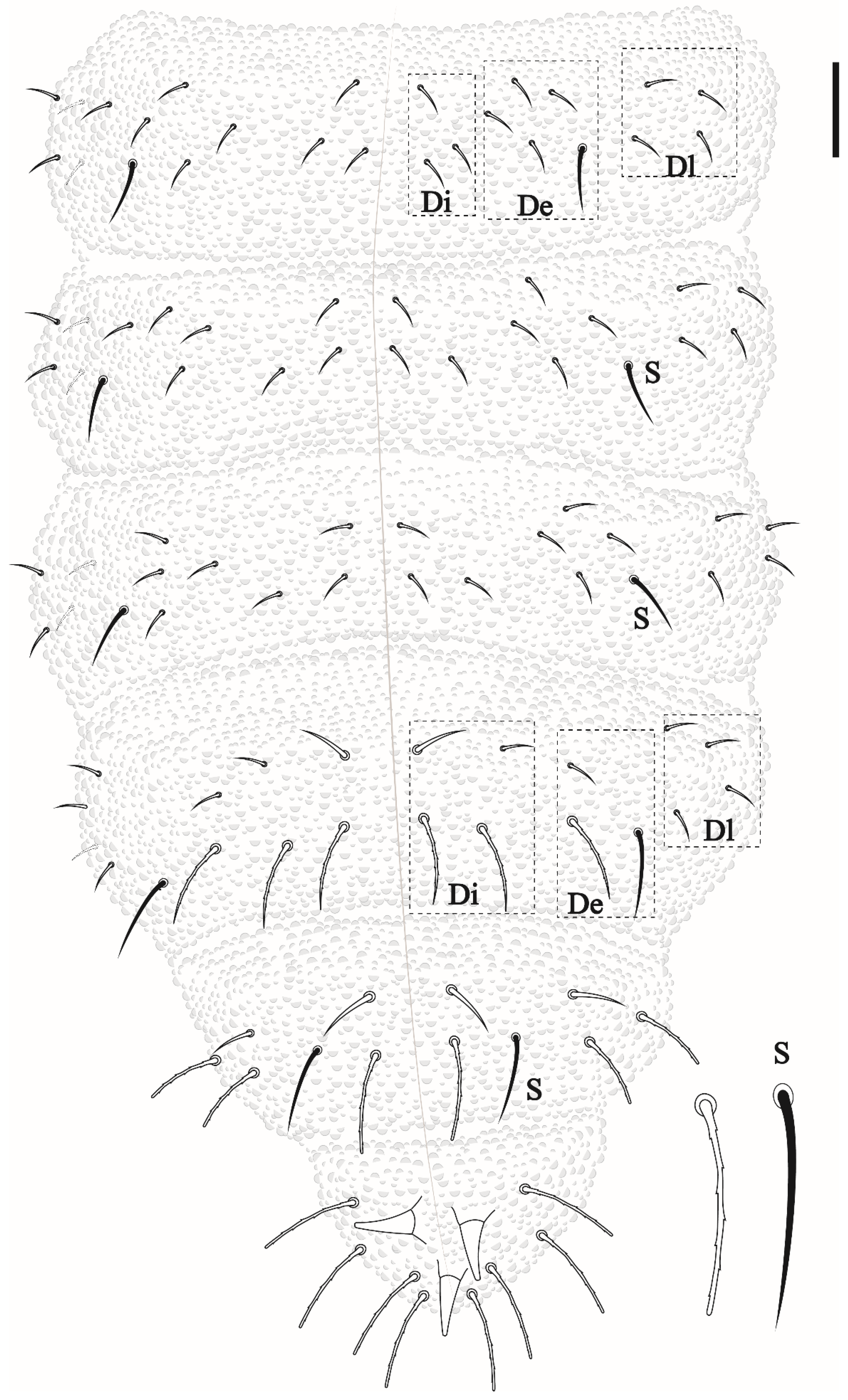
Friesea noronhaensis sp. nov. Lima and Zeppelini
Friesea rochedoensis sp. nov. Lima and Zeppelini
Identification Key for the Species of Friesea Recorded in Brazil
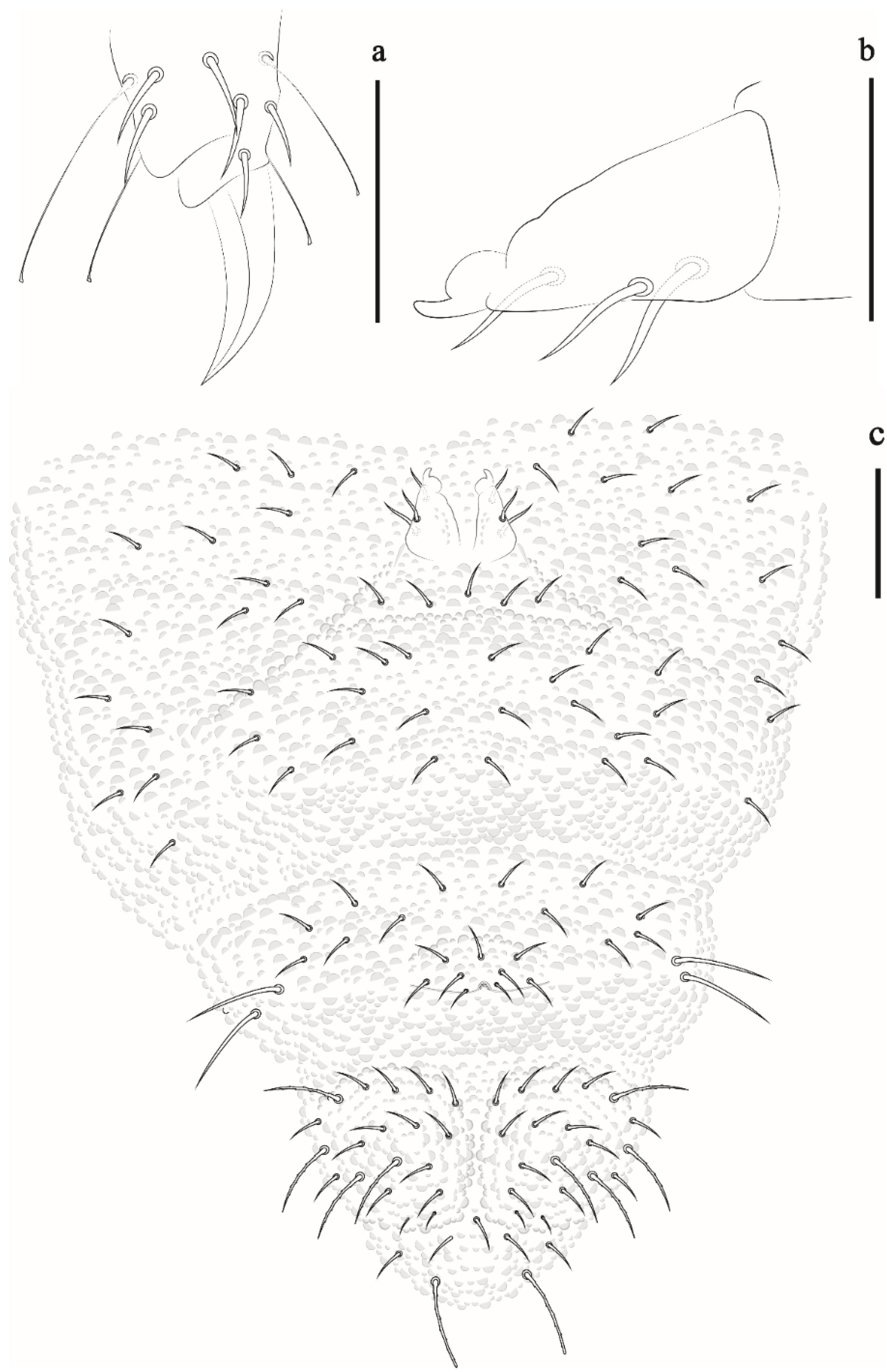
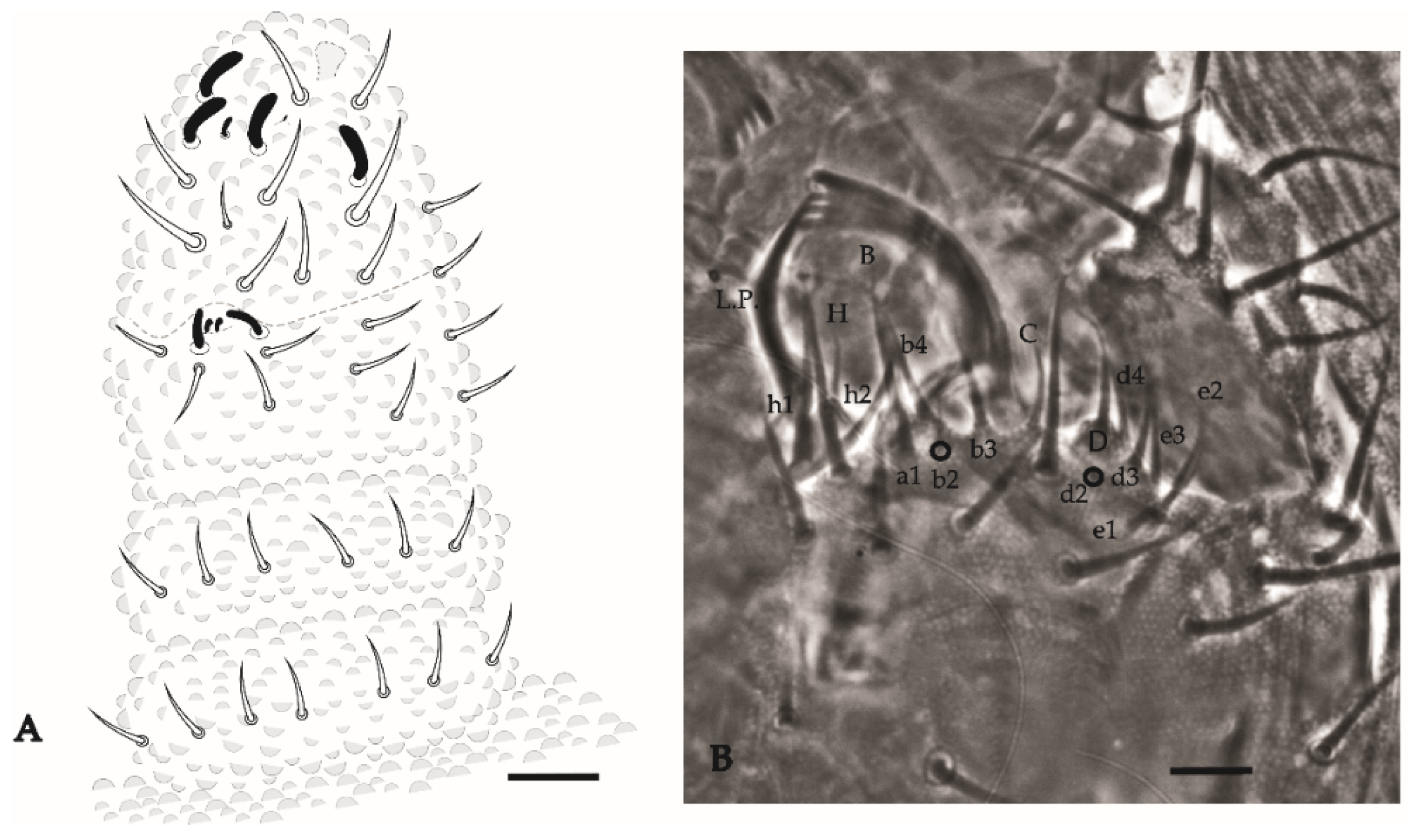
Arlesia albipes (Folsom, 1927)
Identification Key for the Species of Arlesia Handschin, 1942 Recorded in Brazil
3.1.2. Brachystomellidae Stach, 1949
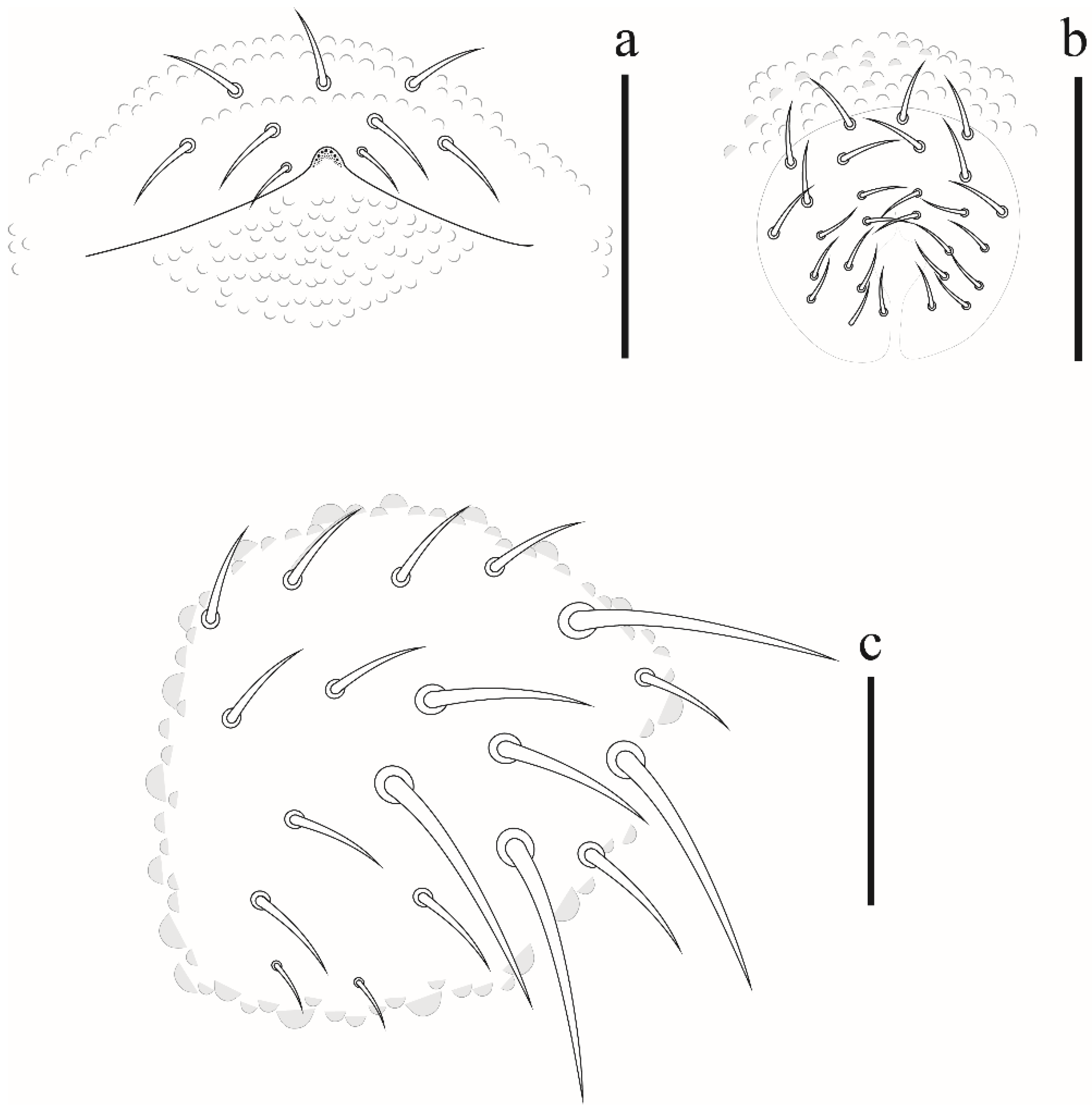
Brachystomella agrosa Wray, 1953
Identification Key for the Species of Brachystomella Ågren, 1903 Recorded in Brazil
3.1.3. Hypogastruridae Börner, 1906
Acherontiella sp.
Identification Key for the Species of Acherontiella Recorded in Brazil
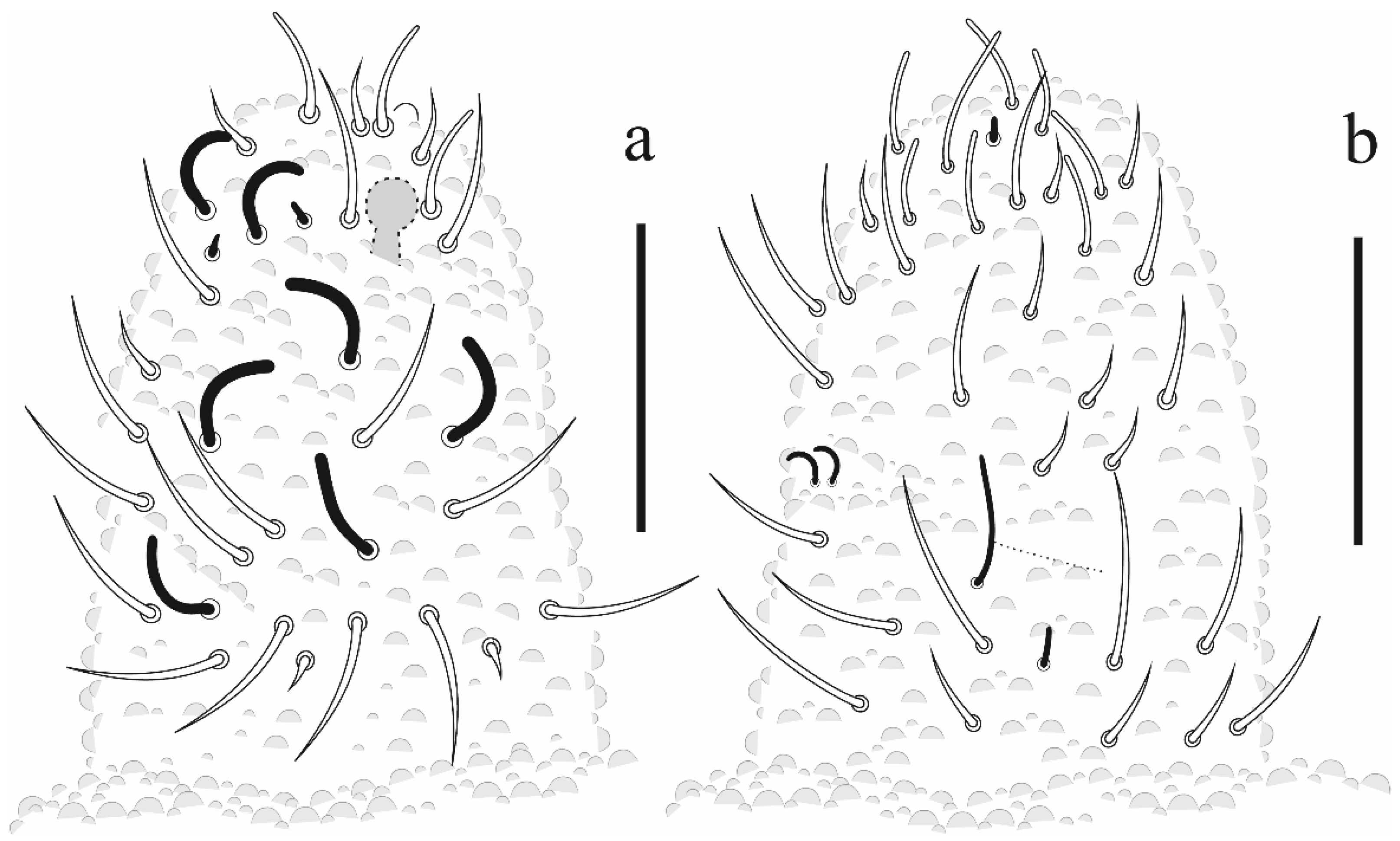
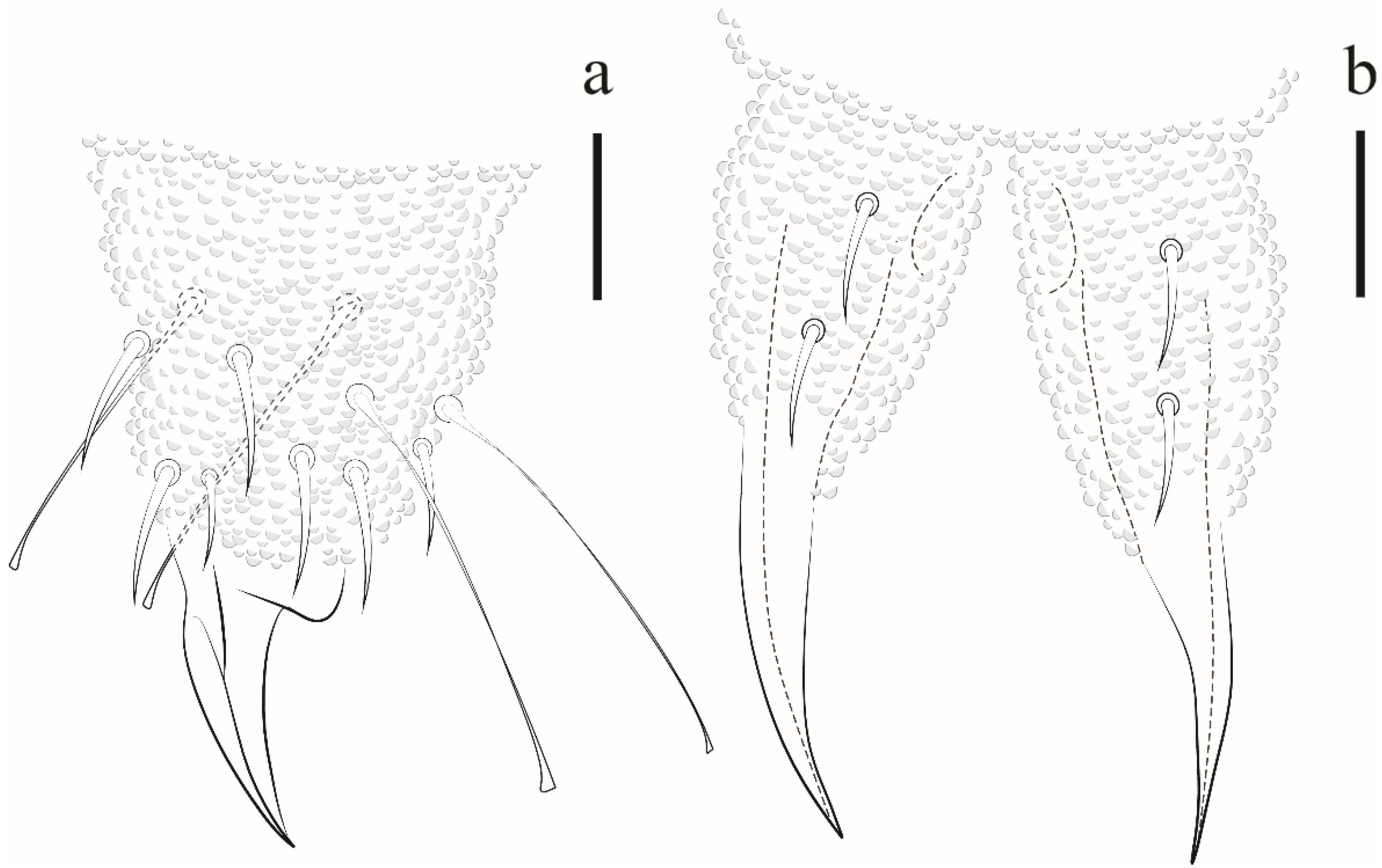
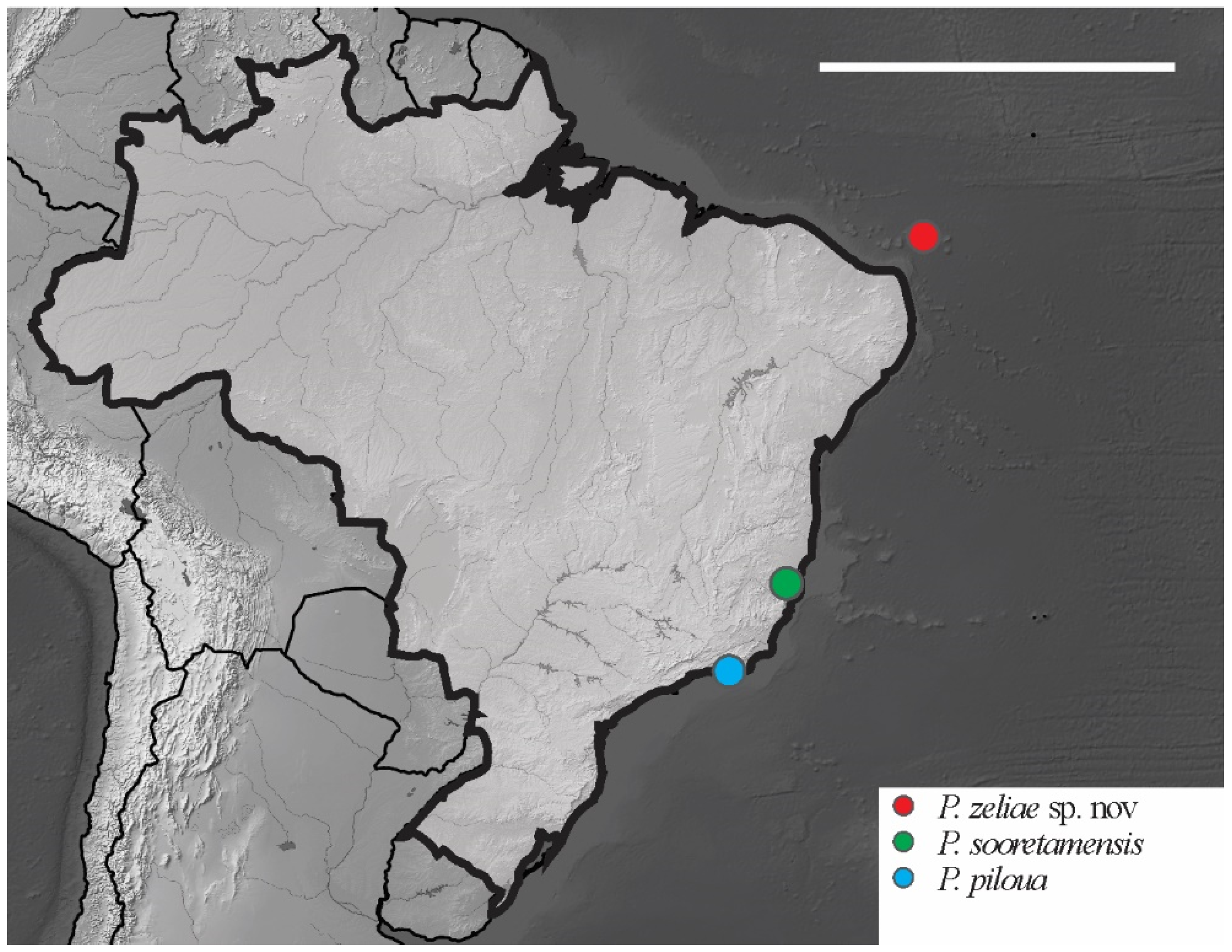
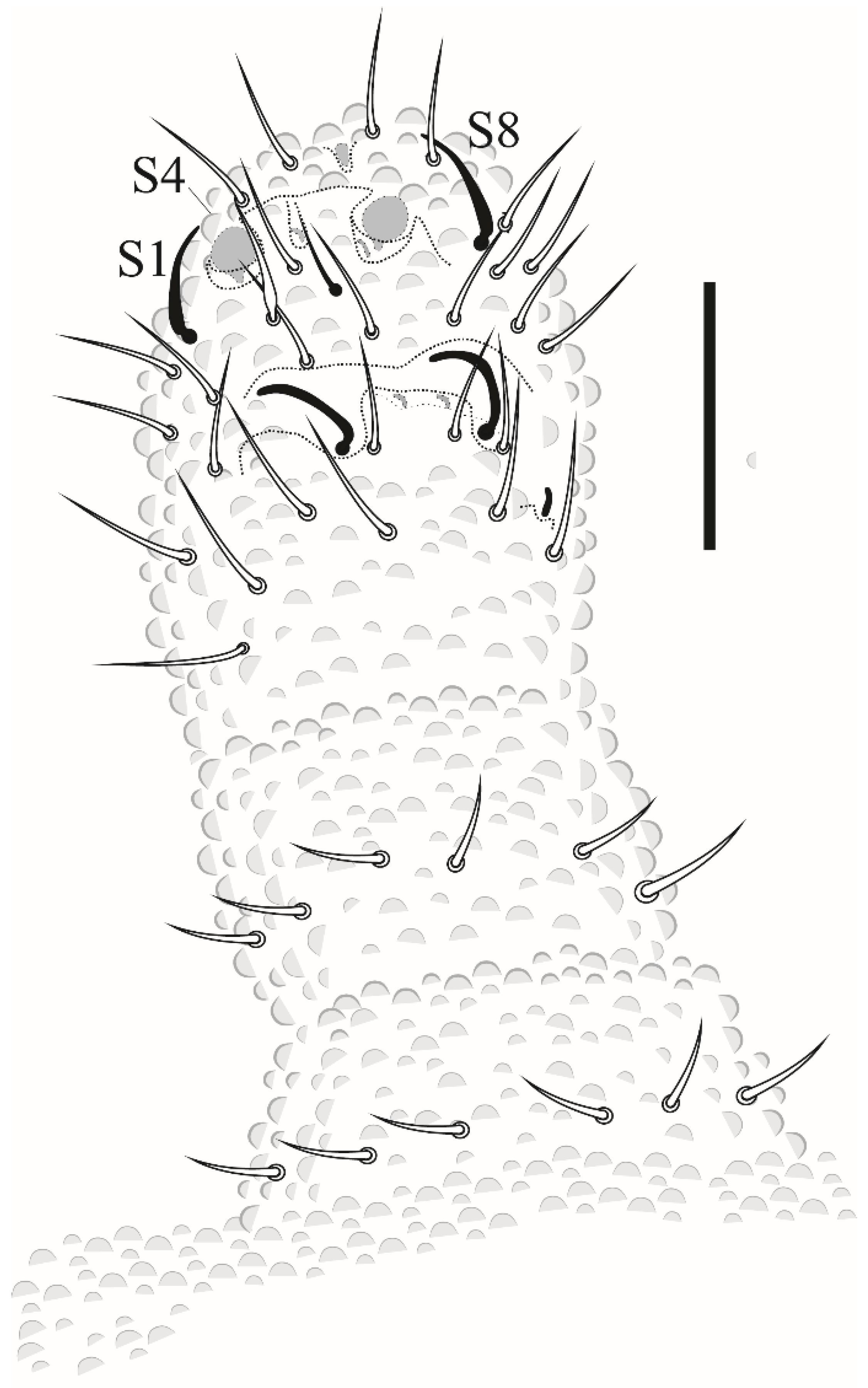
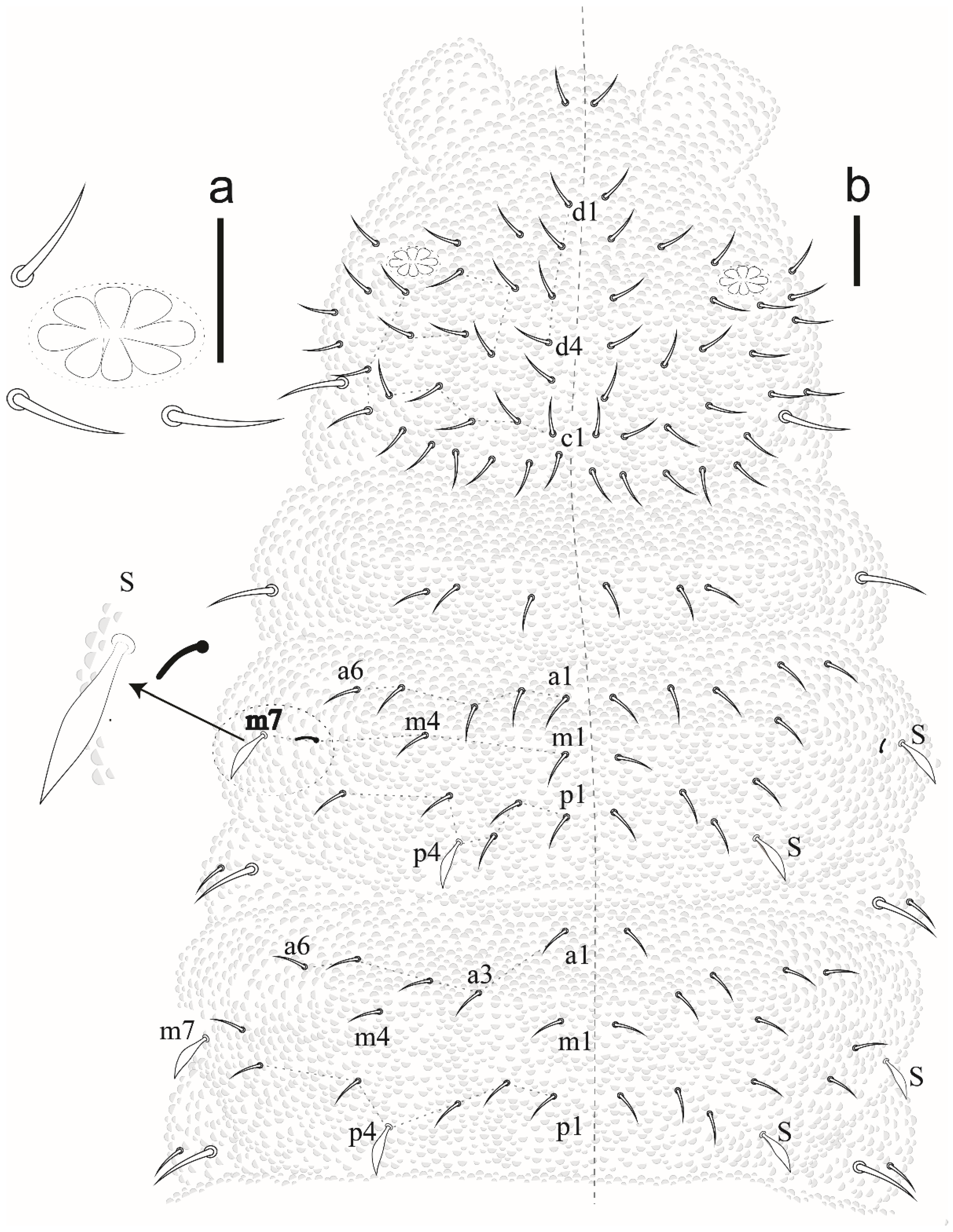
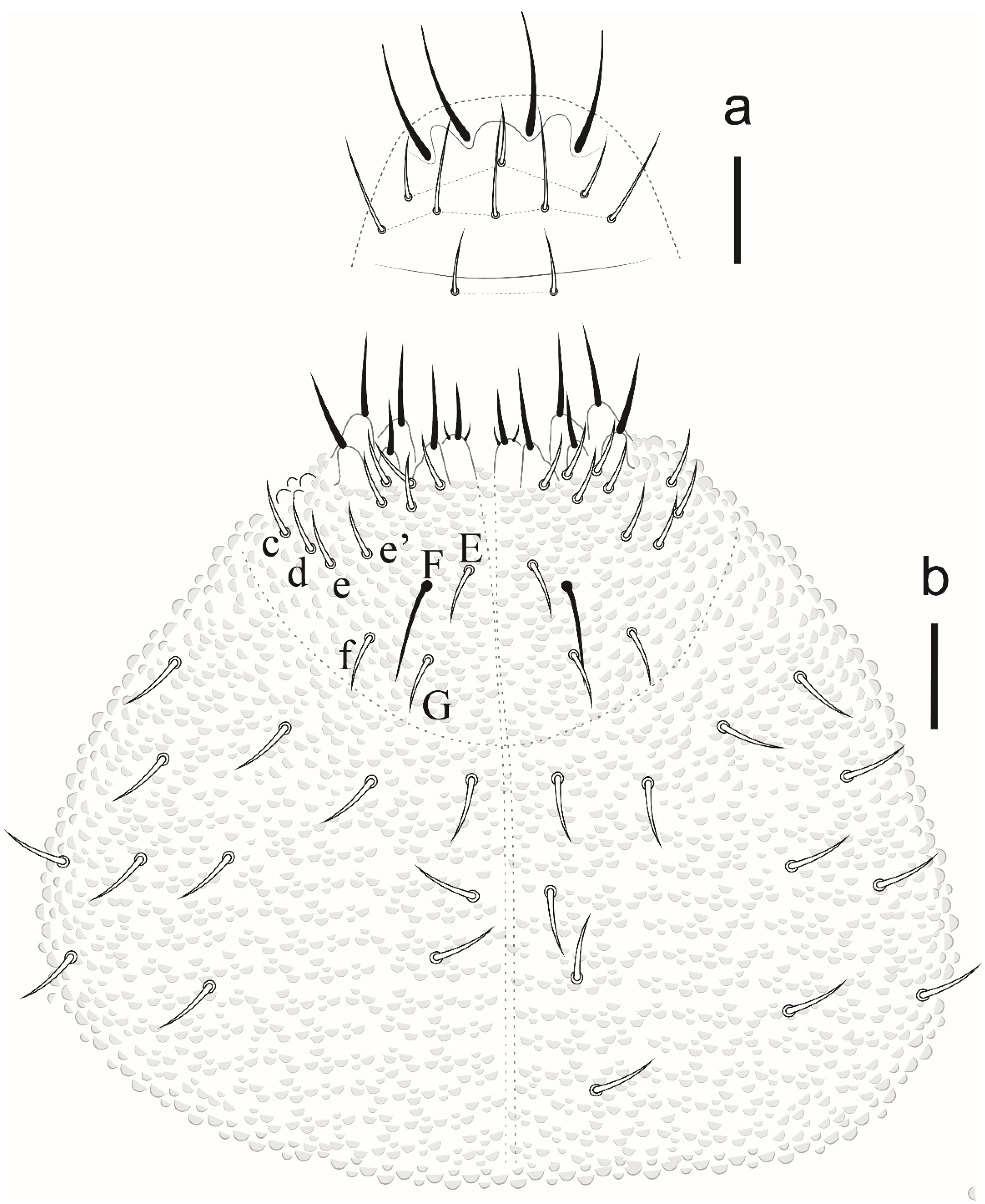
Paraxenylla zeliae sp. nov Lima and Zeppelini
Identification Key for Paraxenylla Murphy, 1965 Species Recorded in Brazil
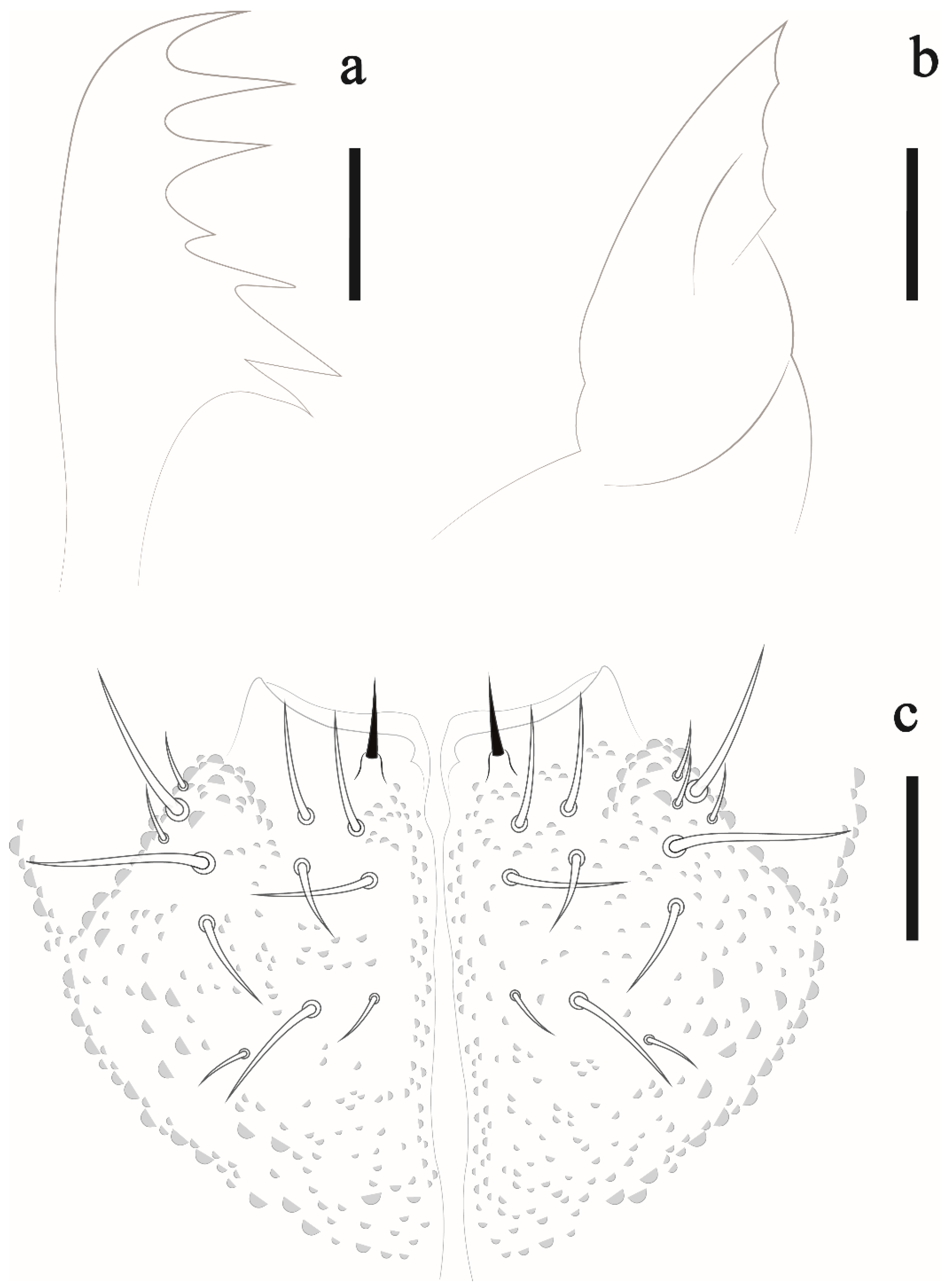
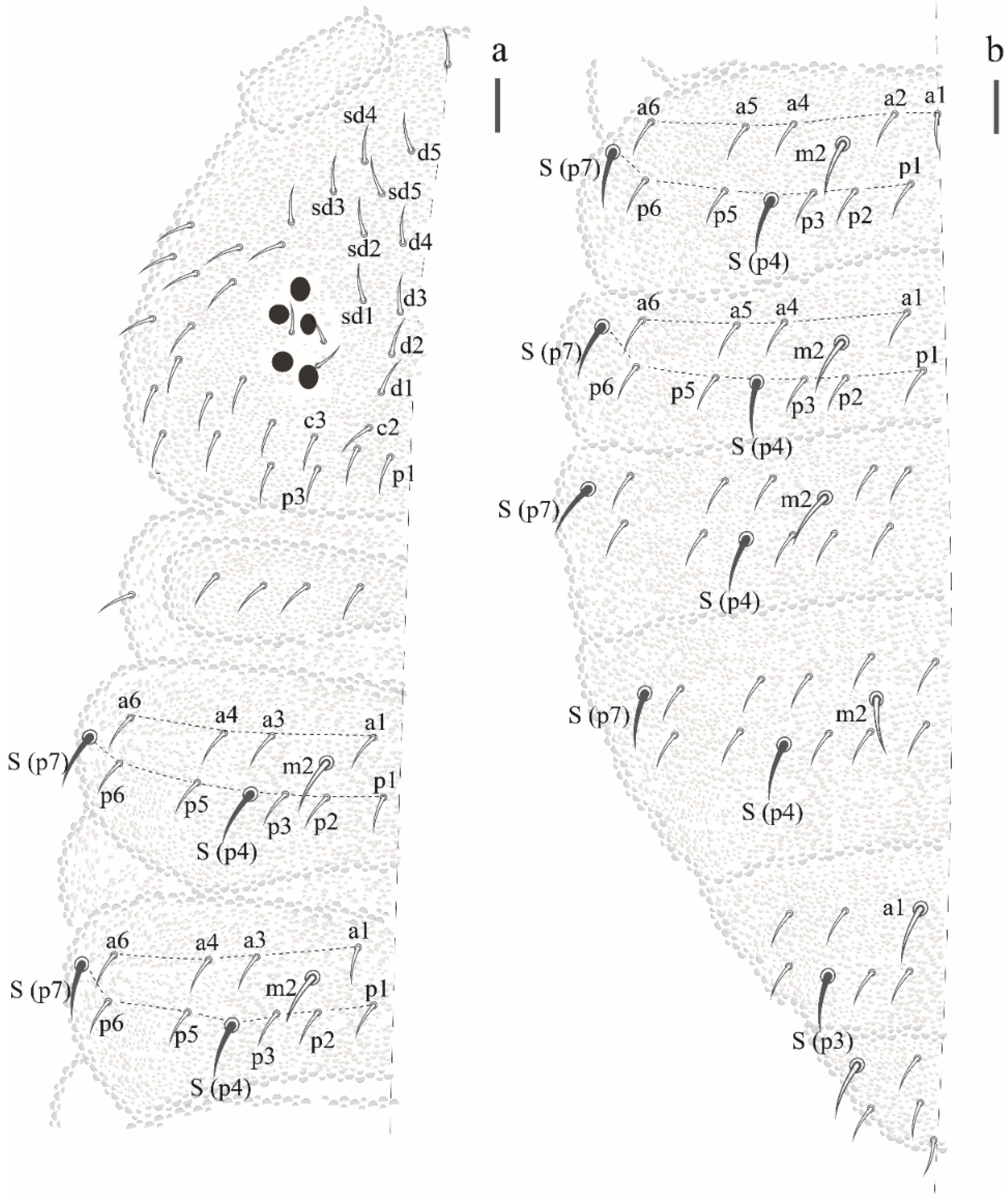
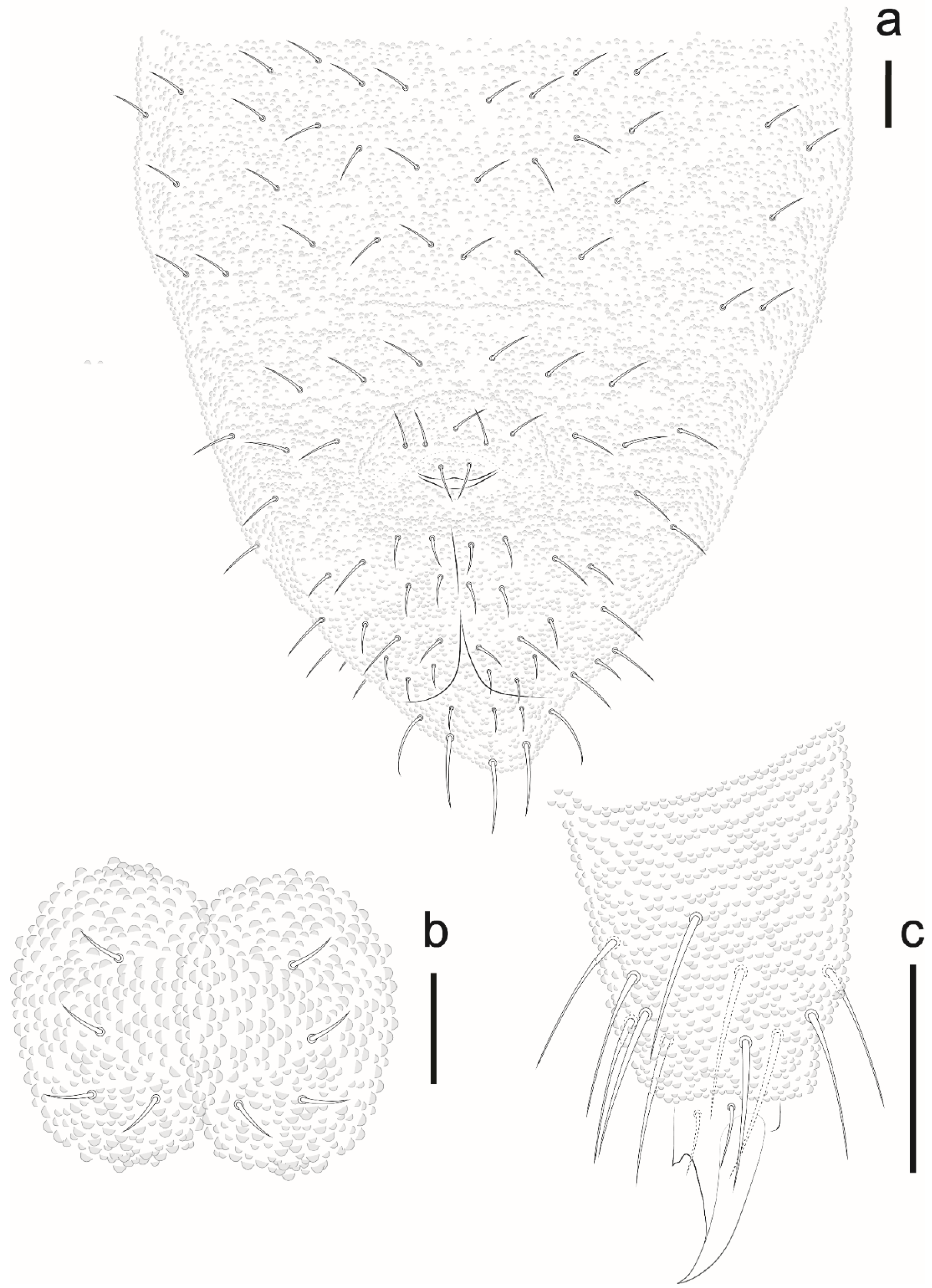
Willemia insularum sp. nov. Lima and Zeppelini
Identification Key for Willemia Species Recorded in Brazil
Xenylla yucatana Mills, 1938
Identification Key for Xenylla Tullberg, 1869 Species Recorded in Brazil
3.1.4. Isotogastruridae Thibaud and Najt, 1992
Isotogastrura mucrospatulata Palacios-Vargas, Lima and Zeppelini, 2013
3.2. Species List of Brazilian Poduromorpha
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bellinger, P.F.; Christiansen, K.A.; Janssens, F. Checklist of the Collembola of the World. Available online: http://www.collembola.org (accessed on 2 December 2019).
- Abrantes, E.A.; Bellini, B.C.; Bernardo, A.N.; Fernandes, L.H.; Mendonça, M.C.; Oliveira, E.P.; Queiroz, G.C.; Sautter, K.D.; Silveira, T.C.; Zeppelini, D. Errata Corrigenda and update for the “Synthesis of Brazilian Collembola: An update to the species list”. Zootaxa 2012, 3168, 1–21. [Google Scholar] [CrossRef]
- Lima, E.C.A.; Zeppelini, D. First survey of Collembola (Hexapoda: Entognatha) fauna in soil of Archipelago Fernando de Noronha, Brazil. Flor. Entomol. 2015, 98, 368–369. [Google Scholar] [CrossRef] [Green Version]
- Jordana, R.; Arbea, J.I.; Simón, C.; Luciáñez, M.J. Fauna Iberica, Collembola Poduromorpha; Museo Nacional de Ciencias Naturales: Madrid, Spain, 1997; Volume 8, pp. 1–807.
- D’Haese, C.A. Homology and morphology in Poduromorpha (Hexapoda, Collembola). Eur. J. Entomol. 2003, 100, 385–407. [Google Scholar] [CrossRef]
- Potapov, M.B.; Banasko, J.A.A. New species of springtail from Cuba with comments on the role of chaetotaxy in diagnostics of the Friesea (Collembola, Neanuridae) species. Zoolog. Zhur. 1985, 64, 1162–1167. [Google Scholar]
- Deharveng, L.; Bedos, A. Taxonomy and cladistics analysis of the Thai species of Friesea Dalla Torre 1895 (Collembola Ne-anuridae). Trop. Zool. 1991, 4, 287–315. [Google Scholar] [CrossRef]
- Weiner, W.M.; Bedos, A.; Deharveng, L. Species of the genus Friesea(Collembola, Neanuridae) from New Caledonia and Vanuatu. Zoosystema 2009, 31, 507–518. [Google Scholar] [CrossRef]
- Culik, M.P.; Filho, D.Z. Diversity and distribution of Collembola (Arthropoda: Hexapoda) of Brazil. Biodivers. Conserv. 2003, 12, 1119–1143. [Google Scholar] [CrossRef]
- Good, R. The Geography of Flowering Plants; Longman Group: London, UK, 1974; pp. 1–574. [Google Scholar]
- Christiansen, K.; Bellinger, P. The biogeography of Collembola. Bull. Entomol. Polog. 1995, 64, 279–294. [Google Scholar]
- Cassagnau, P. Les espèces européennes du genre Friesea [Collemboles Poduromorphes]. Bull. Soc. D’Hist. Nat. Toul. 1958, 93, 17–29. [Google Scholar]
- Zeppelini, D.; Queiroz, G.C.; Bellini, B.C. Collembola in Catálogo Taxonômico da Fauna do Brasil. Available online: http://fauna.jbrj.gov.br/fauna/faunadobrasil/379 (accessed on 8 August 2019).
- Folsom, J.W. Insects of the subclass Apterygota from Central America and the West Indies. Proc. U. S. Natl. Mus. 1927, 72, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Massoud, Z. Les Collemboles Poduromorphes du Surinam. Stud. Suriname Guyanas 1963, 20, 42–51. [Google Scholar]
- Mendonça, M.C.; Fernandes, L.H. Contribuição para o conhecimento do gênero Arlesia Handschin (Collembola: Neanuridae, Pseudachorutinae). Rev. Bras. Zool. 1999, 16, 1195–1201. [Google Scholar] [CrossRef] [Green Version]
- Wray, D.L. New Collembola from Puerto Rico. J. Agric. Univ. Puerto Rico 1953, 37, 140–150. [Google Scholar] [CrossRef]
- Massoud, Z.; Thibaud, J.M. Les Collemboles des Petites II—Neanuridae. Rev. D´Écól. Biol. Sol. 1980, 17, 591–605. [Google Scholar]
- Najt, J.; Thibaud, J.-M.; Weiner, W. Collemboles (Insecta) Poduromorphes de Guyanae Française. Mem. Mus. Nat. D´Hist. Natu. 1990, 4, 95–121. [Google Scholar]
- Mendonça, C.; Arlé, R. Variação quetotáxica em Brachystomella agrosa Wray, 1953 (Collembola, Neanuridae). Bol. Mus. Nac. 1992, 351, 1–9. [Google Scholar]
- Weiner, W.M.; Najt, J. Species of Brachystomella (Collembola: Brachystomellidae) from the Neotropical region. Eur. J. Entomol. 2001, 98, 387–413. [Google Scholar] [CrossRef] [Green Version]
- Fjellberg, A. The Labial Palp in Collembola. Zool. Anz. 1999, 237, 309–330. [Google Scholar]
- D’Haese, C.; Thibaud, J.-M. Description and phylogenetic position of a new Willemia species (Collembola: Hypogas-truridae) from the littoral coast of Brazil. Zootaxa 2011, 2932, 33–40. [Google Scholar] [CrossRef]
- Bu, Y.; Potapov, M.B.; Gao, Y. Littoral Willemia (Collembola: Hypogastrurudae) of china with a description of two new species and a new case of convergente between Hypogastruridar and Onychiuridae. Flor. Entomol. 2015, 95, 580–586. [Google Scholar] [CrossRef]
- Zon, S.D.; Bedos, A.; D´Haese, C. Phylogeny of the genus Willemia (Collembola: Hypogastruridae) and biogeography of the W. buddenbrocki-group with description of a new species from Ivory Coast (western Africa). Zootaxa 2015, 3980, 230–240. [Google Scholar] [CrossRef] [Green Version]
- D´Haese, C. Is Psammophily and Evolutionary dead end? A phylogenetic test in the genus Willemia (Collembola: Hypogas-truriade). Cladistics 2000, 16, 255–273. [Google Scholar] [CrossRef]
- Mills, H.B. Collembola from Yucatan caves. Carn. Inst. Washing. Publ. 1938, 491, 183–190. [Google Scholar]
- Murphy, D.H. Collembola Poduromorpha from the Gambia (West Africa). J. Zool. 2010, 146, 388–411. [Google Scholar] [CrossRef]
- Gama, M.M. Notes taxonomiques et lignées généalogiques de quarante deux espèces et sous-espèces du genre Xenylla (Insecta: Collembola). Mem. Est. Mus. Zool. Univers. Coimbra. 1969, 308, 1–61. [Google Scholar]
- André, H.M. Redescription of Xenylla yucatana Mills, 1938 (Collembola, Hypogastruridae). Fol. Entomol. Mex. 1988, 74, 69–77. [Google Scholar]
- Christiansen, K.; Bellinger, P. Insects of Hawaii. A Manual of the Insects of the Hawaiian Islands, Including an Enumeration of the Species and Notes on Their Origin, Distribution, Hosts, Parasites; University of Hawaii Press: Honolulu, HI, USA, 1992; p. 445. [Google Scholar]
- Potapov, M.B.; Bu, Y.; Gao, Y. First record of the littoral family Isotogastruridade (Collembola) in Asia. Zookeys 2011, 136, 23–29. [Google Scholar]
- Thibaud, J.-M. Recent advances and synthesis in biodiversity and biogeography of arenicolous Collembola. Ann. Soc. Entomol. Fr. 2007, 43, 181–185. [Google Scholar] [CrossRef]
- Palacios-Vargas, J.G.; Lima, E.C.A.; Zeppelini, D. A new species of Isotogastrura (Collembola: Isotogastruridae) from North-eastern Brazil. Flor. Entomol. 2013, 96, 1579–1587. [Google Scholar] [CrossRef]
- Cassagnau, P.; Rapoport, E.H. Collemboles d’Amérique du Sud, I Poduromorphes. Biol. Amér. Aust. 1962, 1, 139–184. [Google Scholar]
- Börner, C. Das System der Collembolen nebst Beschreibung neuer Collembolen des Hamburger Naturhistorischen Museums. Mitt. Natu. Mus. Hamb. 1906, 23, 147–188. [Google Scholar]
- Fernandes, L.H.; Mendonça, M.C. Collembola Poduromorpha do litoral de Maricá, Rio de Janeiro, Brasil. Rev. Bras. Zool. 2004, 21, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Arlé, R. Sôbre Pseudachorutidae da região amazônica (Collembola). At. Soc. Biol. Rio J. 1962, 6, 1–3. [Google Scholar]
- Queiroz, G.C.; Weiner, W.M. A new species of Brachystomella (Collembola: Brachystomellidae) from the Atlantic Forest of southeast Brazil. Zootaxa 2011, 2885, 65–68. [Google Scholar] [CrossRef]
- Bellini, B.; Santos, N.; Souza, P.; Weiner, W. Two new species of Brazilian springtails (Hexapoda: Collembola) with comments on Neotropical Brachystomella Ågren and Seira (Lepidocyrtinus) Börner. Insect Syst. Evol. 2019, 50, 297–326. [Google Scholar] [CrossRef]
- Dos Santos-Rocha, I.M.; Andreazze, R.; Bellini, B.C. Registros de Collembola (Arthropoda, Hexapoda) no Estado do Rio Grande do Norte, Brasil. Biota Neotrop. 2011, 11, 167–170. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, L.H.; Mendonça, M.C. A new species of Xenylla (Collembola: Poduromorpha: Hypogastruridae) with new records for the state of Espirito Santo, southeastern Brazil. Zoologia 2010, 27, 123–127. [Google Scholar] [CrossRef]
- Da Silveira, T.C.; de Mendonça, M.C. Checklist of Poduromorpha (Collembola: Hexapoda) from Rio de Janeiro, Brazil. Zootaxa 2019, 4656, 572–590. [Google Scholar] [CrossRef] [PubMed]
- Brito, R.A.; Lima, E.C.A.; Ferreira, A.S.; Oliveira, J.V.L.C.; Amorim, T.P.L.; Machado, T.G.; Lopes, B.C.H.; Costa, I.A.; Alves, J.L.S.; Brito, N.P.; et al. A Catalogue of Species of Collembola (Hexapoda: Ellipura) Deposited in Coleção de Referência de Fauna de Solo of the Universidade Estadual da Paraíba, Brazil. Fla. Entomol. 2017, 100, 9–14. [Google Scholar] [CrossRef]
- Mendonça, M.C.; Fernandes, L.H.; Abrantes, E.A.; Queiroz, G.C.; Bernardo, A.N.; Silveira, T.C. Fauna colembológica do Estado do Rio de Janeiro, Brasil. Arq. Mus. Nac. 2009, 67, 265–274. [Google Scholar]
- Arlé, R. Collembola Arthropleona do Brasil oriental e central. Arq. Mus. Nac. 1959, 49, 155–211. [Google Scholar]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of this Group of Insects. Families: Neogastruridae and Brachystomellidae. Acta Monogr. Musei Hist. Nat. Kraków 1949, 2, 1–341. [Google Scholar]
- Arlé, R. Collemboles nouveaux de Rio de Janeiro. Ann. Acad. Bras. Ciênc. 1939, 11, 25–32. [Google Scholar]
- De Mendonça, M.C.; Fernandes, L.H.; Abrantes, E.A. Two new species of Folsomiella Bonet and redescription of F. albida (Arlé, 1959) (Collembola, Brachystomellidae). Zootaxa 2005, 1006, 11. [Google Scholar] [CrossRef]
- Mendonça, M.C.; Fernandes, L.H. A new genus of Brachystomellinae from Brazil (Collembola: Neanuridae). Bol. Mus. Nac. 1997, 379, 1–7. [Google Scholar]
- Queiroz, G.C.; Mendonca, M.C. New species of Brachystomellidae and characterization of Micronella porcus (Denis, 1933) from Brazil. ZooKeys 2013, 316, 81–98. [Google Scholar] [CrossRef]
- Mendonça, M.C.; Fernandes, L.H. Nova espécie de Rapoportella Ellis & Bellinger, 1973 do Brasil (Collembola, Neanuridae). Bol. Mus. Nac. 1995, 360, 1–6. [Google Scholar]
- Palacios-Vargas, J.G.; Gnaspini-Netto, P. A new Brazilian species of Acherontides (Collembola: Hypogastruridae), with notes on its ecology. Jour. Kan. Entom. Soc. 1992, 65, 443–447. [Google Scholar]
- De Lima, E.C.A.; Stievano, L.C.; Zeppelini, D. A new species of Acherontides (Collembola: Hypogastruridae) from Brazil, with an identification key. Zootaxa 2019, 4623, 141–150. [Google Scholar] [CrossRef]
- Culik, M.P.; Martins, D.D.S.; Ventura, J.A. Collembola (Arthropoda: Hexapoda) communities in the soil of papaya orchards managed with conventional and integrated production in Espírito Santo, Brazil. Biota Neotrop. 2006, 6, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, L.H.; Bellini, B.C.; de Mendonça, M.C. Revision of the genus Austrogastrura Thibaud & Palacios-Vargas, 1999 (Collembola: Poduromorpha: Hypogastruridae). Zootaxa 2010, 2542, 61–68. [Google Scholar] [CrossRef]
- Arlé, R. Collembola, anexo N. 2, ao relatório da excursão científica do Instituto Oswaldo Cruz realizada na zona da E. F. N. O. B., em outubro de 1938. Boletim Biol. 1939, 4, 295–300. [Google Scholar]
- Arlé, R. Uma nova espécie de Onychiurus (Collembola-Onychiuridae) de ocorrência periódica em Belém (Pará). Bol. Mus. Para. Emilio Goeldi 1970, 72, 1–11. [Google Scholar]
- Thibaud, J.-M.; Palacios-Vargas, J.G. Brazilian Collembola from littoral sand with description of Austrogastrura gen. n. and Isotomodes carioca sp. n. [Hypogastruridae; Isotomidae]. Rev. Franç. Entom. 1999, 21, 25–31. [Google Scholar]
- Moniez, R. Isotoma pallida, collembole nouveau de Brésil. Rev. Biol. Nord Fr. 1984, 6, 354. [Google Scholar]
- Queiroz, G.; Mendonça, M. New Hypogastruridae (Collembola) and first record of Cephalachorutes (Collembola: Neanuridae) for the Neotropical Region. Insect Syst. Evol. 2016, 47, 33–52. [Google Scholar] [CrossRef]
- Palacios-Vargas, J.; Cipola, N.; Bellini, B. Two new species of Ceratophysella (Collembola: Hypogastruridae) from Brazil with a key to South American species of the genus. Insect Syst. Evol. 2018, 49, 207–230. [Google Scholar] [CrossRef]
- Denis, J.R. Sur Isotoma pallida Moniez et un autre collembole nouveau du Brésil: Hypogastrura moniezi n. sp. Bull. Soc. Zool. Fr. 1923, 48, 95–98. [Google Scholar]
- Fernandes, L.H.; Mendonça, M.C. Collembola Poduromorpha de áreas preservadas e impactadas do litoral sudeste do Brasil. Rev. Bras. Zool. 2007, 24, 777–785. [Google Scholar] [CrossRef]
- Queiroz, G.C.; Deharveng, L. Description of a new species of Paraxenylla (Collembola, Hypogastruridae) from Brazil, with comments on the genus. Zootaxa 2008, 1858, 1–8. [Google Scholar] [CrossRef]
- Fernandes, L.H.; Mendonça, C. Uma nova espécie de Schoettella Schaeffer (Collembola, Hypogastruridae). Rev. Bras. Entom. 1998, 41, 453–457. [Google Scholar]
- Oliveira, E.P.; Thibaud, J.-M. Un nouveau genre d’insecte collembole Hypogastruridae d’Amazonie. Amazoniana 1988, 10, 299–302. [Google Scholar]
- Gama, M.M. Systématique évolutive des Xenylla. X. Espèces provenant du Brésil, du Vietnam et de Madagscar (Insecta: Collembola). Ciênc. Biol. 1978, 4, 45–48. [Google Scholar]
- Neves, A.C.R.; Mendonça, M.C. A new species of Xenylla Tullberg, 1869 (Collembola: Hypogastruridae) from Central Amazon, Brazil. Zootaxa 2017, 4347, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Gama, M.M.; Oliveira, E.P. Systématique évolutive des Xenylla. XVI. Description d’une nouvelle espèce provenant d’Amazonie (Insecta: Collembola). Amazoniana 1994, 13, 205–208. [Google Scholar]
- Da Silveira, T.C.; de Mendonça, M.C.; Da-Silva, E.R. A second new species of Isotogastrura Thibaud & Najt (Collembola: Isotogastruridae) from Brazil. J. Insect Biodivers. 2014, 2, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Arlé, R. Conspecto das espécies brasileiras de Pseudachorutinae, com descrição de uma espécie nova da Colômbia (Insecta, Collembola). Acta Amaz. 1981, 11, 583–593. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, L.H.; Mendonça, M.C. Duas novas espécies de Pseudachorutinae (Collembola, Neanuridae) do Brasil. Bol. Mus. Nac. 2002, 496, 1–8. [Google Scholar]
- Da Paz, R.V.; Queiroz, G.C.; Bellini, B.C. A new species of Aethiopella Handschin, 1942 (Collembola, Poduromorpha, Neanuridae) from Neotropical Region, with comments on the genus. Zootaxa 2019, 4629, 39–50. [Google Scholar] [CrossRef]
- Arlé, R. Generalidades e importância ecológica da ordem Collembola (Apterygota). At. Soc. Biol. Rio J. 1959, 3, 4–7. [Google Scholar]
- Schuster, R. Über die Ökologie und Artengliederung der thalassobionten Collembolenfauna Brasiliens1. Beitr. Neotropischen Fauna 1965, 4, 191–208. [Google Scholar] [CrossRef]
- Arlé, R. Un nouveau collembole marin du littoral brésilien. Rev. Bras. Biol. 1966, 26, 367–372. [Google Scholar]
- Zeppelini, D.; Bellini, B.C.; Creão-Duarte, A.J.; Hernández, M.I.M. Collembola as bioindicators of restoration in mined sand dunes of Northeastern Brazil. Biodivers. Conserv. 2008, 18, 1161–1170. [Google Scholar] [CrossRef]
- Arlé, R. Collemboles d’Amazonie, I. Poduromorphes nouveaux ou peu connus et notes biologiques sur Neotropiella carli (Denis). Bol. Mus. Para. Emilio Goeldi 1966, 60, 1–19. [Google Scholar]
- Queiroz, G.; Deharveng, L. New Australonura Cassagnau 1980 (Neanuridae: Neanurinae) from Brazil. J. Nat. Hist. 2014, 48, 2293–2305. [Google Scholar] [CrossRef]
- Arlé, R. Novas espécies de Pseudachorutini (Collembola) do Rio de Janeiro e arredores. Bol. Biol. 1939, 4, 67–72. [Google Scholar]
- Zeppelini, D.; Queiroz, G.C.; Lopes, N.P.; Mendonça-Junior, F.J.B. Chemical analysis of Brasilimeria Stach, 1949 (Hexapoda, Collembola, Neanuridae) hemolymphatic secretion, and description of a new species. PLoS ONE 2019, 14, e0212451. [Google Scholar] [CrossRef] [Green Version]
- Arlé, R.; Rufino, E. Contribução ao conhecimento dos Pseudachorutinae da Amazônia (Collembola). Acta Amaz. 1976, 6, 99–107. [Google Scholar] [CrossRef] [Green Version]
- Arlé, R. Deux nouveaux collemboles du Brésil. Rev. Bras. Biol. 1943, 3, 109–112. [Google Scholar]
- Queiroz, G.C.; Deharveng, L. New genus, new species and new record of Neanurinae (Collembola, Neanuridae) for the Neotropics. Zootaxa 2015, 4020, 134–152. [Google Scholar] [CrossRef] [Green Version]
- Neves, A.C.; Mendonça, M.C.; Queiroz, G.C. Two new species and new records of Neanuridae (Hexapoda: Collembola) from Brazilian central Amazonia. Zoologi 2019, 36, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Queiroz, G.C.; de Mendonça, M.C. Two new Brazilian species of Friesea Dalla Torre, 1895 (Collembola: Neanuridae). Austral. Entomol. 2014, 54, 110–116. [Google Scholar] [CrossRef]
- Silveira, T.C.; Mendonça, M.C. Two new species of Neanuridae (Collembola: Poduromorpha) from littoral of Rio de Janeiro, Brazil. Neotrop. Entomol. 2018, 47, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, G.C.; Fernandes, L.H. New Brazilian species of Furculanurida Massoud, 1967 (Collembola: Neanuridae). Zootaxa 2011, 2805, 57–64. [Google Scholar] [CrossRef]
- Queiroz, G.C.; Mendonça, M.C. Revalidation of Handschinia Stach, 1949 (Collembola, Neanuridae) with description of a new species from Brazil. Soil Org. 2018, 86, 169–173. [Google Scholar]
- Zeppelini, D.; Palacios-Vargas, J.G. A New Brazilian Species of Hylaeanura(Collembola: Neanuridae). Fla. Entomol. 2013, 96, 1401–1405. [Google Scholar] [CrossRef]
- Arlé, R. Notas sôbre a família Oncopoduridae, com descrição de duas espécies novas do Brasil (Collembola). Arq. Mus. Nac. 1960, 50, 9–23. [Google Scholar]
- Oliveira, E.P.; Deharveng, L. Response of soil Collembola (Insecta) communities to forest disturbance in central Amazonia (Brazil). In Functioning and Dynamics of Natural and Perturbed Ecosystems; Bellan, D., Bonin, G., Emig, C., Eds.; Technique et Documentation, Lavoisier, Intercept Ltd.: Paris, France, 1995; pp. 361–376. [Google Scholar]
- Queiroz, G.C.; Silveira, T.C.; Mendonça, M.C. New species of Neotropiella Handschin, 1942 (Collembola: Neanuridae) from Brazil. Soil Org. 2013, 85, 41–49. [Google Scholar]
- Thibaud, J.M.; Oliveira, E.P. Note sur les Collemboles de L’Amazonie Brezilienne II—Neanuridae: Pseudachorutinae ad. p. avec la description de deux espèces nouvelles. Rev. Franç. d’Entom. 2011, 32, 135–140. [Google Scholar]
- Cassagnau, P.; Oliveira, E.P. Les collemboles Neanurinae d’Amérique du Sud. Bull. Soc. d’Hist. Natur. Toulouse 1990, 126, 19–23. [Google Scholar]
- Oliveira, E.P.; Deharveng, L. Deux nouvelles especes de Pseudachorutes d’Amazonie (Collemboles, Neanuridae). Bull. Soc. Entom. Fr. 1994, 99, 389–395. [Google Scholar]
- Börner, C. Voläufige Mitteilung über einige neue Aphorurinen und zur Systematik der Collembola. Zool. Anz. 1901, 633, 1–15. [Google Scholar]
- Mendonça, M.C.; Silveira, T.C. A new species of Tijucameria from Brazil (Collembola: Neanuridae: Pseudachorutinae). Soil Org. 2012, 84, 563–568. [Google Scholar]
- Mendonça, M.C.; Fernandes, L.H. Novo gênero e nova espécie de Pseudachorutini do sudeste do Brasil (Collembola, Neanuridae, Pseudachorutinae). Rev. Bras. Zool. 2005, 22, 699–701. [Google Scholar] [CrossRef] [Green Version]
- Arlé, R. Collemboles d’Amazonie, II. Pseudostachia folsomi gen. n. sp. n., intéressante espèce euédaphique. Bol. Mus. Para. Emilio Goeldi 1968, 68, 1–7. [Google Scholar]
- Oliveira, E.; Thibaud, J.-M. Notes sur les Collemboles de l’Amazonie, Brésil. 1. Hypogastruridae et Onychiuridae, avec la desciption de deux espèces nouvelles (Collembola). Opu. Zool. Flum. 1992, 95, 1–8. [Google Scholar]
- Rufino, E.O.; Schubart, H.O.R. Uma experiência sobre a nutrição de Onychiurus cunhai Arlé, 1970 (Insecta, Collembola), com notas sobre sua ecologia. Acta Amaz. 1974, 4, 53–55. [Google Scholar] [CrossRef] [Green Version]
- Palacios-Vargas, J.G.; Díaz, M. Survey of the Onychiuridae (Collembola) from the Neotropical region. Ana. Inst. Biol. Univ. Nac. Aut. Mexico 1996, 66, 165–180. [Google Scholar]


























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation—State | Species | Region | Abbreviation—Oceanic Island | Species | Region |
|---|---|---|---|---|---|
| AC—Acre | 0 | N | FN—Fernando de Noronha | 7 | EA |
| AL—Alagoas | 0 | NE | AR—Atol das Rocas | 2 | EA |
| AM—Amazonas | 24 | N | SPSP—São Pedro e São Paulo | 1 | EA |
| AP—Amapá | 2 | N | TR—Trindade e Martim Vaz | 0 | SA |
| BA—Bahia | 1 | NE |  | ||
| CE—Ceará | 1 | NE | |||
| ES—Espiro Santo | 17 | SE | |||
| GO—Goiás | 0 | CW | |||
| MA—Maranhão | 0 | NE | |||
| MG—Minas Gerais | 17 | SE | |||
| MS—Mato Grosso do Sul | 1 | CW | |||
| MT—Mato Grosso | 5 | CW | |||
| PA—Pará | 11 | N | |||
| PB—Paraíba | 4 | NE | |||
| PE—Pernambuco | 6 | NE | |||
| PI—Piauí | 2 | NE | |||
| PR—Paraná | 3 | SE | |||
| RJ—Rio de Janeiro | 80 | SE | |||
| RN—Rio Grande do Norte | 5 | NE | |||
| RO—Rondônia | 1 | N | |||
| RR—Roraima | 0 | N | |||
| RS—Rio Grande do Sul | 0 | S | |||
| SC—Santa Catarina | 0 | S | |||
| SE—Sergipe | 4 | NE | |||
| SP—São Paulo | 9 | SE | |||
| TO—Tocantins | 0 | N | |||
| Character | F. noronhaensis | F. rochedoensis | W. insularum | Acherontiella. sp. | P. zeliae |
|---|---|---|---|---|---|
| Body | 528.57 | 1106.69 | 520.64 | 746.69 | 710.50 |
| Cd | 135.52 | 234.98 | 78.58 | 130.36 | 149.12 |
| Ant I | 14.82 | 33.42 | 8.93 | 12.31 | 14.51 |
| Ant II | 16.23 | 30.08 | 14.18 | 16.47 | 18.75 |
| Ant III | 11.97 | 19.75 | 16.38 | 16.38 | 26,85 |
| Ant IV | 19.03 | 23.07 | 11.90 | 31.92 | 32.63 |
| Mucro | 4.52 | 6.70 | Absent | Absent | 27.41 |
| Dens | 10.41 | 23.49 | Absent | Absent | 28.02 |
| Manubrium | 22.96 | 23.87 | Absent | Absent | 36.28 |
| Unguis III | 12.50 | 20.28 | 10.57 | 13.35 | 18.27 |
| Tita III | 22.26 | 34.61 | 10.91 | 18.55 | 28.40 |
| Femur III | 19.82 | 32.36 | 17.73 | 18.09 | 15.82 |
| Tr | 15.55 | 24.61 | 9.08 | 15.50 | 10.32 |
| Cx | 32.53 | 36.09 | 12.63 | 23.92 | 17.34 |
| Th II ordinary chaetae | 4.98 | 13.16 | 4.19 | 7.89 | 8.02 |
| Th II s-chaetae | 11.83 | 16.17 | 7.98 | 13.53 | 14.22 |
| Anal spine | 12.78 | 20.66 | 6.97 | Absent | Absent |
| Abd V chaetae a1 or ord. | 5.25 | 19.56 | 9.65 | 8.02 | 8.21 |
| Abd V chaetae p1 or Mac. | 13.09 | 23.10 | 9.98 | 8.13 | 15.03 |
| Abd V s-chaetae | 15.08 | 20.35 | 21.70 | 17.08 | 16.31 |
| Species | Eyes | S Abd VI | Furca | M Abd VI | Ten h | BD | AP | Th I | Tita I-III | TL | Color |
|---|---|---|---|---|---|---|---|---|---|---|---|
| F. arlei Massoud and Bellinger, 1963 | 4 + 4 | Absent | 2 | Clavate | 1 clavate | ? | simple | 2 + 2 | ? | Jamaica | Blue |
| F. boitata Queiroz and Mendonça, 2015 | 8 + 8 | Absent | 5 | Clavate | 4,5,5 clavete | 400–800 | simple | 4 + 4 | 18,18,17 | Brazil | Blue |
| F. claviseta Axelson, 1900 | 8 + 8 | 3 | 2 | Clavate | ? | 550–700 | Finland | ? | |||
| F. cubensis Potapov and Banasko, 1985 | 8 + 8 | 3 | 1 | Acuminate | Acuminate | 670–1000 | simple | 4 + 4 | ? | Cuba | ? |
| F. curupira Queiroz and Mendonça, 2015 | 1 + 1 | 2 | 5 | Acuminate | Acuminate | 450–470 | simple | 4 + 4 | 15,15,14 | Brazil | Dark blue |
| F. josei Palacios-Vargas, 1986 | 8 + 8 | Absent | 5 | Spinne | 3,3,3 clavate | 1100 | simple | 3 + 3 | 17,17,16 | Porto Rico | Blue |
| F. jurubatiba Silveira and Mendonça, 2018 | 8 + 8 | Absent | 5 | Clavate | 4,5,5 clavate | 540–680 | simple | 4 + 4 | 18,18,17 | Brazil | Dark gray |
| F. magnicornis Denis, 1931 | 8 + 8 | 3 * | 2 | Clavate | 5 clavate | 900 | ? | ? | ? | Costa Rica | Gray |
| F. mirabilis (Tullberg, 1871) | 8 + 8 | 3 | 2 | Acuminate | ? | ? | ? | ? | ? | ? | ? |
| F. noronhaensis sp. nov. | 8 + 8 | 3 | 1 | Clavate | 3,4,4 clavate | 440–530 | simple | 4 + 4 | 18,18,17 | Brazil | Dark blue |
| F. reducta Denis, 1931 | 8 + 8 | Absent | 5 | Clavate | 4,5,5 clavete | 600 | simple | 3 + 3 | ? | Costa Rica | Gray |
| F. rochedoensis sp. nov. | 8 + 8 | 3 | 1 | Acuminate | 3,4,4 clavate | 890–1180 | simple | 4 + 4 | 18,18,17 | Brazil | Light blue |
| F. sublimis Macnamara, 1921 | 8 + 8 | 3 | 2 | Clavate | ? | ? | ? | ? | ? | ? | ? |
| Species | Eyes | BD | AP | Th I | Tita I-III | Color | SF | MT | TL |
|---|---|---|---|---|---|---|---|---|---|
| A. albipes (Folsom, 1927) | 5 + 5 | 0.75–1.5 | 3 | 2 + 2 | 19,19,18 | Dark blue | 022/11111 | 4 | Panama |
| A. arleana Mendonça and Fernandes, 1999 | 5 + 5 | 1.88 | 3 | 3 + 3 | 19,19,18 | Black and yellow | 022/11111 | 6 | Brazil |
| A. intermedia Fernandes and Mendonça, 2004 | 5 + 5 | 0.98–1.6 | 3 | 3 + 3 | 19,19,18 | Light gray | 022/11111 | 5 | Brazil |
| Species | Eyes | AP | PAO | Mx T | Th I | Ten h | TT | Tita I–III | DC | TL |
|---|---|---|---|---|---|---|---|---|---|---|
| B. septemoculata Denis, 1931 | 7 + 7 | 1 | 5 | 8 | 2 + 2 | acuminate | 3 | 19,19,18 | 5 | Costa Rica |
| B. garayae Queiroz and Weiner, 2011 | 2 + 2 | 3 | 5–6 | 8–9 | 2 + 2 | acuminate | 3 | 19,19,18 | 5 | Brazil |
| B. platensis Najt and Massoud, 1974 | 8 + 8 | 2–3 | 5 | 7 | 2 + 2 | capitate | 3 | 19,19,18 | 5 | Argentina |
| B. parvula (Schäffer, 1896) | 8 + 8 | 3 | 6–8 | 7 | 3 + 3 | capitate | ? | 19,19,18 | 5 | Germany |
| B. agrosa Wray, 1953 | 8 + 8 | 1 | 4 | 7–10 | 2 + 2 | acuminate | 3 | 19,19,18 | 5 | Puerto Rico |
| B. ceciliae Fernandes and Mendonça, 2004 | 8 + 8 | 1–2 | 4 | 8 | 2 + 2 | capitate | 3 | 18,18,17 | 6 | Brazil |
| B. nordestina Souza, Bellini and Weiner, 2018 | 8 + 8 | 3 | 4 | 6–8 | 2 + 2 | acuminated | 3 | 19,19,18 | 5 | Brazil |
| B. villalobosi Casagnau and Rapoport, 1962 | 8 + 8 | 3 | 3–4 | 6 | 3 + 3 | capitate | 3 | 18,18,17 | 3 | Brazil |
| B. saladaensis Weiner and Najt, 2001 | 8 + 8 | 3 | 6 | 7 | 2 + 2 | acuminate | 3 | 19,19,18 | 5 | Argentina |
| B. contorta Denis, 1931 | 8 + 8 | 1 | 5 | 7 | 2 + 2 | acuminate | 3 | 19,19,18 | 5 | Costa Rica |
| Species | S VI | AL | Ten h | BD | NS | Th I | Tita I–III | Femur | Tr | TL |
|---|---|---|---|---|---|---|---|---|---|---|
| A. globulata Thibaud e Massoud, 1980 | Absent | Smooth | 1 | 520 | 6 | 3 + 3 | ? | ? | ? | Guadalupe |
| A. colotiplana Palacios-Vargas and Thibaud, 1985 | Absent | Smooth | 1 | 450 | 5 | 3 + 3 | 17,17,16 | 12,11,10 | 3,4,4 | Mexico |
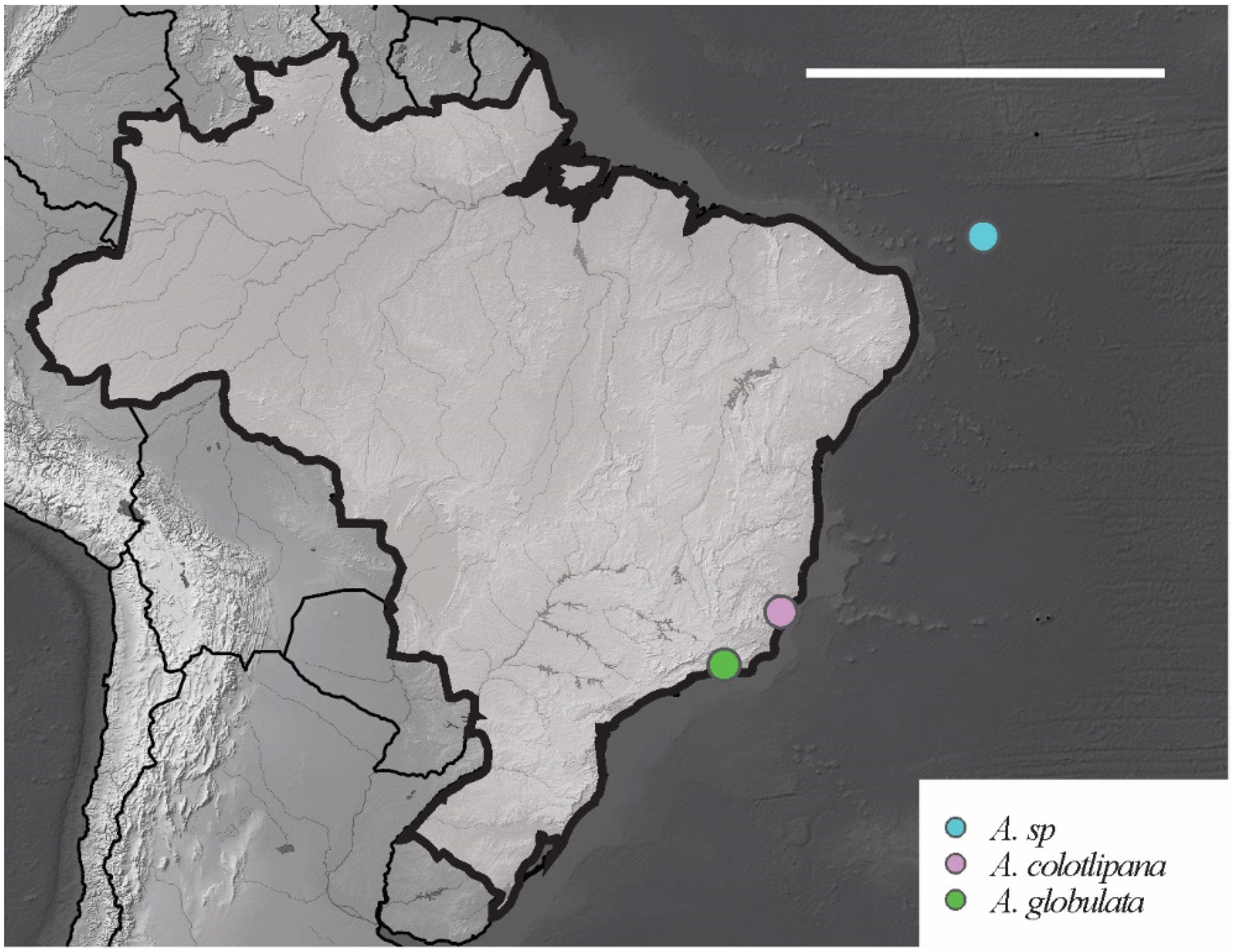
| A. sp. Lima and Zeppelini, 2015 | Absent | Spine | 1 | 746 | 5 | 3 + 3 | 18,18,17 | 11,11,10 | 3,4,4 | Brazil |
| Species | VT | TT | VC | Ten h | BD | Th I | Tita I-III | Femur | Trochanter | TL |
|---|---|---|---|---|---|---|---|---|---|---|
| Paraxenylla piloua Thibaud and Weiner, 1997 | 1 + 1 | 3 | 1 + 1 | 4 Ac | 360–400 | 3 + 3 | 19,19,18 | 12,12,10 | 5,5,5 | New Caledonia |
| Paraxenylla sooretamensis Queiroz and Deharveng, 2008 | 4 + 4 | 3 | ? | 0 | 440–820 | 3 + 3 | 19,19,18 | 12,12,10 | 5,5,4 | Brazil |
| Paraxenylla zeliae sp. nov. | 1 + 1 | 3 | 1 + 1 | 4 Cl | 490–720 | 3 + 3 or 4 + 4 | 19,19,18 | 12,12,11 | 5,5,5 | Brazil |
| Species | 00000 00,000 11111 11,111 22222 22,222 33333 33,333 44444 44,444 55 |
|---|---|
| 01234 56,789 01234 56,789 01234 56,789 01234 56,789 01234 56,789 01 | |
| W. brevispina Hüther, 1962 | 13032 03,111 11311 11,101 00122 10,100 01101 11,111 11111 11,111 01 |
| W. zeppelini D’Haese and Thibaud, 2011 | 0c0b2 2b111 01,211 1110? 11,122 10000 01,101 11110 11,001 01111 11 |
| W. insularum sp. nov. | 13032 03,111 11?11 10,101 00033 11,110 11111 11,111 11011 11,100 01 |
| Species | Eyes | Ant IV Sensilla | Furca | Manubrial Chaeta | OCM | Ten h | TL |
|---|---|---|---|---|---|---|---|
| X. maritima Tullberg, 1869 | 5 + 5 | 4 | MD | ? | Smooth | 2,2,2 | ? |
| X. nirae Gama and Oliveira, 1994 | 5 + 5 | 6 | MSD | ? | ? | 1,1,1 | Brazil |
| X. brasiliensis da Gama, 1978 | 5 + 5 | 4 | MD | ? | Smooth | 1,1,1 | Brazil |
| X. subcavernarum Gama, 1969 | 5 + 5 | 4 | MD | ? | ? | 2,2,2 | Brazil |
| X. welchi Folsom, 1916 | 5 + 5 | 4 | MSD | 34 | Smooth | 1,2,2 | North America |
| X. hodori Neves and Mendonça, 2017 | 5 + 5 | 4 | MSD | 28 | Serrated | 2,2,2 | Brazil |
| X. yucatana Mills, 1938 | 4 + 4 | 4 | MSD | 30 | Smooth | 2,2,2 | Mexican |
| X. manuelae Queiroz and Mendonça, 2016 | 5 + 5 | 4 | MSD | 34 | Serrated | 2,2,2 | Brazil |
| X. capixaba Fernandes and Mendonça, 2010 | 5 + 5 | 4–5 | MSD | 34 | Serrated | 2,2,2 | Brazil |
| X. wandae Queiroz and Mendonça, 2016 | 5 + 5 | 6 | MSD | 34 | Serrated | 1,2,2 | Brazil |
| µm | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I. mucrospatulata | 350 | + | 1 | — | — | BI | 4 | SP | MOD | 7 + 1 | — | 4 | 16 | 5/2 | 5 + 5 |
| I. praiana | 450 | + | ? | + | + | BI | 4 | H | un | 6 | — | 5 | 13 | 5/2 | ? |
| Family/Species | Brazil Distribution | World Distribution | Habitats | Record |
|---|---|---|---|---|
| Brachystomellidae Stach, 1949 | ||||
| Brachystomella agrosa Wray, 1953 | FN, PE, BA, PE, RJ, SP | Neo | Halophyte–psammophyte vegetation, sand dunes, flooded areas, pasture, eucalyptus, citric plantation, understory, riparian vegetation, moss, litter and soil over beach rocks, oceanic island | [3,20,35] * |
| Brachystomella aspera (Börner, 1906) | São Francisco | NCB? | un | [36] |
| Brachystomella ceciliae Fernandes and Mendonça, 2004 | RJ (ES) | NCB | Halophyte–psammophyte vegetation, litter on sand dunes, flooded areas | [37] |
| Brachystomella contorta Denis, 1931 | RJ, (ES) | Bor, Neo, Pal | Halophyte–psammophyte vegetation, sand dunes, beach sand, moss, soil over beach rocks | [37,38] |
| Brachystomella garayae Queiroz and Weiner, 2011 | ES | NCB | Forest litter | [39] |
| Brachystomella nordestina Souza, Bellini and Weiner, 2018 | RN | NCB | Leaf litter over sandy soil of an urban forested | [40] * |
| Brachystomella parvula (Schäffer, 1896) | RN | Cos | Forest litter | [41] |
| Brachystomella platensis Najt and Massoud, 1974 | ES, RJ | Neo, Aus | Beach sand, litter over sandy dunes, halophyte–psammophyte vegetation, guriri vegetation, dunes and supralittoral vegetation | [42,43] * |
| Brachystomella saladaensis Weiner and Najt, 2001 | CE | Neo | Leaf litter on the shore of lakes | [44] |
| Brachystomella septemoculata Denis, 1931 | RJ | Pal, Neo | Dunes, rotten wood | [45] |
| Brachystomella villalobosi Casagnau and Rapoport, 1962 | PE | Neo | un | [35] |
| Brachystomellides compositus Arlé, 1959 | RJ | Neo | Beach sand, litter | [46] |
| Folsomiella albida (Arlé, 1959) | RJ | Neo | Leaf litter | [38,46] |
| Folsomiella ceca (Folsom, 1927) | (RJ) | Neo | Soil | [47] |
| Folsomiella intermedia (Arlé, 1939) | RJ | NCB | Soil | [38,48] |
| Folsomiella pseudoceca Mendonça, Fernandes and Abrantes, 2005 | RJ | NCB | Soil | [49] |
| Folsomiella trisetosa Mendonça, Fernandes and Abrantes, 2005 | RJ, SP | NCB | Soil and rotten logs, soil near streams | [49] |
| Maricaella duna Mendonça and Fernandes, 1997 | RJ | NCB | Leaf litter, sand dunes | [50] |
| Micronella itacaman Queiroz and Mendonça, 2013 | RJ, MG | NCB | Leaf litter and soil, altitude 2400 m | [51] * |
| Micronella longisensilla Queiroz and Mendonça, 2013 | RJ | NCB | Leaf litter and soil, altitude 2100 m | [51] * |
| Micronella porcus (Denis, 1933) | RJ | Neo | Soil | [46] |
| Neorganella rotundatae Queiroz and Mendonça, 2013 | RJ, MG | NCB | Leaf litter and soil, altitude 2400 m | [51] * |
| Rapoportella pitomboi Mendonça and Fernandes, 1995 | MG, RJ | NCB | Halophyte–psammophyte vegetation, sand dunes, flooded areas | [37,52] |
| Setanodosa occidentalis (Arlé, 1959) | RJ | NCB | un | [38,46] |
| Hypogastruridae Börner, 1906 | ||||
| Acherontides eleonorae Palacios-Vargas and Gnaspini-Netto, 1992 | PR, SP | NCB | Guano piles of hematophagous bats in caves | [53] |
| Acherontides serrasapoensis Lima, Stievano and Zeppelini, 2019 | MG | NCB | Superficial subterranean habitats and cave | [54] * |
| Acherontiella colotipana Palacios-Vargas and Thibaud, 1985 | ES | Pal, Neo | Soil | [55] |
| Acherontiella globulata Thibaud and Massoud, 1980 | RJ | Neo | Halophyte–psammophyte vegetation, sand dunes | [37] |
| Austrogastrura marambaia Fernandes, Bellini and Mendonça, 2010 | RJ | NCB | Halophyte–psammophyte vegetation | [56] |
| Austrogastrura travassosi (Arlé, 1939) | MS, RJ, PB | Neo | Halophyte–psammophyte vegetation, litter, over rocks and littoral sand | [56,57,58,59] |
| Ceratophysella armata (Nicolet, 1842) | (RJ) | Bor, Cos?, Neo | un | [60] |
| Ceratophysella bengtssoni (Ågren, 1904) | RJ | Bor, NCB | Littoral sand | [59] |
| Ceratophysellaengadinensis (Gisin 1949) | PR, RJ | Neo, Pal | Leaf litter and soil of “campos de altitude” | [61] * |
| Ceratophysella nataliae Cipola, Bellini and Palacios-Vargas, 2017 | PR | NCB | Soil, semi-arid vegetation | [62] * |
| Ceratophysella rogerarlei Palacios- Vargas, Bellini and Cipola, 2017 | PI | NCB | Soil, anthropized forest from Atlantic forest | [62] * |
| Hypogastrura manubrialis (Tullberg, 1869) | RJ | Cos | un | [63] |
| Hypogastrura rehi Börner, 1906 | (SP) | NCB | un | [36] |
| Mesogastrura ojcoviensis (Stach, 1919) | RJ | Bor, Neo | Littoral sand | [59] |
| Paraxenylla piloua Thibaud and Weiner, 1997 | RJ | NCB | Halophyte–psammophyte vegetation | [64] |
| Paraxenylla sooretamensis Queiroz and Deharveng, 2008 | ES | NCB | Forest litter | [65] |
| Paraxenylla zelliae sp.nov. | AR | NCB | Oceanic islands | sp. nov.* |
| Schoetella celiae Fernandes and Mendonça, 1998 | SP | NCB | Forest litter | [66] |
| Willemgastrura coeca Oliveira and Thibaud, 1988 | AM, RO | Amz, NCB | Soil | [67] |
| Willemia brevispina Hüther, 1962 | RJ, ES | Ant, Neo, Pal | Soil and littoral sand | [55,59] |
| Willemia insularum sp. nov | FN, AR | NCB | Oceanic islands | sp. nov.* |
| Willemia zeppelinii D’Hese and Thibaud, 2011 | PB | NCB | Coastal interstitial psammophilous | [23] |
| Xenylla brasiliensis da Gama, 1978 | MG, RJ | NCB | Leaf litter, forest soil, foredunes zone, halophyte‒psammophyte vegetation, dunes vegetation and supralittoral vegetation | [43,68] * |
| Xenylla capixaba Fernandes and Mendonça, 2010 | ES | NCB | Leaf litter, sand dunes | [42] |
| Xenylla hodori Neves and Mendonça, 2017 | AM | Amz | Leaf litter and soil | [69] * |
| Xenylla manuelae Queiroz and Mendonça, 2016 | MG, ES, RJ | NCB | Soil leaf litter, altitude 2400 m | [61] * |
| Xenylla maritima Tullberg, 1869 | RJ | Cos | Halophyte–psammophyte vegetation, sand dunes, flooded areas | [37] |
| Xenylla nirae Gama and Oliveira, 1994 | AM | Amz | Soil | [70] |
| Xenylla wandae Queiroz and Mendonça, 2016 | RJ | NCB | Soil and leaf litter of altitude, 2100 m | [61]* |
| Xenylla welchi Folsom, 1916 | ES, RJ | Cos | Soil, halophyte–psammophytevegetation, foredunes zone | [55,64] |
| Xenylla yucatana Mills, 1938 | FN, ES, RJ | Pal, Neo, Aus | Beach sand, halophyte‒psammophyte vegetation, oceanic islands, dunes vegetation and supralittoral vegetation | [3,42,43] * |
| Isotogastruridae Thibaud and Najt, 1992 | ||||
| Isotogastrura mucrospatulata Palacios-Vargas, Lima and Zeppelini, 2013 | FN | NCB | Oceanic islands | [34] * |
| Isotogastrura praiana Silveira, Mendonça and Da-Silva, 2014 | RJ | NCB | halophyte‒psammophyte vegetation and dunes vegetation | [71] * |
| Neanuridae Börner, 1901 | ||||
| Aethiopella delamarei Arlé, 1960 | MG, RJ, RN | Neo | Leaf litter | [41,46,72] |
| Aethiopella littoralis Fernandes and Mendonça, 2002 Aethiopella ricardoi Paz, Bellini and Queiroz, 2019 | RJ RN | NCB NCB | Halophyte–psammophyte vegetation, sand dunes, flooded areas sandy soil surrounded by dead foliage near vegetation and lentic freshwater | [73,74] * |
| Anurida maritima (Guérin-Méneville, 1836) | Brazilian coast, PE, SP, RJ, ES, RN | Cos | Surface of rock-pools on the coast | [41,72,75,76,77] |
| Arlesia albipes (Folsom, 1927) | PA, PE, RJ, PB, PI, FN | Neo | Soil, leaf litter, sand dunes and forest, oceanic islands | [38,43,78] * |
| Arlesia arleana Mendonça and Fernandes, 1999 | PE | NCB | Leaf litter | [16] |
| Arlesia intermedia Fernandes and Mendonça, 2004 | RJ | NCB | Sand dunes, flooded areas | [37] |
| Arlesiella amazonica Arlé, 1966 | AM | Amz | Leaf litter | [72,79] |
| Australonura gili Queiroz and Deharveng, 2014 | RJ | NCB | Leaf litter and soil of “campos de altitude” | [80] * |
| Australanura neotropica Queiroz and Deharveng, 2014 | RJ | NCB | Leaf litter and soil of “campos de altitude” | [80] * |
| Brasilimeria anura Arlé, 1939) | RJ | NCB | Humus under dead leaf | [81] |
| Brasilimeria assu Queiroz and Zeppelini, 2019 | RJ | NCB | Leaf litter and soil of Brazilian Páramos | [82] * |
| Brasilimeria wygodzinskyi (Arlé, 1943) | RJ, MG | NCB | un | [83,84] |
| Cephalachorutes anneae Queiroz and Mendonça, 2016 | RJ | NCB | Soil leaf litter, altitude 2500 m | [61] * |
| Ectonura snowdeni Queiroz and Deharveng, 2015 | MG | NCB | Leaf litter and soil, altitude 2500–2800 m | [85] * |
| Friesea arlei Massoud and Bellinger, 1963 | MT | Neo | un | [79] |
| Friesea multiclavata Neves, Mendonça and Queiroz, 2019 | AM | NCB | Forest leaf litter of Amazon Rainforest, | [86] * |
| Friesea boitata Queiroz and Mendonça, 2015 | RJ | Neo | Soil and leaf litter 2582 m | [87] * |
| Friesea claviseta Axelson, 1900 | RJ | Cos | Halophyte–psammophyte vegetation | [64] |
| Friesea cubensis Potapov and Banasko, 1985 | RJ | Neo | Littoral sand | [59] |
| Friesea curupira Queiroz and Mendonça, 2015 | RJ | NCB | Soil and leaf litter 2500 m | [87] * |
| Friesea josei Palacios- Vargas, 1986 | RJ | Neo | Littoral sand | [59] |
| Friesea jurubatiba Silveira and Mendonça, 2018 | RJ | NCB | Beach sand and litter of “restinga” soil | [88] * |
| Friesea magnicornis Denis, 1931 | RJ | Neo | Halophyte–psammophyte vegetation | [64] |
| Friesea mirabilis (Tullberg, 1871) | RJ | Neo | Halophyte–psammophyte vegetation | [64] |
| Friesea noronhaensis sp. nov. | FN | NCB | Oceanic islands | sp. nov.* |
| Friesea reducta Denis, 1931 | RJ | Neo | Neo Halophyte–psammophytevegetation, sand dunes | [37,64] |
| Friesea rochedoensis sp.nov. | SPSP | NCB | Oceanic islands | sp. nov.* |
| Friesea sublimis Macnamara, 1921 | ES | Neo, Bor, Pal | Soil | [55] |
| Furculanurida belemensis Arlé and Rufino, 1976 | PA | Amz | Soil | [83] |
| Furculanurida boiunia Neves, Mendonça and Queiroz, 2019 | AM | Amz | Forest leaf litter of Amazon rainforest | [86] * |
| Furculanurida goeldiana Arlé and Rufino, 1976 | PA | Amz | Leaf litter | [83] |
| Furculanurida nessimiani Fernandes and Mendonça, 2002 | SP | NCB | Forest litter | [74]) |
| Furculanurida tropicalia Queiroz and Fernandes, 2011 | ES | NCB | Forest litter | [89] |
| Halachorutes schusteri Arlé, 1966 | PA, RJ | Amz, NCB | Leaf litter | [72,77] |
| Handschinurida fluminensis (Arlé, 1939) | RJ, SE | NCB | un | [38,81] |
| Handschinurida proxima (Arlé, 1939) | RJ, SE, SP | NCB | un | [16,38,81] |
| Handschinurida rauli Queiroz and Mendonça, 2014 | MG | NCB | Soil and leaf litter, altitude 2700 m | [90] * |
| Hylaeanura infima (Arlé, 1959) | MT, AM, PA, RJ | Neo | Soil, leaf litter, sand dunes, flooded areas | [37,38,46,79] |
| Hylaenura mendoncae Zeppelini and Palacios- Vargas, 2013 | MG | NCB | Areas of iron ore mining; leaf litter of semi-deciduous forest fragments | [91] |
| Itanura brasiliensis (Arlé, 1960) | RJ, MG | NCB | [92] | |
| Kenyura delicata Arlé, 1966 | AM | Amz | Leaf litter | [72,79] |
| Kenyura porcula (Arlé, 1959) | RJ | NCB | [46] | |
| Kenyura xinguensis Arlé, 1966 | MT | NCB | Leaf litter | [72,79] |
| Micranurida fluminensis Fernandes and Mendonça, 2004 | RJ | NCB | Sand dunes, flooded areas | [37]) |
| Neotropiella arlei Najt, Thibaud and Weiner, 1990 | AM | Neo | Primary and secondary forest soil and litter | [93] |
| Neotropiella barbatae Queiroz, Silveira and Mendonça, 2013 | RJ | NCB | Soil and leaf-litter of “campos de altitude” 2400m | [94] * |
| Neotropiella carli (Denis, 1924) | (AM, AP, PA) | Neo | Leaf litter | [38] |
| Neotropiella denisi (Arlé, 1939) | RJ, SE, MT | NCB | Soil, litter on roots | [38,72,81] |
| Neotropiella digitomucronata Thibaud and Massoud, 1983 | AM | Neo | Primary and secondary forest soil and litter | [93] |
| Neotropiella insularis Queiroz, Silveira and Mendonça, 2013 | RJ | NCB | Leaf litter of Atlantic rainforest | [94] * |
| Neotropiella macunaimae Queiroz, Silveira and Mendonça, 2013 | MG | NCB | Soil and leaf litter, altitude 2700 m | [94] * |
| Neotropiella meridionalis (Arlé, 1939) | SE, MT, RJ, MG, PA, AM | Neo | Leaf litter, rotten woods | [38,72,79,81,93] |
| Neotropiella minima Thibaud and Oliveira, 2011 | AM | Amz | Floodable rain forest, clay soil | [95] |
| Neotropiella plurichaetosa Thibaud and Oliveira, 2011 | AM | Amz | Floodable rain forest, clay soil | [95] |
| Neotropiella quinqueoculata (Denis, 1931) | RJ, AM, AP, MT, PA, RJ | Bor, Neo | un | [35,79] |
| Neotropiella silvestrii (Denis, 1929) | AM | Neo | [95] | |
| Neotropiella vanderdrifti Massoud, 1963 | AM | Neo | Primary and secondary forest soil and litter | [93] |
| Paleonura brasiliensis (Arlé, 1959) | MG (RJ) | NCB | Leaf litter | [46] |
| Paleonura nuda Cassagnau and Oliveira, 1990 | AM | Amz | Primary and secondary forest soil and litter | [96] |
| Pronura amazonica Cassagnau and Oliveira, 1990 | AM | Amz | Leaf litter | [96] |
| Pseudachorutes bifasciatus Oliveira and Deharveng, 1994 | AM | Amz | Soil | [97] |
| Pseudachorutes difficilis Denis, 1931 | RJ, MG, ES | Neo | Sand dunes, flooded areas, soil and leaf litter of highlands grasslands | [37,43] * |
| Pseudachorutes gilvus Oliveira and Deharveng, 1994 | AM | Amz | Soil | [97] |
| Pseudachorutes herberti Arlé and Rufino, 1976 | AM | NCB | Leaf litter | [83] |
| Pseudachorutes massoudi Arlé, 1966 | AM | Amz | Leaf litter | [72,79] |
| Pseudachorutes parvulus Börner,1901 | PB | Cos | un | [98] |
| Pseudachorutes solaris Silveira and Mendonça, 2018 | RJ | NCB | Sand beach and litter of “restinga” | [88] * |
| Pseudanurida sawayana Schuster, 1965 | PE, SP, (RJ) | Bor, Neo, Pal | On sand and seaweed on the rocky coast | [76] |
| Tijucameria gabrieli Mendonça and Silveira, 2012 | RJ | NCB | Leaf litter and soil between roots on montane vegetation of the Atlantic rainforest | [99] * |
| Tijucameria mame Mendonça and Fernandes, 2005 | RJ | NCB | Forest litter | [100] |
| Odontellidae Massoud, 1967 | ||||
| Stachia folsomi (Arlé, 1968) | PA, RJ | Amz, Pal | Soil, dunes vegetation | [43,101] * |
| Onychiuridae Lubbock, 1867 | ||||
| Agraphorura fernandae (Oliveira and Thibaud, 1992) | PA | Amz | Leaf litter | [102] |
| Agraphorura mariapetrae (Thibaud, 1993) | RJ | Neo | Flooded areas | [37] |
| Onychiurus cunhai Arlé, 1970 | PA, AM | Neo | Soil | [58,103] |
| Protaphorura cryptopyga (Denis, 1931) | RJ | Neo, Pal | un | [35] |
| Tullbergiidae Bagnall, 1935 | ||||
| Fissuraphorura cubanica Rusek, 1991 | RJ, ES | Neo | Soil, halophyte–psammophytevegetation | [59,63] |
| Mesaphorura amazonica Oliveira and Thibaud, 1992 | AM, RJ, ES | Amz, NCB | Litter, halophyte–psammophyte vegetation, sand dunes, soil | [37,55,102] |
| Mesaphorura iowensis (Mills, 1932) | SP | Cos | Soil and litter | [104] |
| Mesaphorura maricaensis Fernandes and Mendonça, 2004 | RJ | NCB | Sand dunes | [37] |
| Mesaphorura yosiii (Rusek, 1967) | AM, SP, RJ | Cos | Primary and secondary forest soil and litter; halophyte–psammophyte vegetation, sand dunes and flooded areas, soil, litter | [59,102,104] |
| Tullbergia minensis Arlé, 1959 | MG | NCB | Litter | [46] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Lima, E.C.A.; de Mendonça, M.C.; Queiroz, G.C.; da Silveira, T.C.; Zeppelini, D. Synthesis of the Brazilian Poduromorpha (Collembola: Hexapoda) with Special Emphasis on the Equatorial Oceanic Islands. Insects 2021, 12, 268. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12030268
de Lima ECA, de Mendonça MC, Queiroz GC, da Silveira TC, Zeppelini D. Synthesis of the Brazilian Poduromorpha (Collembola: Hexapoda) with Special Emphasis on the Equatorial Oceanic Islands. Insects. 2021; 12(3):268. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12030268
Chicago/Turabian Stylede Lima, Estevam C. A., Maria Cleide de Mendonça, Gabriel Costa Queiroz, Tatiana Cristina da Silveira, and Douglas Zeppelini. 2021. "Synthesis of the Brazilian Poduromorpha (Collembola: Hexapoda) with Special Emphasis on the Equatorial Oceanic Islands" Insects 12, no. 3: 268. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12030268