
Within-Season Changes in Land-Use Impact Pest Abundance in Smallholder African Cassava Production Systems

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
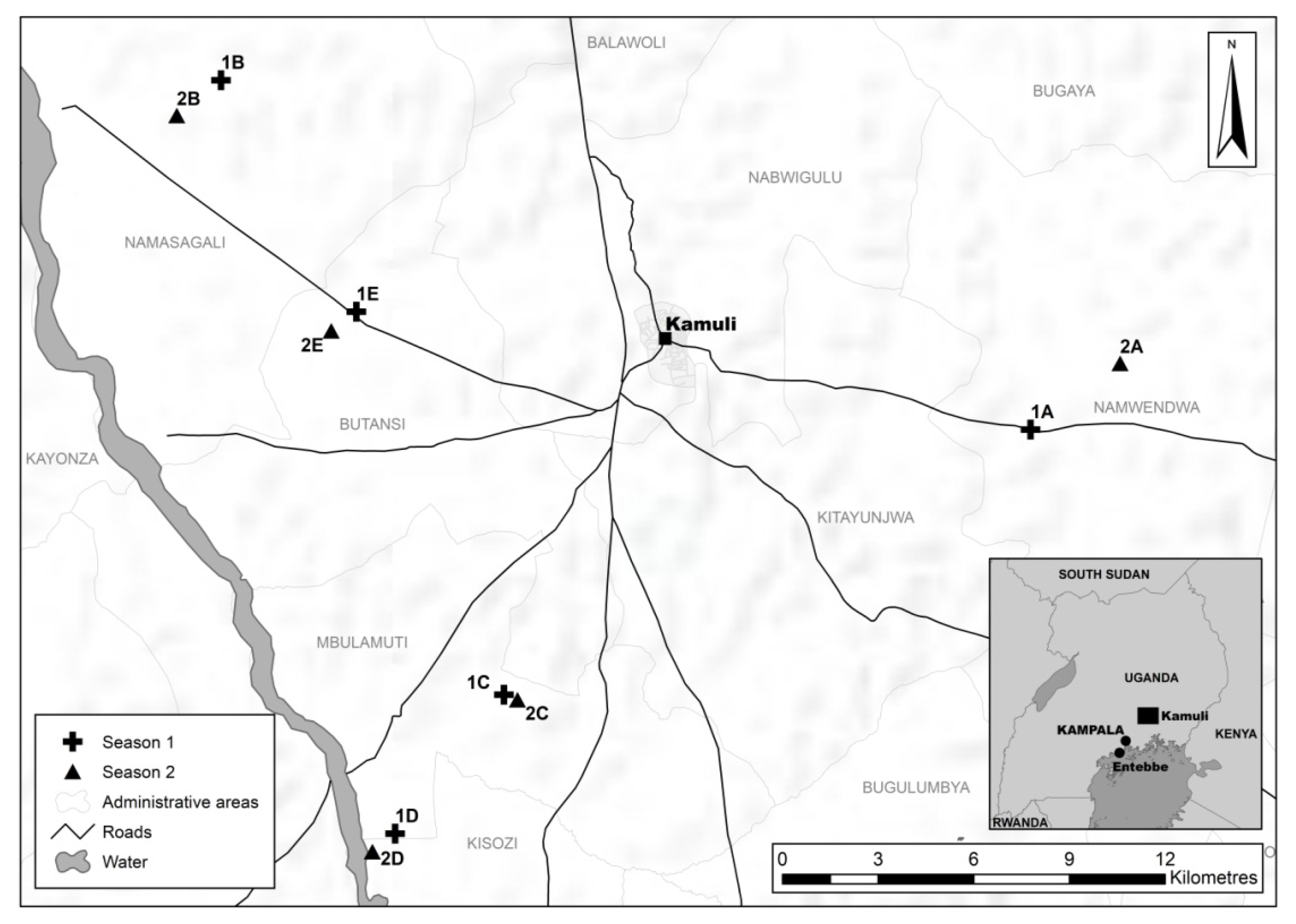
2.1. Study Area
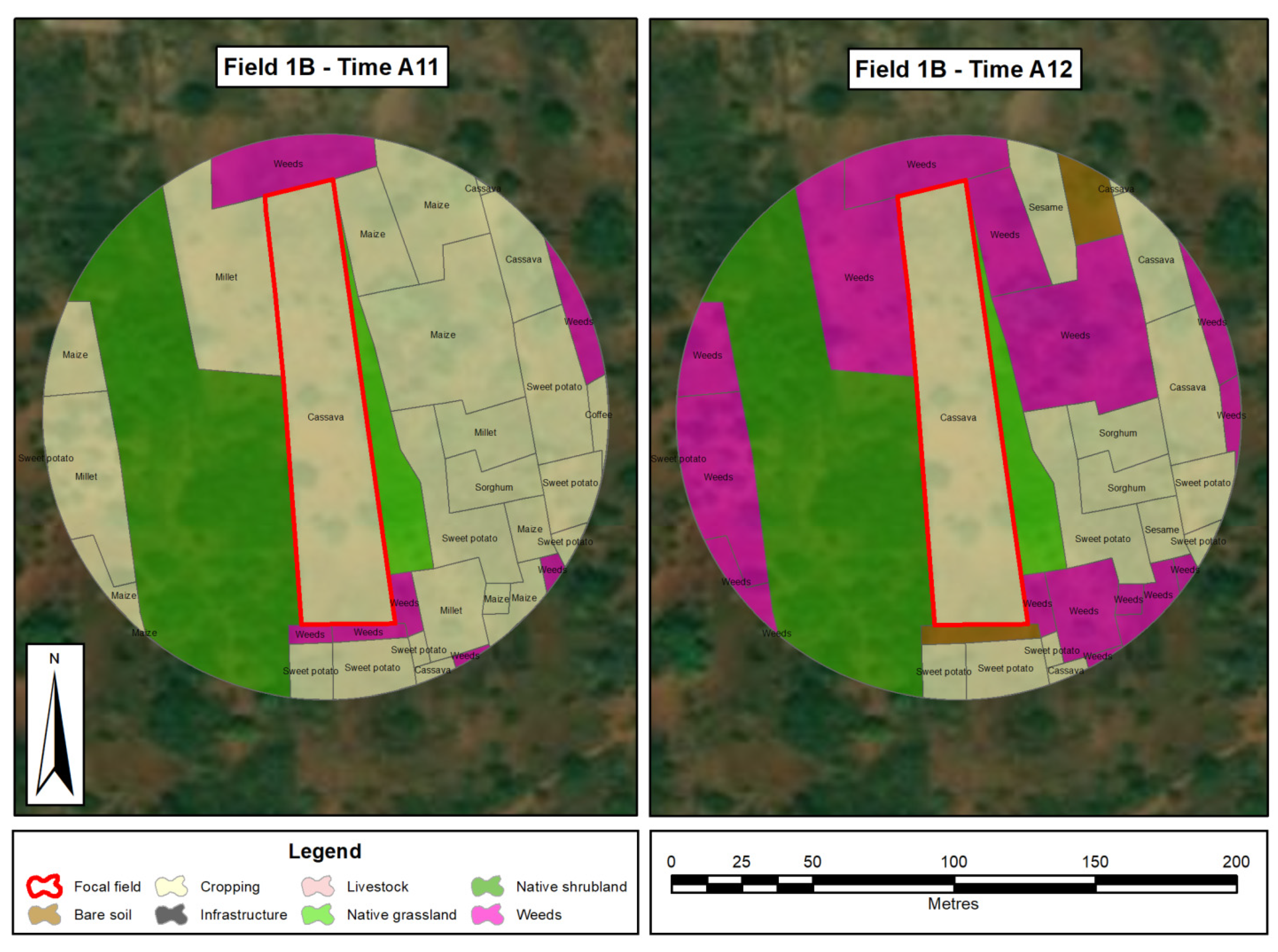
2.2. Planting of Focal Cassava Fields
2.3. Sampling Bemisia Species and Their Parasitoids
2.4. Molecular Characterization of Bemisia Species Complex
2.4.1. gDNA Extraction.
2.4.2. PCR and High Throughput Amplicon Sequencing
2.5. Data Analysis
2.5.1. Predictor and Explanatory Variables
2.5.2. Cluster Analysis
3. Results
3.1. Identity of the Bemisia Cryptic Species
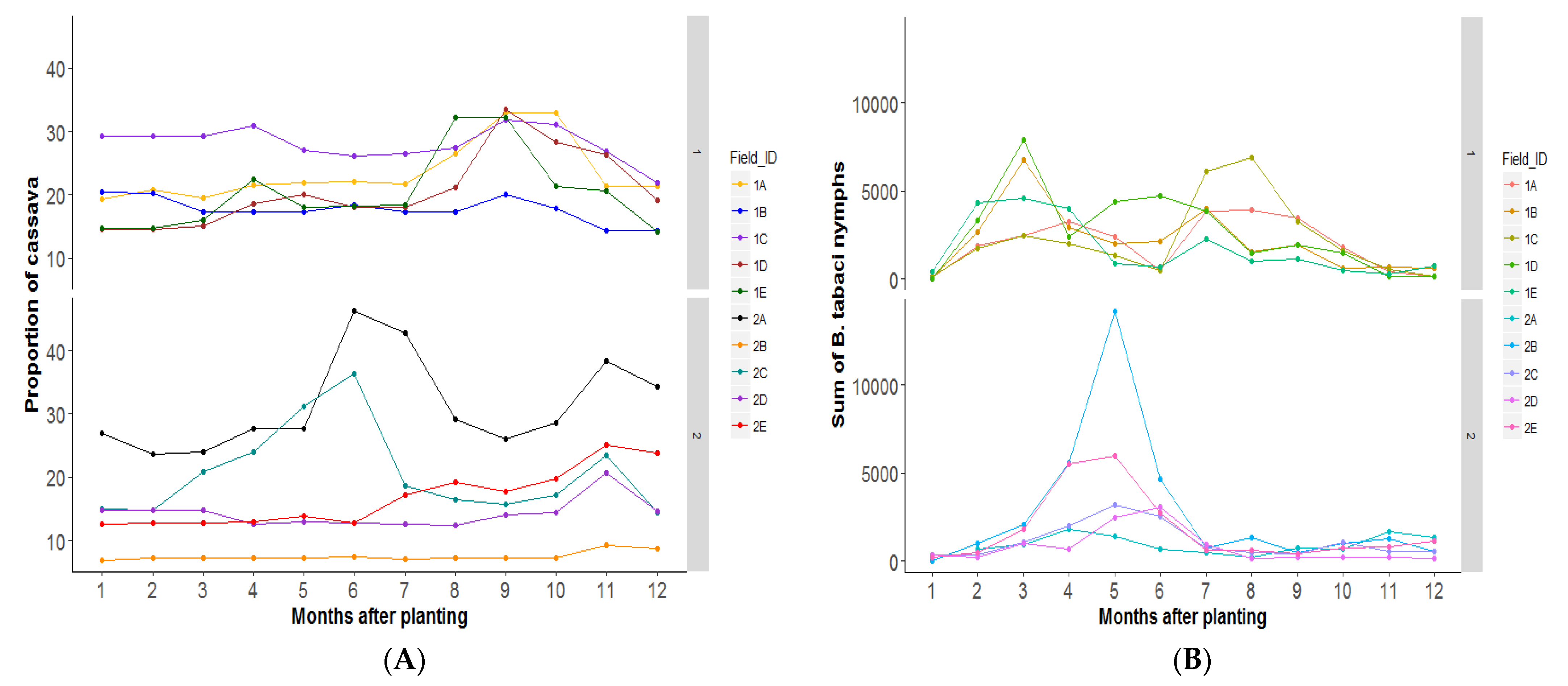
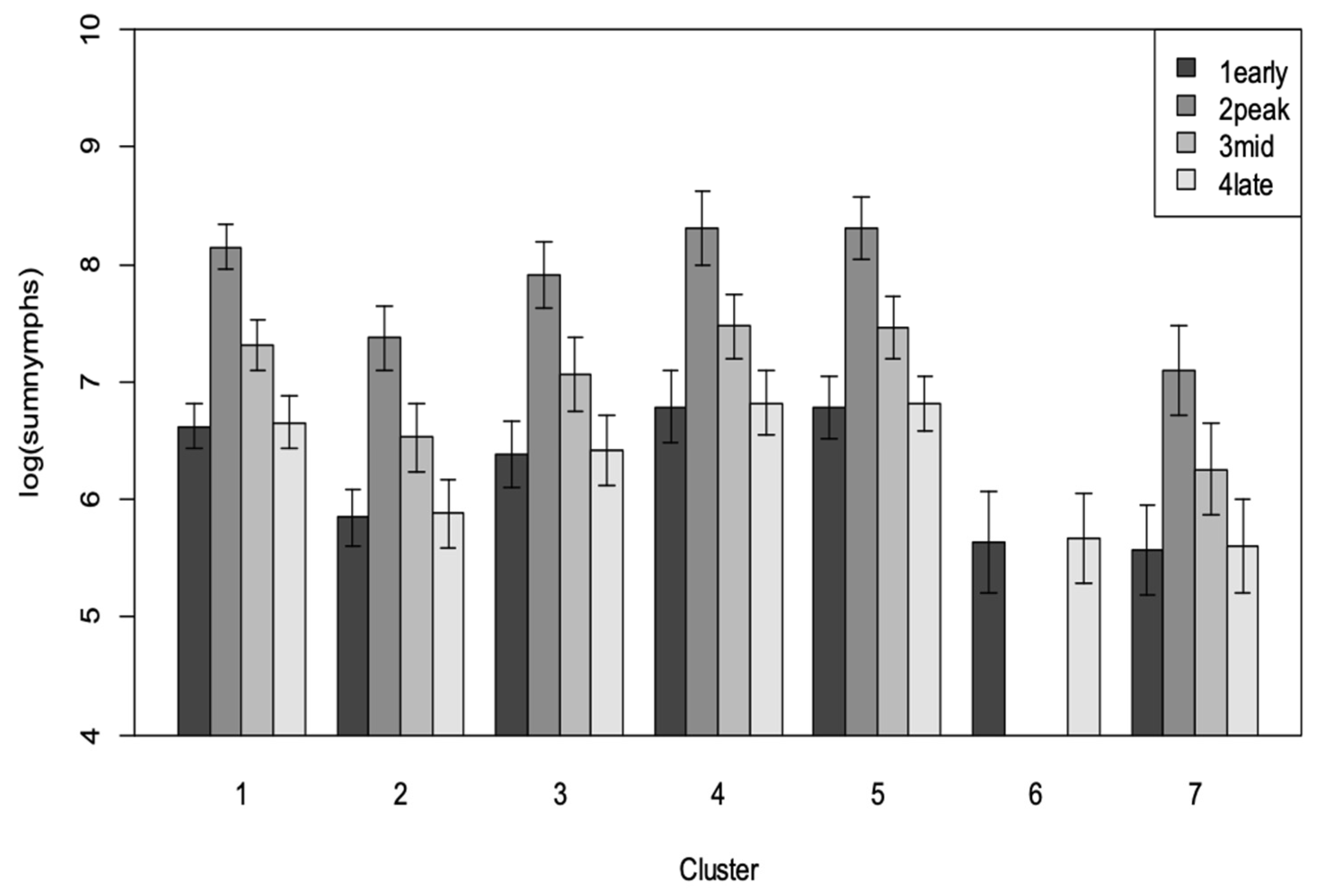
3.2. Research Question 1: Does the Area and Age of Cassava in the Landscape at Any Point in Time Correlate Positively with Bemisia Species Abundance in the Focal Field?
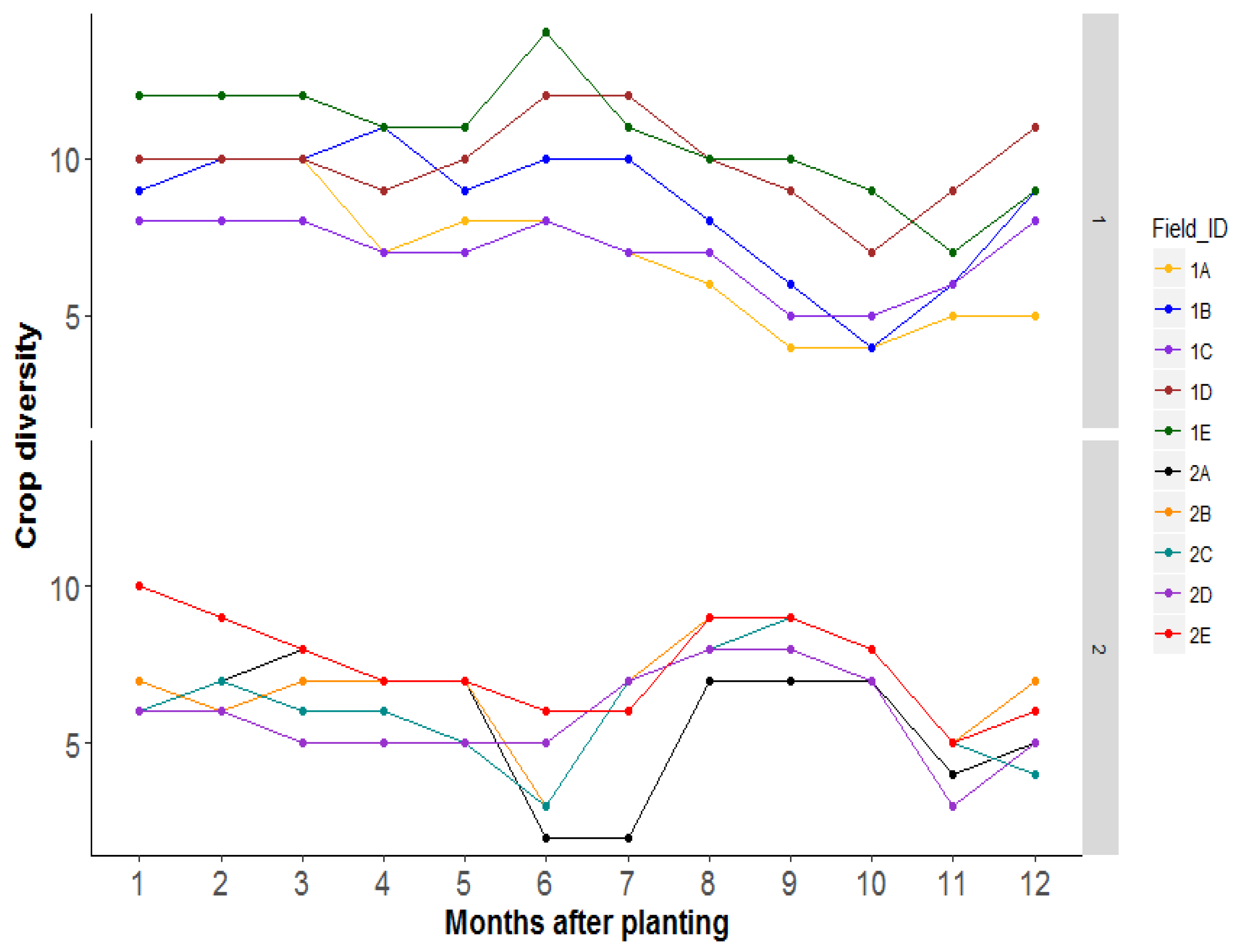
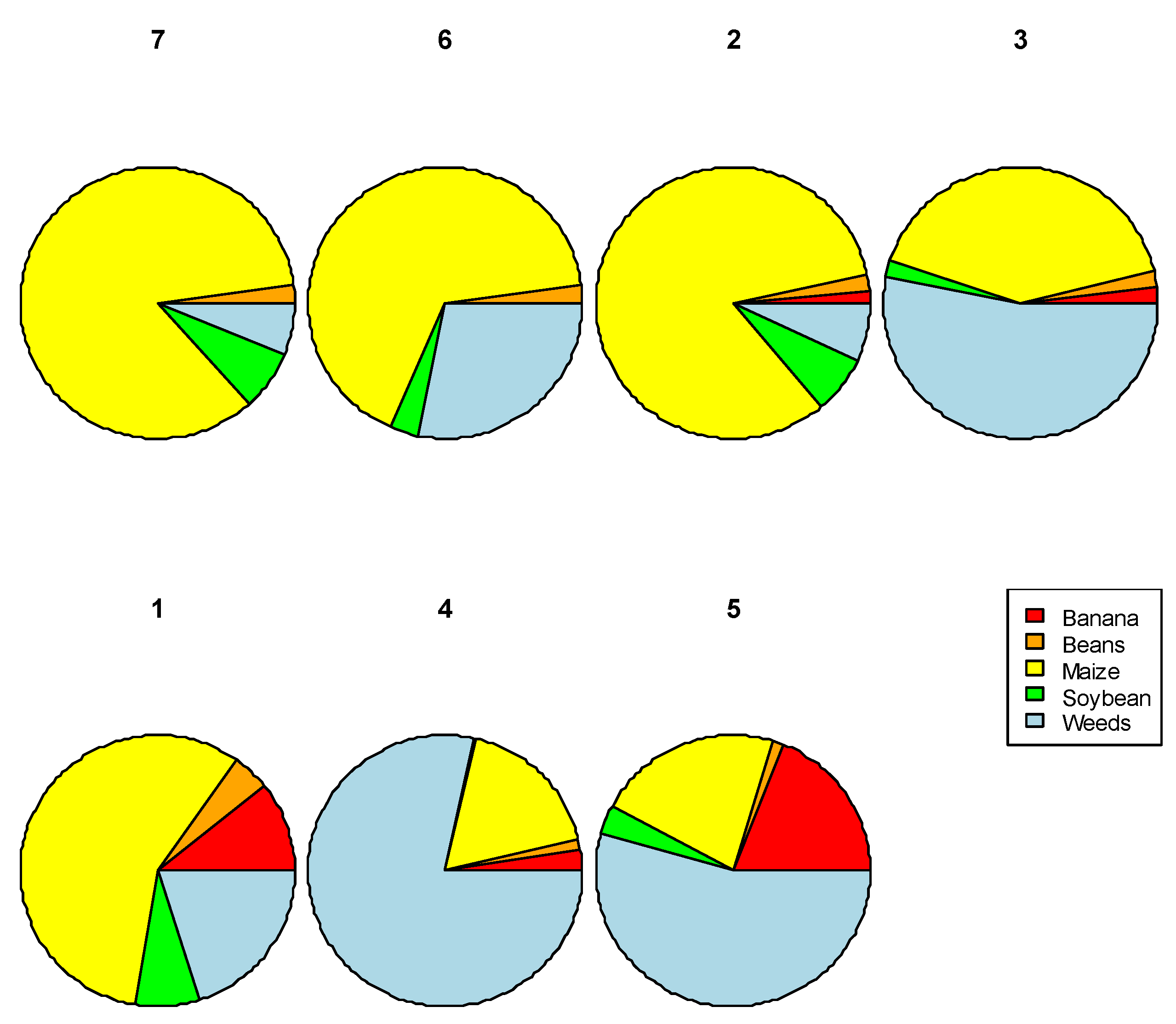
3.3. Research Question 2: What Combination of the Dominant Crop Types in the Landscape Correlates Positively with Bemisia Species Abundance in the Focal Field?
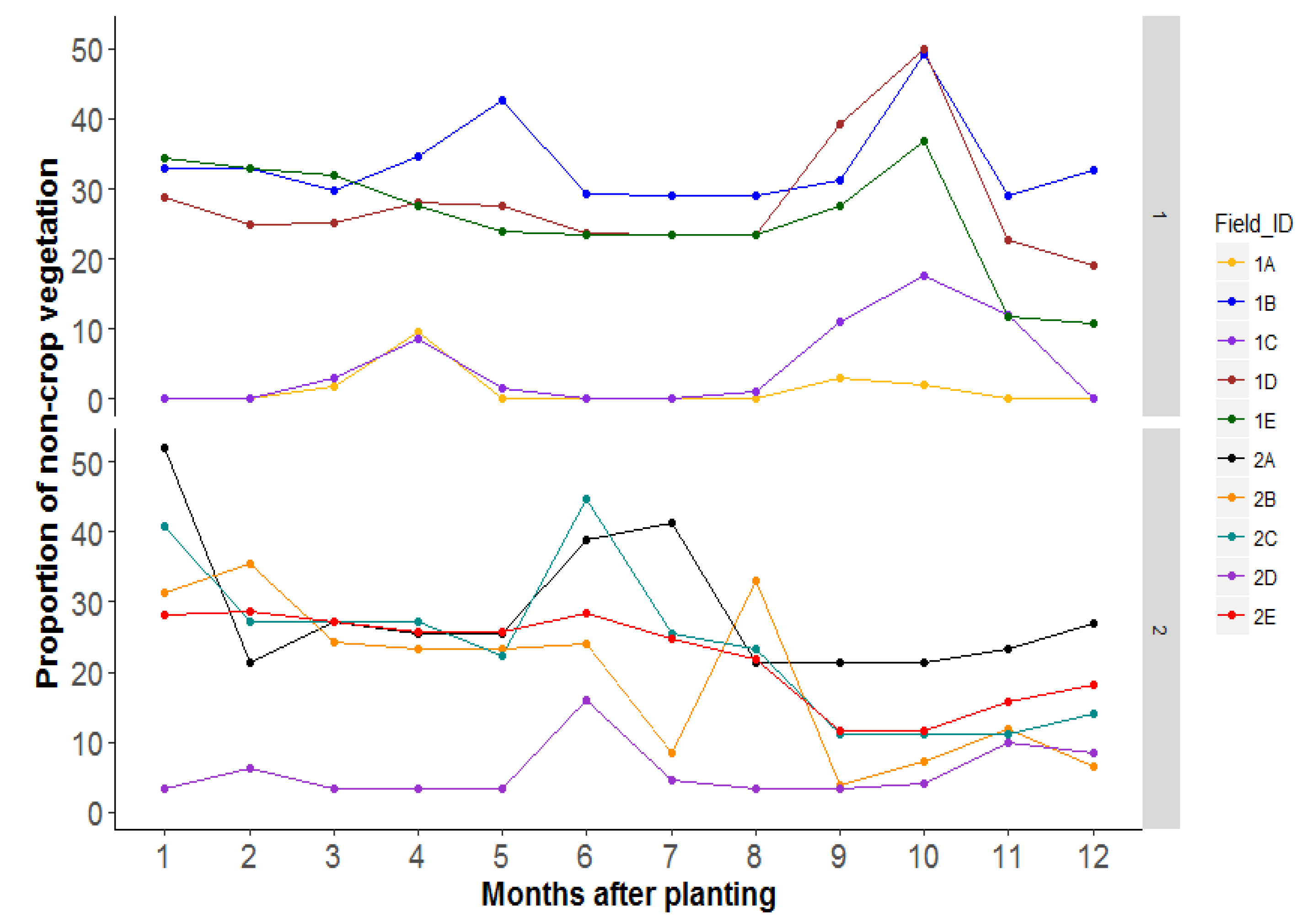
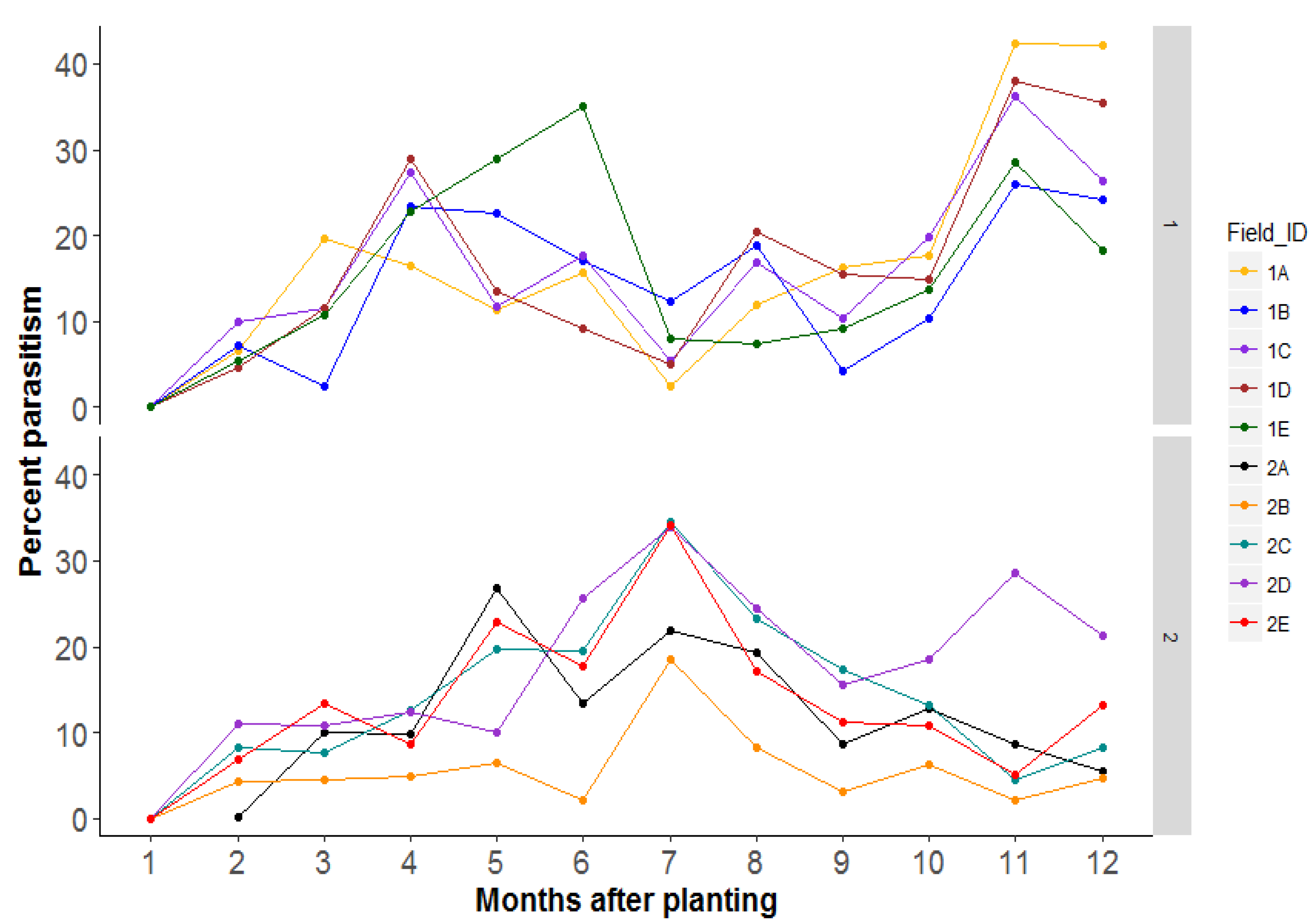
3.4. Research Question 3: Does an Increased Area of Semi-Natural Habitats Lead to Increased Parasitism of Bemisia Species Nymphs in the Focal Field?
4. Discussion
4.1. Research Question 1
4.2. Research Question 2
4.3. Research Question 3
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bianchi, F.J.J.; Booij, C.J.; Tscharntke, T. Sustainable pest regulation in agricultural landscapes: A review on landscape composition, biodiversity and natural pest control. Proc. R. Soc. B Biol. Sci. 2006, 273, 1715–1727. [Google Scholar] [CrossRef] [Green Version]
- Schneider, G.; Krauss, J.; Riedinger, V.; Holzschuh, A.; Steffan-Dewenter, I. Biological pest control and yields depend on spatial and temporal crop cover dynamics. J. Appl. Ecol. 2015, 52, 1283–1292. [Google Scholar] [CrossRef]
- Knops, J.J.M.; Tilman, D.; Haddad, N.M.; Naeem, S.; Mitchell, C.E.; Haarstad, J.; Ritchie, M.E.; Howe, K.M.; Reich, P.B.; Siemann, E.; et al. Effects of plant species richness on invasion dynamics, disease outbreaks, insect abundances and diversity. Ecol. Lett. 1999, 2, 286–293. [Google Scholar] [CrossRef]
- Vidal, M.C.; Murphy, S.M. Bottom up vs. top-down effects on terrestrial insect herbivores: A meta-analysis. Ecol. Lett. 2018, 21. [Google Scholar] [CrossRef] [PubMed]
- Sequeira, R.V.; Shields, A.; Moore, A.; De Barro, P. Inter-seasonal population dynamics and pest status of Bemisia tabaci (Gennadius) biotype B in an Australian cropping system. Bull. Entomol. Res. 2009, 99, 325. [Google Scholar] [CrossRef] [Green Version]
- De Barro, P. Getting the Most out of Eretmocerus Hayati, an Effective Natural Enemy of Silverleaf Whitefly; Final Report 2012, VG08051, Elizabeth Street Sydney NSW; Horticulture Australia Ltd.: Sydney, Australia, 2012. [Google Scholar]
- FAOSTAT. FAOSTAT Database 2019; Food and Agriculture Organization of the United Nations: Rome, Italy; Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 23 February 2020).
- Legg, J.P.; Sseruwagi, P.; Boniface, S.; Okao-Okuja, G.; Shirima, R.; Bigirimana, S.; Gashaka, G.; Herrmann, H.-W.; Jeremiah, S.; Brown, J.K.; et al. Spatio-temporal patterns of genetic change amongst populations of cassava Bemisia tabaci whiteflies driving virus pandemics in East and Central Africa. Virus Res. 2014, 186, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Pennisi, E. Armed and dangerous. Science 2010, 327, 804–805. [Google Scholar] [PubMed] [Green Version]
- Macfadyen, S.; Paull, C.; Boykin, L.M.; De Barro, P.; Maruthi, M.; Otim, M.; Kalyebi, A.; Vassão, D.; Sseruwagi, P.; Tay, W.; et al. Cassava whitefly, Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) in East African farming landscapes: A review of the factors determining abundance. Bull. Entomol. Res. 2018, 108, 565–582. [Google Scholar] [CrossRef] [Green Version]
- Abele, S.; Twine, E.; Ntawuruhunga, P.; Baguma, Y.; Kanobe, C.; Bua, A. Development and dissemination of improved cassava varieties in Uganda: Analysis of adoption rates, variety attributes and speed of adoption. AAAE Ghana Conf. Proc. 2007, 52198. [Google Scholar] [CrossRef]
- Tumwegamire, S.; Kanju, E.; Legg, J.; Shirima, R.; Kombo, S.; Mkamilo, G.; Mtunda, K.; Sichalwe, K.; Kulembeka, H.; Ndyetabura, I.; et al. Exchanging and managing invitro elite germplasm to combat Cassava Brown Streak Disease (CBSD) and Cassava Mosaic Disease (CMD) in Eastern and Southern Africa. Food Secur. 2018, 10, 351–368. [Google Scholar] [CrossRef] [Green Version]
- Legg, J.P.; Jeremiah, S.C.; Obiero, H.M.; Maruthi, M.N.; Ndyetabula, I.; Okao-Okuja, G.; Bouwmeester, H.; Bigirimana, S.; Tata-Hangy, W.; Kumar, P.L.; et al. Comparing the regional epidemiology of the cassava mosaic and cassava brown streak virus pandemics in Africa. Virus Res. 2011, 159, 161–170. [Google Scholar] [CrossRef]
- Otim, M.; Legg, J.; Kyamanywa, S.; Polaszek, A.; Gerling, D. Occurrence and activity of Bemisia tabaci parasitoids on cassava in different agro-ecologies in Uganda. BioControl 2005, 50, 87–95. [Google Scholar] [CrossRef]
- Otim, M.; Kyalo, G.; Kyamanywa, S.; Asiimwe, P.; Legg, J.P.; Guershon, M.; Gerling, D. Parasitism of Bemisia tabaci (Homoptera: Aleyrodidae) by Eretmocerus mundus (Hymenoptera: Aphelinidae) on cassava. Int. J. Trop. Insect Sci. 2008, 28, 158. [Google Scholar] [CrossRef]
- Guastella, D.; Lulah, H.; Tajebe, L.S.; Cavalieri, V.; A Evans, G.; Pedata, P.A.; Rapisarda, C.; Legg, J.P. Survey on whiteflies and their parasitoids in cassava mosaic pandemic areas of Tanzania using morphological and molecular techniques. Pest Manag. Sci. 2015, 71, 383–394. [Google Scholar] [CrossRef] [PubMed]
- Macfadyen, S.; Tay, W.T.; Hulthen, A.; Paull, C.; Kalyebi, A.; Jacomb, F.; Parry, H.; Sseruwagi, P.; Seguni, Z.; Omongo, C.A.; et al. Landscape factors and how they influence whitefly pests in cassava fields across east Africa. Landsc. Ecol. 2020. [Google Scholar] [CrossRef]
- Wormann, C.S.; Eledu, C.A. Uganda’s Agroecological Zones: A Guide for Planners and Policy Makers; Centro International de Agriculture Tropicale: Kampala, Uganda, 1999. [Google Scholar]
- Kriticos, D.J.; Darnell, R.E.; Yonow, T.; Ota, N.; Sutherst, R.W.; Parry, H.R.; Mugerwa, H.; Maruthi, M.N.; Seal, S.E.; Colvin, J.; et al. Improving climate suitability for Bemisia tabaci in East Africa is correlated with increased prevalence of whiteflies and cassava diseases. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- ESRI-Environmental Systems Research Institute, Inc. Collector for ArcGIS Version 10.4; ESRI: Redlands, CA, USA, 2015. [Google Scholar]
- ESRI-Environmental Systems Research Institute, Inc. ArcGIS Desktop Version 10.2; ESRI: Redlands, CA, USA, 2010. [Google Scholar]
- Tay, W.T.; Court, L.; Macfadyen, S.; Jacomb, F.; Vyskočilová, S.; Colvin, J.; De Barro, P.J. A High-Throughput Amplicon sequencing approach for population-wide species diversity and composition survey’. Preprint. Evol. Biol. 2020. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Kunz, D.; Tay, W.T.; Elfekih, S.; Gordon, K.H.J.; De Barro, P.J. Take out the rubbish—Removing NUMTs and pseudogenes from the Bemisia tabaci cryptic species mtCOI database. bioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- McGarigal, K.; Cushman, S.A.; Ene, E. FRAGSTATS v4: Spatial Pattern Analysis Program for Categorical and Continuous Maps, Computer Software Program Produced by the Authors at the University of Massachusetts, Amherst. 2012. Available online: http://www.umass.edu/landeco/research/fragstats/fragstats.html (accessed on 18 March 2018).
- Sseruwagi, P.; Otim-Nape, G.W.; Osiru, D.S.O.; Thresh, J.M. Influence of NPK fertilizer on populations of the whitefly vector and incidence of cassavamosaic virus disease. Afr. Crop Sci. J. 2003, 11, 171–179. [Google Scholar]
- R Core Team. The R Foundation for Statistical Computing. 2018. Available online: https://www.r-project.org/ (accessed on 1 December 2018).
- Romesburg, H.C. Cluster Analysis for Researchers; Lifetime Learning Publication: Belmont, CS, USA, 1984. [Google Scholar]
- Kalyebi, A.; Macfadyen, S.; Parry, H.; Tay, W.T.; De Barro, P.; Colvin, J. African cassava whitefly, Bemisia tabaci, cassava colonization preferences and control implications. PLoS ONE 2018, 13, e0204862. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, I.U.; Ghosh, S.; Maruthi, M.N. Host and virus effects on reversion in cassava affected by cassava brown streak disease. Plant Pathol. 2016, 65, 593–600. [Google Scholar] [CrossRef] [Green Version]
- Malka, O.; Santos-Garcia, D.; Feldmesser, E.; Sharon, E.; Krause-Sakate, R.; Delatte, H.; Van Brunschot, S.; Patel, M.; Visendi, P.; Mugerwa, H.; et al. Species-complex diversification and host plant associations in Bemisia tabaci: A plant-defence, detoxification perspective revealed by RNA-Seq analyses. Mol. Ecol. 2018, 27, 4241–4256. [Google Scholar] [CrossRef] [Green Version]
- Abd-Rabou, S.; Simmons, A.M. Survey of reproductive host plants of Bemisia tabaci (Hemiptera: Aleyrodidae) in Egypt, including new host records. Entomol. News 2019, 121, 456–465. [Google Scholar] [CrossRef]
- Smilanich, A.M.; Malia, F.R.; Dyer, L.A. Does plant apparency matter? Thirty years of data provide limited support but reveal clear patterns of the effects of plant chemistry on herbivores. New Phytol. 2016, 210, 1044–1057. [Google Scholar] [CrossRef] [Green Version]
- Fargette, D.; Fauquet, C. A preliminary study on the influence of intercropping maize and cassava on the spread of African cassava mosaic virus by whiteflies. Asp Appl. Biol. 1988, 17, 195–202. [Google Scholar]
- Quintela, E.D.; Abreu, A.G.; Lima, J.F.d.S.; Mascarin, G.M.; Santos, J.B.D.; Brown, J.K. Reproduction of the whitefly B. tabaci (Hemiptera: Aleyrodidae) B biotype in maize fields. Pest Manag. Sci. 2016, 72, 2181–2187. [Google Scholar] [CrossRef]
- Boykin, L.M.; Bell, C.D.; Evans, G.; Small, I.; De Barro, P.J. Is agriculture driving the diversification of the Bemisia tabaci species complex (Hemiptera: Sternorrhyncha: Aleyrodidae)?: Dating, diversification and biogeographic evidence revealed. BMC Evol. Biol. 2013, 13, 228. [Google Scholar] [CrossRef] [Green Version]
- Gold, C.S.; Altieri, M.A.; Bellotti, A.C. The effects of intercropping and mixed varieties of predators and parasitoids of cassava whiteflies (Hemiptera, Aleyrodidae) in Colombia. Bull. Entomol. Res. 1989, 79, 115–121. [Google Scholar] [CrossRef]
- Uzokwe, V.N.E.; Mlay, D.P.; Masunga, H.R.; Kanju, E.; Odeh, I.O.; Onyeka, J. Combating viral mosaic disease of cassava in the Lake Zone of Tanzania by intercropping with legumes. Crop Prot. 2016, 84, 69–80. [Google Scholar] [CrossRef] [Green Version]
- Ellango, R.; Singh, S.T.; Rana, V.S. Priya, N.G.; Raina, H.; Chaubey, R.; Naveen, N.C.; Mahmood, R.; Ramamurthy, V.V.; Asokan, R.; et al. Distribution of Bemisia tabaci genetic groups in India. Environ. Entomol. 2015, 44, 1258–1264. [Google Scholar] [CrossRef]
- Chaplin-Kramer, R.; O’Rourke, M.E.; Blitzer, E.J.; Kremen, C. A meta-analysis of crop pest and natural enemy response to landscape complexity. Ecol. Lett. 2011, 14, 922–932. [Google Scholar] [CrossRef]
- Veres, A.; Petit, S.; Conord, C.; Lavigne, C. Does landscape composition affect pest abundance and their control by natural enemies? A review. Agric. Ecosyst. Environ. 2013, 166, 110–117. [Google Scholar] [CrossRef]
- Karp, D.S.; Chaplin-Kramer, R.; Meehan, T.D.; Martin, E.A.; Declerck, F.; Grab, H.; Gratton, C.; Hunt, L.; Larsen, A.E.; Martínez-Salinas, A.; et al. Crop pests and predators exhibit inconsistent responses to surrounding landscape composition. Proc. Natl. Acad. Sci. USA 2018, 115, E7863–E7870. [Google Scholar] [CrossRef] [Green Version]
- Wyckhuys, K.A.G.; Burra, D.D.; Tran, D.H.; Graziosi, I.; Walter, A.J.; Nguyen, T.G.; Trong, H.N.; Le, B.V.; Le, T.T.N.; Fonte, S.J. Soil fertility regulates invasive herbivore performance and top-down control in tropical agro-ecosystems of Southeast Asia. Agric. Ecosyst. Environ. 2017, 249, 38–49. [Google Scholar] [CrossRef]
- Le, T.T.N.; Graziosi, I.; Cira, T.M.; Gates, M.; Parker, L.; Wyckhuys, K. Landscape context does not constrain biological control of Phenacoccus manihoti in intensified cassava systems of southern Vietnam. Biol. Control 2018, 121, 129–139. [Google Scholar] [CrossRef]
- Gurr, G.M.; Wratten, S.D.; Landis, D.A.; You, M. Habitat management to suppress pest populations: Progress and prospects. Annu. Rev. Entomol. 2017, 62, 91–109. [Google Scholar] [CrossRef] [PubMed]
- Parry, H.R.; Kalyebi, A.; Bianchi, F.; Sseruwagi, P.; Colvin, J.; Schellhorn, N.; MacFadyen, S. Evaluation of cultural control and resistance-breeding strategies for suppression of whitefly infestation of cassava at the landscape scale: A simulation modeling approach. Pest Manag. Sci. 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Research Question | Name | Description |
|---|---|---|
| In-Field Factors | ||
| 1,2,3 | Time/Months after planting (MAP) | Represented as the age of the focal cassava (months after planting). There were 12 levels of this factor (1,2,3,4,5,6,7,8,9,10,11,12). |
| 1,2,3 | Field/site | These are the 10 different landscapes represented as (1A,1B,1C,1D,1E,2A,2B,2C,2D,2E) planted out during the first rains and second rains. |
| Landscape Factors that Change Across Time | ||
| 1,2 | Cassava | Percentage area covered by cassava † |
| 2 | Sweet potato | Percentage area covered by sweet potato † |
| 2 | Soybean | Percentage area covered by soybean † |
| 2 | Groundnuts | Percentage area covered by groundnuts † |
| 2 | Sesame | Percentage area covered by sesame † |
| 2 | Beans | Percentage area covered by beans † |
| 2 | Maize | Percentage area covered by maize † |
| 2 | Coffee, Rice, Banana, Bambara, Pumpkin, Mango, Citrus | Percentage area covered by each of these crops † |
| 2,3 | Weeds | Percentage area covered by weeds † |
| 3 | Non-crop | Percentage area covered by non-crop (native vegetation/woodland) † |
| 1 | Young cassava (less suitable) | Percentage area covered by cassava 0–3 MAP † |
| 1 | Ideal cassava (most suitable) | Percentage area covered by cassava 3–7 MAP † |
| 1 | Old cassava (least suitable) | Percentage area covered by cassava greater than 7 MAP † |
| Bemisia tabaci Sample Code and Source Host Crop | Similarity with Published GenBank Sequences | Identity (%) |
|---|---|---|
| LF (Beans) | Bemisia Uganda1_KX570868 | 100 |
| DF1 (Sesame) | Mediterranean_UG254_KX570768 | 100 |
| CF † (Groundnut) | SubSahAf1_Uganda_KX570800 | 88.65 |
| GF1 (Soybean) | SubSahAf1_Uganda_Masaka_AY903462 | 100 |
| HF(Euphorbia) | SubSaharan Africa 13_KX570833 | 99.35 |
| GF2 (Soybean) | Bemisia Uganda1_KX570858 | 99.85 |
| IF (Sweet potato) | Bemisia Uganda1_KX570863 | 99.85 |
| EF (Beans) | Bemisia Uganda1_KX570863 | 99.85 |
| BF (Sweet potato) | Mediterranean_UG254_KX570768 | 99.70 |
| MF (Pumpkin) | Mediterranean_Uganda_ASL_MH205754 | 100 |
| KF (Sesame) | Mediterranean_UG254_KX570768 | 99.85 |
| AR (Sweet potato) | Bemisia Uganda1_KX570868 | 100 |
| NR (Soybean) | SubSahAf1_Uganda_KX570785 | 100 |
| FR (Soybean) | SubSahAf1_Uganda_KX570785 | 100 |
| DF2 (Sesame) | SubSahAf1_Uganda_KX570785 | 100 |
| Crop Categorization | Relationship with Nymph Abundance in Cassava | p-Value |
|---|---|---|
| Cassava * | 0.10 | 0.23 |
| Cassava old * | –0.093 | 0.05 |
| Cassava young * | –0.019 | 0.04 |
| Cassava ideal * | 0.36 | 0.0002 |
| Maize | –0.40 | <0.001 |
| Beans * | –0.22 | <0.01 |
| Eggplant | 0.28 | 0.002 |
| Soybean * | –0.095 | 0.03 |
| Sweet potato * | –0.22 | 0.0002 |
| Rice | 0.16 | 0.04 |
| Banana | 0.23 | 0.009 |
| Citrus | 0.17 | 0.02 |
| Cocoyam | 0.20 | 0.05 |
| Bambara | –0.011 | 0.69 |
| Coffee | 0.76 | 0.035 |
| Cowpea | –0.03 | 0.72 |
| Groundnut * | 0.019 | 0.02 |
| Weeds | –0.049 | 0.56 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalyebi, A.; Macfadyen, S.; Hulthen, A.; Ocitti, P.; Jacomb, F.; Tay, W.T.; Colvin, J.; De Barro, P. Within-Season Changes in Land-Use Impact Pest Abundance in Smallholder African Cassava Production Systems. Insects 2021, 12, 269. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12030269
Kalyebi A, Macfadyen S, Hulthen A, Ocitti P, Jacomb F, Tay WT, Colvin J, De Barro P. Within-Season Changes in Land-Use Impact Pest Abundance in Smallholder African Cassava Production Systems. Insects. 2021; 12(3):269. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12030269
Chicago/Turabian StyleKalyebi, Andrew, Sarina Macfadyen, Andrew Hulthen, Patrick Ocitti, Frances Jacomb, Wee Tek Tay, John Colvin, and Paul De Barro. 2021. "Within-Season Changes in Land-Use Impact Pest Abundance in Smallholder African Cassava Production Systems" Insects 12, no. 3: 269. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12030269