1. Introduction
The Asian tiger mosquito
Aedes albopictus (Skuse, 1894) is an invasive species responsible for the transmission of arboviruses such as dengue, Zika and chikungunya [
1]. In recent years, the distribution of
A. albopictus and
Aedes aegypti has increased drastically in all continents, and consequently, these arboviral diseases have become a global health concern [
2].
Aedes albopictus was first detected in Spain (Catalonia) in 2004 [
3], and since then, it has spread in all Mediterranean regions of Spain and in Basque Country [
4]. In 2018, the first autochthonous cases of dengue were detected in Spain, which provided evidence of
A. albopictus being an effective vector of this virus in continental Europe [
5,
6,
7]. In the absence of efficient vaccines to prevent these diseases, vector control remains a key strategy. Over several decades, the fight against vector-borne human diseases has been based on the use of insecticides. However, the resistance of mosquitoes against these chemicals has been widely reported, and few insecticides are currently approved for public health use [
8]. Consequently, more sustainable and environment-friendly vector control tools are required. The sterile insect technique (SIT) is a birth control method which consists of the production and release of sterile males to mate with wild females in the field. As a consequence, sterility is induced in the native wild female population, which will decline over the generations [
9].
First studies on the application of SIT against mosquitoes started in the 1960s. In 1967, an isolated population of
Culex quinquefasciatus was successfully eliminated in Myanmar by releasing mosquitoes which were sterile due to
Wolbachia-induced cytoplasmic incompatibility [
10]. The release of chemosterilized males resulted in suppression and elimination of
Culex quinquefasciatus on an island of Florida in 1969 [
11]. The same strategy was deployed in El Salvador, achieving a 99% reduction in
Anopheles albimanus wild population [
12]. Interest in this technique was only recently revived and focused on its potential as a tool for the population suppression of
Aedes mosquitoes. Several SIT trials have been initiated in different regions of the world to fight against mosquito vectors and they are currently in their initial phase [
13]. Their successful application depends on many parameters including the ability of mass-rearing facilities to produce sufficient numbers of sterile males [
9,
14]. The SIT-incompatible insect technique (IIT) program in Guangzhou of China is currently the largest release program, producing and releasing more than 160,000
Wolbachia-infected and irradiated male
A. albopictus per hectare each week in 2016 and 2017 [
15].
In Europe, the first pilot field trials of SIT have been performed in three villages of North Italy, releasing
A. albopictus males sterilized by gamma radiation. The sterility level in the wild population reached 70–80%, followed by a similar reduction in the egg density recorded in the ovitraps [
16].
The increasing interest in the application of SIT against mosquitoes is accelerating the development of equipment, guidelines and operational protocols for rearing, sex sorting, irradiation, transport and release of sterile male mosquitoes. There is considerable literature available in respect to methods and devices for mass-rearing at different stages [
17,
18,
19,
20,
21,
22,
23]. However, specific information on the different production strategies implemented in the ongoing programs is missing. In addition, in view of several ongoing and planned SIT field trials against
Aedes mosquitoes, there is a lack of referenced and standardized mass-rearing methodology which would support new programs.
The Agriculture Department of the Valencian Region has been promoting a pilot project to evaluate the efficacy of an integrated vector management program (IVM) based on the use of the SIT as the main method of control. The project roadmap has been based on the phased conditional approach framework proposed by World Health Organization and the International Atomic Energy Agency (WHO-IAEA) for the implementation of SIT programs [
13]. Phase I described in these guidelines focuses on the definition of laboratory studies to assess the feasibility of the SIT when it will be applied on the field. On the other hand, phase II involves semi-field and field trials in order to adjust the technique before scaling it up to larger areas. Both phases have been fulfilled in this project over four years.
The present study reports the laboratory studies performed in phase I for evaluating the entomological efficacy of SIT. Moreover, the different steps of the mass-rearing process and quality control protocols for sterile male A. albopictus in support of the SIT program in the Valencia Region are detailed. The key parameters that describe the production process are analyzed to evaluate its efficiency. The results will allow comparison of the described process with those followed in different programs and will contribute to the development of a standardized procedure for the mass-rearing of Aedes mosquitoes for SIT applications.
4. Discussion
The objective of the SIT pilot project against
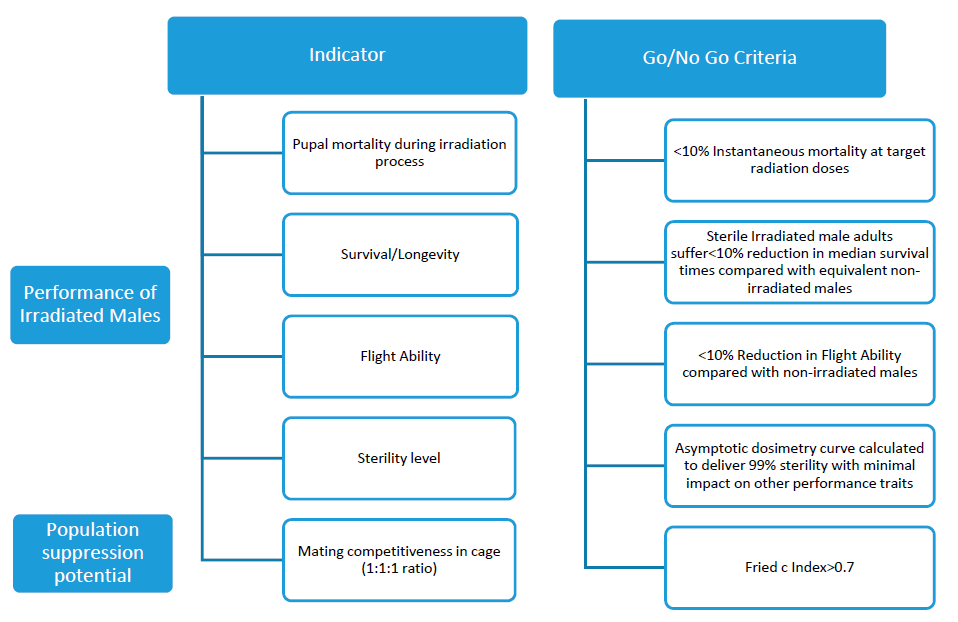
A. albopictus in the Valencian Region was to evaluate the use of the sterile insect technique as part of an integrated vector control program. Prior to the mass production and field release phase, the suitability of the project was evaluated at a laboratory level following the criteria proposed by WHO-IAEA [
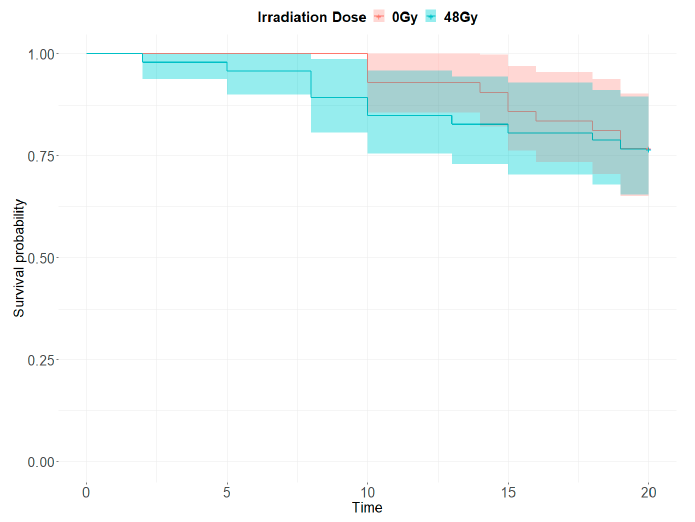
13]. All criteria in this document were met, supporting the transition to a second phase consisting of increasing production to test the release of sterile males in the selected areas. The irradiation dose of 48 Gy needed to achieve 99% sterility in males seems high compared to previous experiments [
40,
41,
42,
43]. Nonetheless, there is recent evidence that several factors such as pupae age and oxygen availability affect the response to radiation in mosquitoes [
44,
45]. In addition, in small self-contained gamma irradiators such as the Gammacell 220, the irradiation intensity is spatially heterogeneous and only the center of the irradiation chamber receives the dose of reference. These factors are difficult to control in mass production, and a higher dose assures that all the pupae are treated over a minimal irradiation threshold. Ensuring acceptable sterility implies a loss of quality, which is reflected in the post-emergence flight ability but not in the longevity test. Irradiation may also affect the male mating competitiveness. In our case, the mean value of Fried Index c was 0.92, which is higher than the value recommended in the WHO-IAEA “Guidance Framework for Testing the Sterile Insect Technique as a Vector Control Tool against
Aedes-Borne Diseases” [
13] (c > 0.7).
The sterile insect technique and other rear-and-release vector control methods, such as IIT or the Release of Insects carrying a Dominant Lethal (RIDL) [
46], depend on the capacity to produce large quantities of insects with an adequate quality that ensures their effectiveness in the field. Therefore, the optimization of production parameters is key to obtain efficient production in a successful SIT program against vector mosquitoes. There is extensive literature describing optimized rearing methods and their outcomes in experimental contexts [
20,
47,
48,
49,
50,
51]. References showing real parameters of a mass production in practice are, however, scarce [
16,
52], as there are few projects that produce sterile males on a medium or large scale.
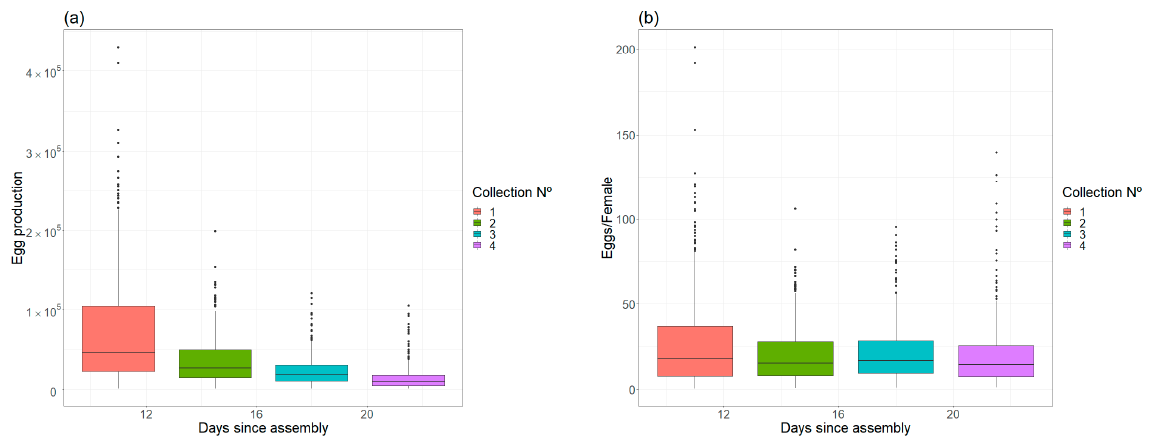
The final objective of adult rearing is the production of eggs, and maximization of fecundity is desirable. Research efforts have usually been focused on finding the optimal shape and vertical resting surface [
47,
53], sex ratio [
47,
54] and blood feeding methods [
54,
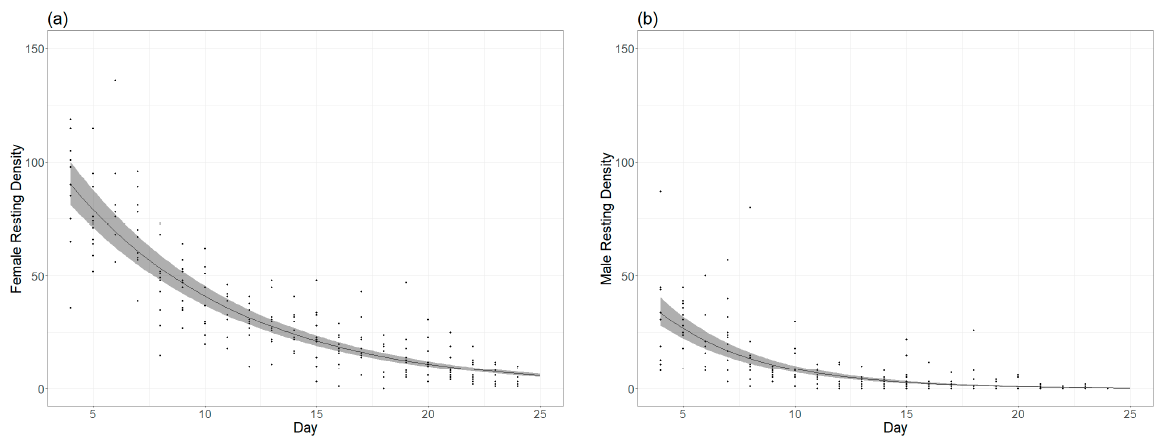
55]. In our rearing cages, egg production was highly variable (
Figure 6), probably influenced by blood quality and rearing experience. The number of eggs laid per initial female pupae reached an average of 21.03. Even taking into account differences due to biotic factors such as different strains, and also due to colonization processes, we consider that this productivity is relatively low compared to laboratory studies [
20,
47,
54,
56], although the rearing methods were basically equivalent. There is no comparable information for mass-rearing programs. The chosen design of the adult cage probably did not influence the productivity. According to Balestrino et al. [
53], medium-sized cubic cages, such as the ones used in our program, are similar in productivity to large space-optimized cages, and Zhang et al. [
48] found that short cages allow for higher blood-feeding rates and egg production. The vertical resting surface density (1.56 mosquitoes/cm
2) was similar to the optimal density proposed by Zhang et al. [
47] (1.48 mosquitoes/cm
2), although both are high compared to the density of 0.8 mosquitoes/cm
2 used in similar experiments [
53,
54,
56]. It should be noted that the results may have been affected by the method of estimating the number of produced eggs. Visual egg estimation is a fast method in the context of mass production when personnel are trained but needs to be improved to obtain a more accurate measure.
Concerning the life cycle of rearing adult cages, Zhang et al. and Maïga et al. [
47,
56] suggested a two-week cycle for
A. albopictus in a medium-scale rearing facility to increase efficiency per cage. In our rearing process, 74% of the eggs were collected in the first two weeks, and 26% in the third one. Currently, the cost of dismantling, cleaning and reassembling each cage is high, and the maintenance of the cages for three weeks makes production more cost-effective.
The total egg production per cage is related to the fecundity of females but, apparently, also to the mortality rate. As described in the Results section, egg production per surviving female remained constant during the four collection dates, with female mortality likely being the factor that reduced the productivity of the cage over time. The fecundity per estimated female in the first gonotrophic cycle can be considered low (30 eggs/female) compared with laboratory studies (mean value ranging from 13 to 143 eggs per female) [
53,
55,
57,
58,
59,
60,
61]. In any case, the methodology used to estimate the fecundity of the females was different from the referenced works, since it was obtained from eggs collected per cage and the survival curve was obtained according to the date of collection, and some artefact may be influencing the results. This methodology was used because the objective of the presented values was to evaluate the productivity of the rearing cages in a routine mass-rearing context. The survival of females was similar to that reported by Maiga et al. [
56] with a comparable methodology (88.5%). In our colony, the mortality of males was higher than females. Maiga et al. reported similar decay rates between males and females with an average density, but Balestrino et al. [
53] found that differences in mortality between sexes were linked to density. It is possible, then, that males are more affected by overcrowding than females.
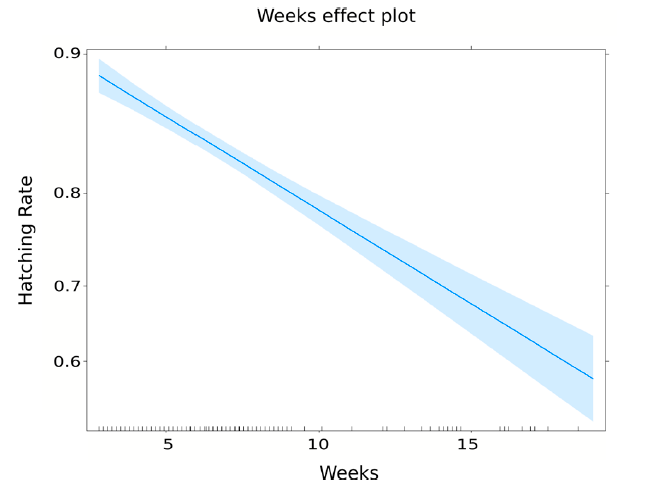
The hatching rate of the eggs decreased over time in storage, as has been generally observed [
17]. It is, therefore, desirable to define a strategy that balances the maintenance of an egg reserve and an acceptable hatching rate. A stock of eggs is advisable to compensate for unforeseen events in production. The hatching rate can be kept above 80% for nine weeks, as described in
Table 5. These results are consistent with those obtained by Zheng et al. [
17], which obtained a hatching rate greater than 80% after 10 weeks using nutrient broth as a hatching medium. Longer-term egg conservation is highly recommended in temperate regions with a stationary wild population dynamic. In the period when field populations are low or non-existent, it would be possible to continue the production of an egg bank, which would reinforce the high population season releases.
The goal of larval rearing is not only to provide pupae for the production of sterile adult males to be released, but also to restock the egg colony. In our program, male pupae were sex sorted on the 8th day after hatching. Of the initial L1 larvae (males and females), 17.2% of male pupae were recovered at sex sorting to be irradiated and released in the field. The rest of the pupae produced in the trays remained available to maintain the rearing colony. These values are slightly higher than the productivities of equivalent mass rearing programs with
A. albopictus [
16] or
A. aegypti [
52], but significantly lower than the values for simulations of mass-rearing obtained in laboratory experiments [
48,
49,
50]. The use of a multi-step sex sorting method based on artificial intelligence can recover almost the totality of the male production [
62].
The average contamination rate of females in the batches of the released males was 0.15% using the Fay–Morlan apparatus [
26]. Carvalho et al. [
52], using a similar device, released only 0.02% of females, probably related to the more efficient size separation of
A. aegypti due its more pronounced sexual dimorphism [
63]. A more sophisticated method was used by Crawford et al. to release a female every 900 million males [
62]. On the other hand, Bellini et al. [
16], using a less efficient separation method with
A. albopictus, obtained 1.21% females. Compared to larval rearing, the labor invested in sex sorting is considerably high [
64,
65] and its optimization could boost the productivity in mosquito mass-rearing facilities. New approaches to mosquito sex sorting are being developed to develop large-scale programs against vector mosquitoes [
62,
63,
66,
67]. The parameters derived from laboratory experiments likely overestimate the productivity since they are obtained in ideal conditions, but parameters derived from a real context offer a more informative point of view of the process. The availability of realistic production parameters is useful as a reference for the design of future projects and it is necessary in order to understand and optimize the production process. Currently, there are few projects that produce sterile males of mosquitoes on a medium or larger scale that can offer this kind of information. It is also noteworthy that during the four years of the project, parameters such as the number of eggs per female or male production increased considerably, partly due to an adaptation of the colony to the rearing conditions, but also mainly due to improved procedures and training of the staff.
In addition, most of the designs and methods proposed for mosquito rearing tend to be biologically optimized (gross production per individual) rather than in terms of handling efficiency (gross production per hour of labor). Despite the fact that larger cages and larval rearing trays present a greater capacity and allow a reduction in the number of units, their handling and cleaning become more complex than small units due to bigger dimensions. In order to optimize cost efficiency in a large number of small cages, efforts are being made to automate the rearing processes. In this sense, adaptation to industrial stacking containers such as the Standard Euro Containers can allow easier adaptation of washing, storage or handling systems already existing in the industry, which are usually highly optimized due to their wide range of applications.
On the other hand, the use of self-stacking trays reduces the space required and facilitates its displacement by means of rolling bases. Likewise, the height of the tray towers can be adapted to the dimensions of the rearing chamber or the production level. The weight (5 kg) and dimensions of the trays (60 × 40 cm) allow for easy handling.
The proposed methodology allows for evaluating the efficiency of the SIT as part of an integrated vector control program, by achieving optimal sterile/wild male ratios in field trials complying with the standard quality parameters proposed by IAEA-WHO. However, a greater effort has to be made to automate the rearing, irradiation and sterile male release processes to enable scaling up to an operational level. For this purpose, the incorporation of automatic systems for feeding larvae and adults, sex sorting, methods of irradiation in adults and release with automatic terrestrial or aerial systems are expected to be implemented in the future.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}