
Repat33 Acts as a Downstream Component of Eicosanoid Signaling Pathway Mediating Immune Responses of Spodoptera exigua, a Lepidopteran Insect


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Immune Challenge
2.3. Chemicals
2.4. Larval Tissue Preparation
2.5. RNA Extraction and cDNA Preparation
2.6. Bioinformatics Analysis
2.7. RT-PCR and RT-qPCR
2.8. RNA Interference (RNAi)
2.9. Hemocyte-Spreading Behavior Analysis
2.10. Nodulation Assay
2.11. Expression Analysis of Antimicrobial Peptide (AMP) Genes
2.12. Data Analysis
3. Results
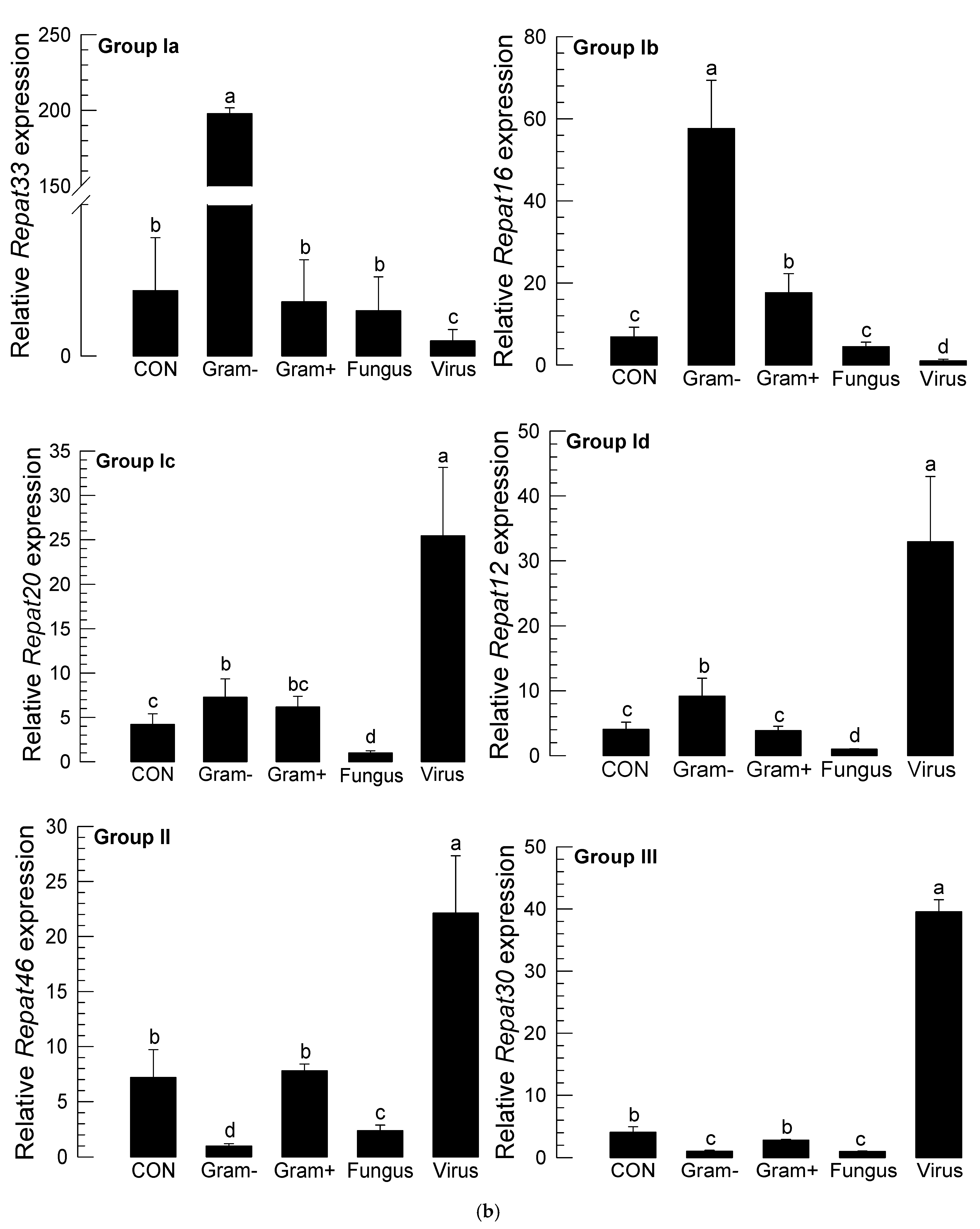
3.1. Repat Family Is Subdivided into Three Groups
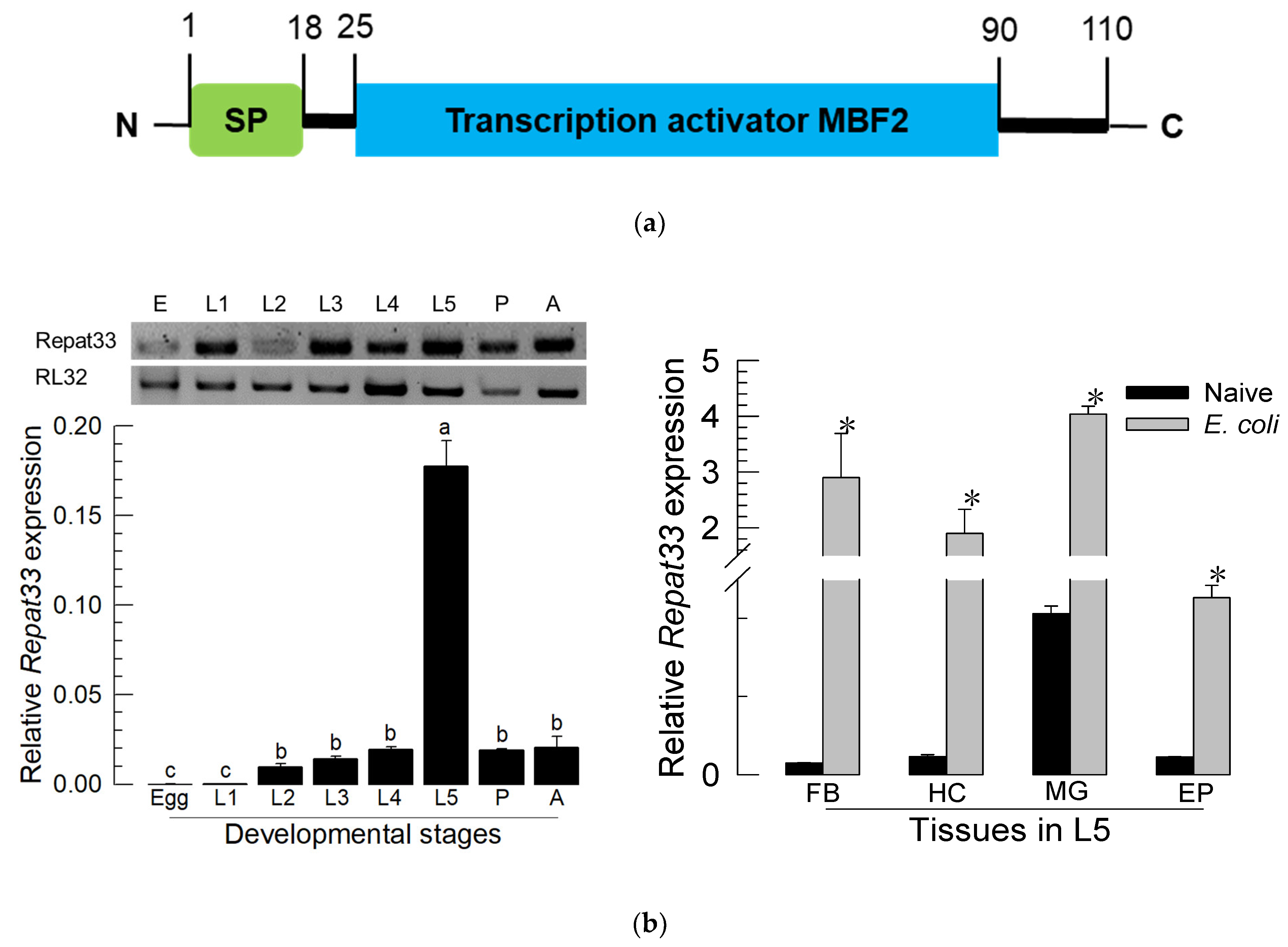
3.2. Expression Profile of Repat33
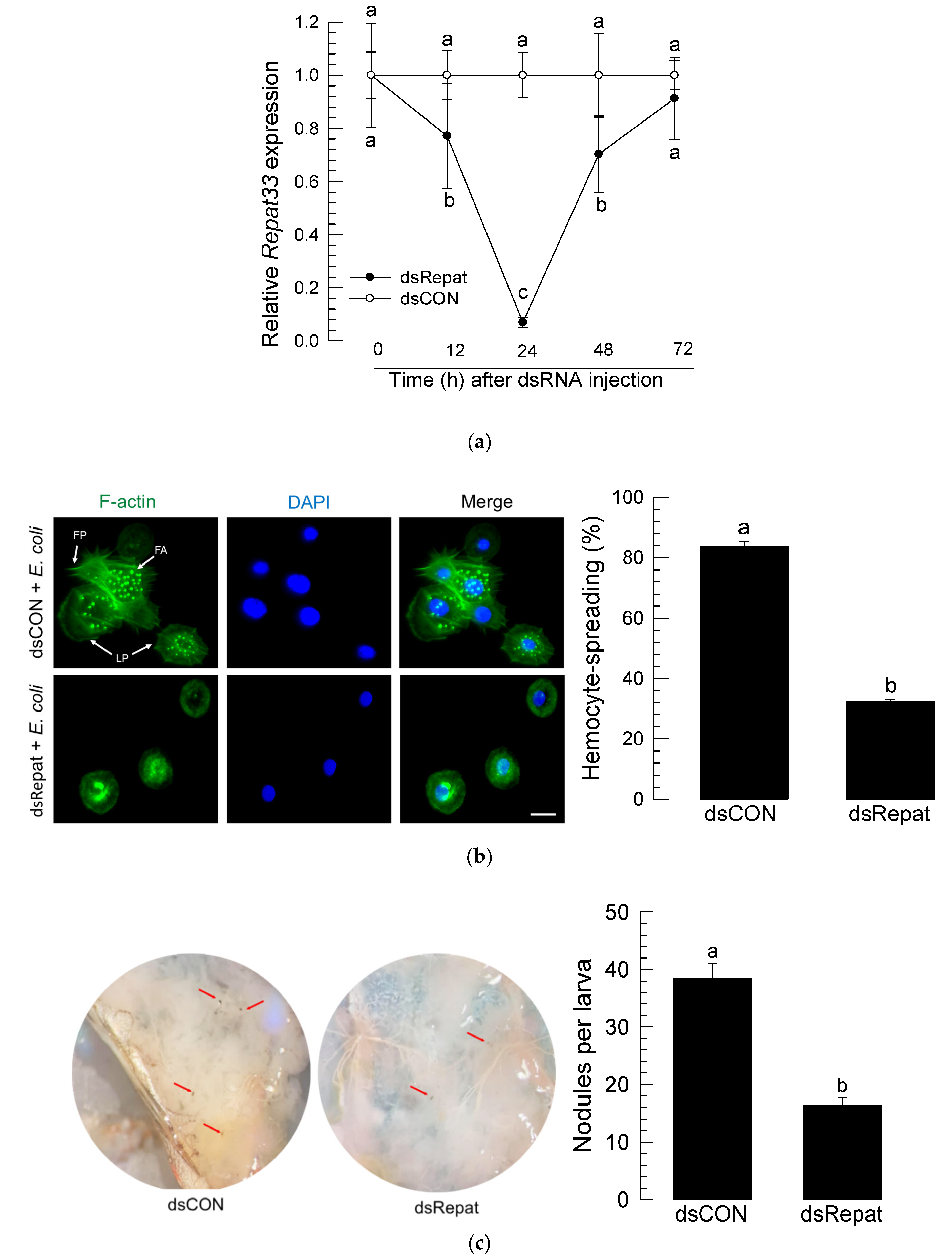
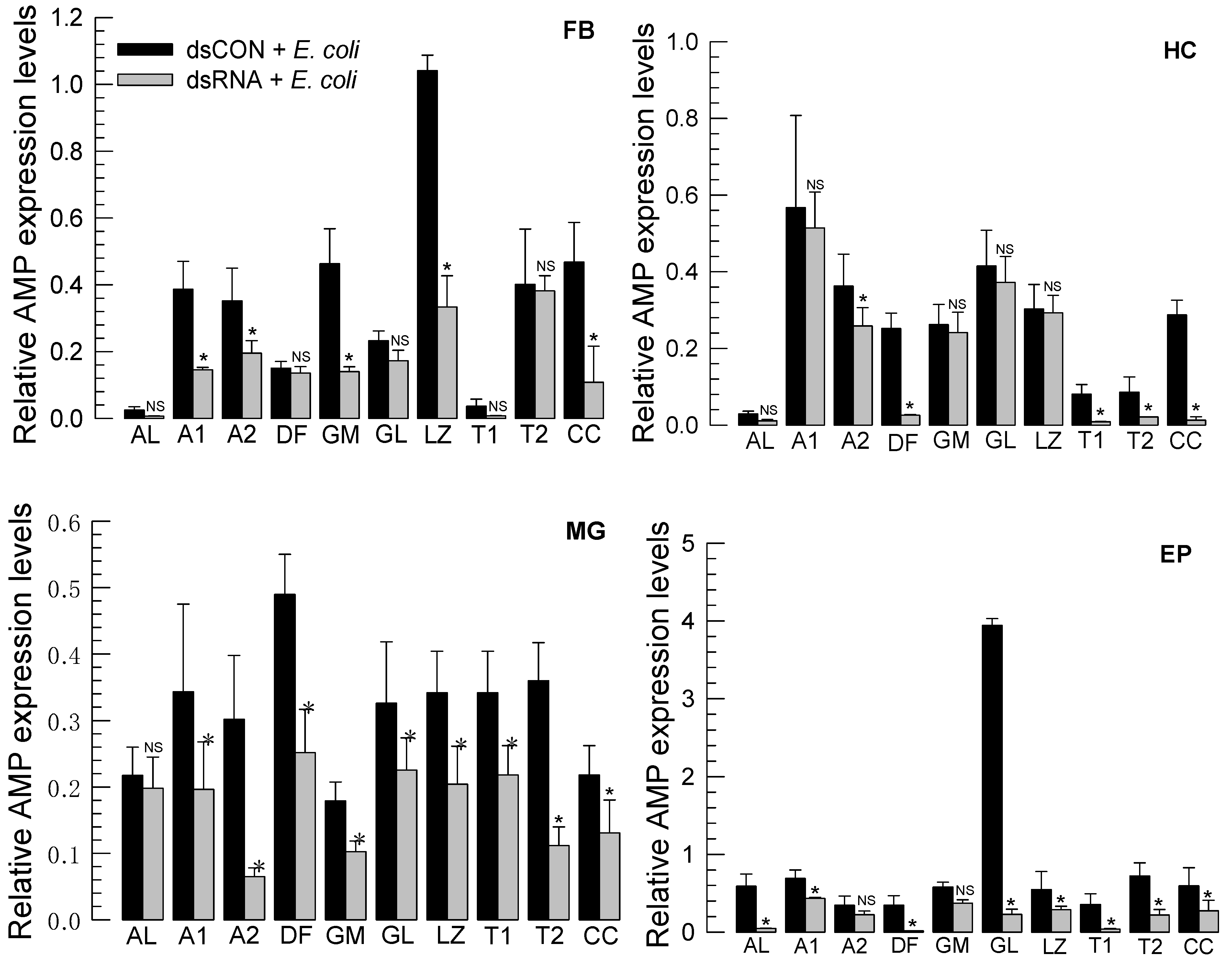
3.3. RNAi of Repat33 and Subsequent Influence on Immune Responses
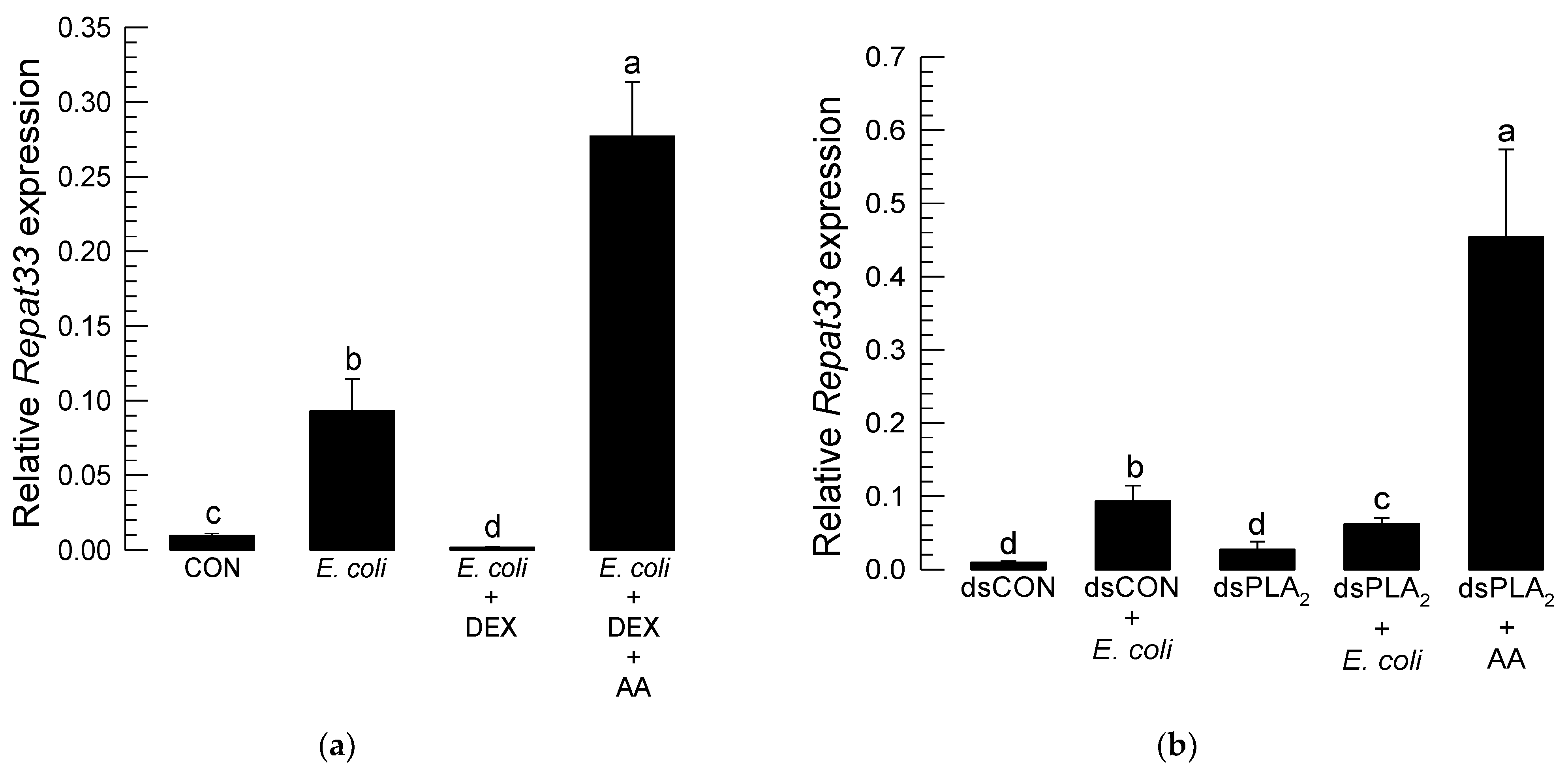
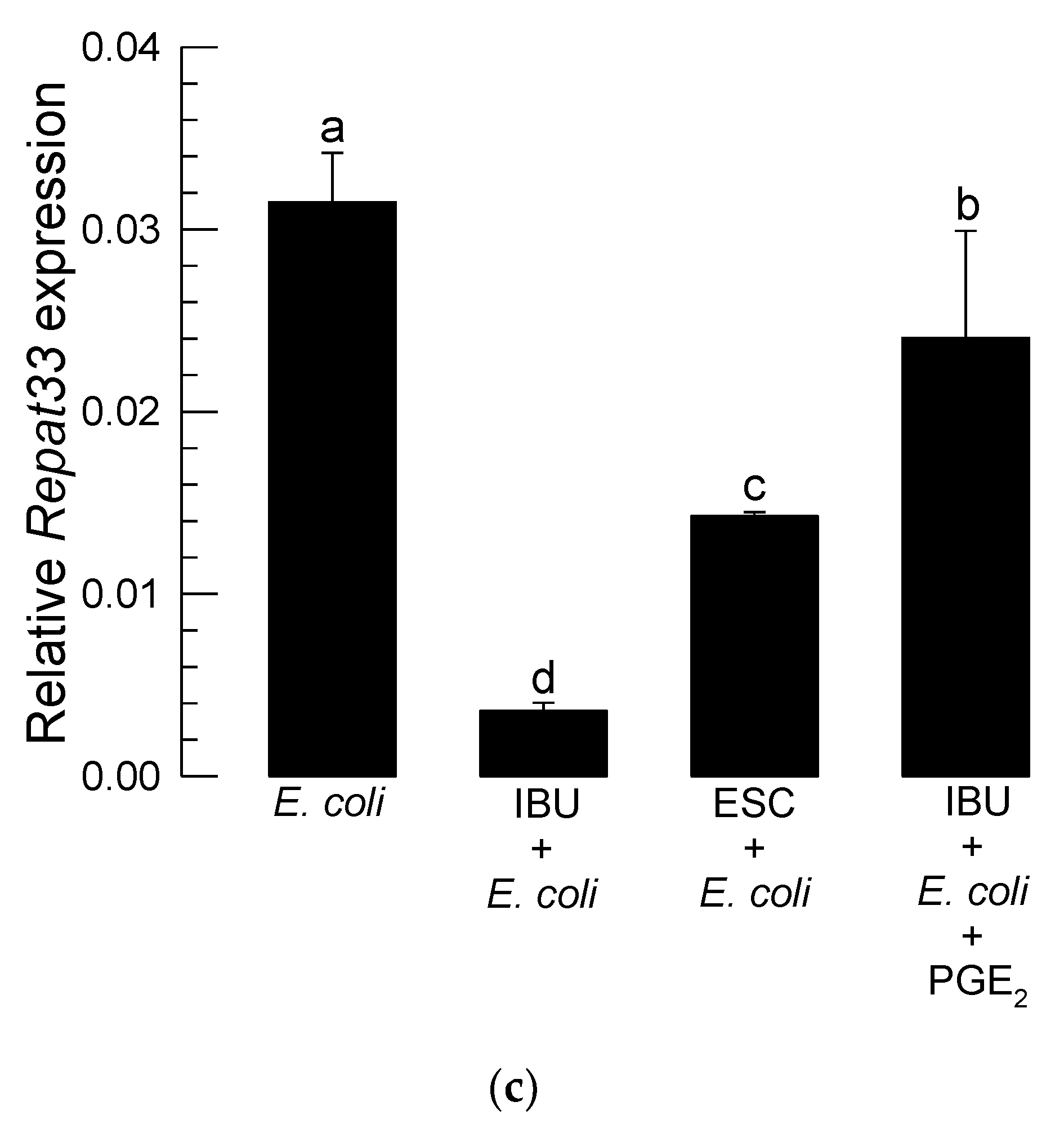
3.4. Repat33 Expression Is Controlled by Eicosanoids
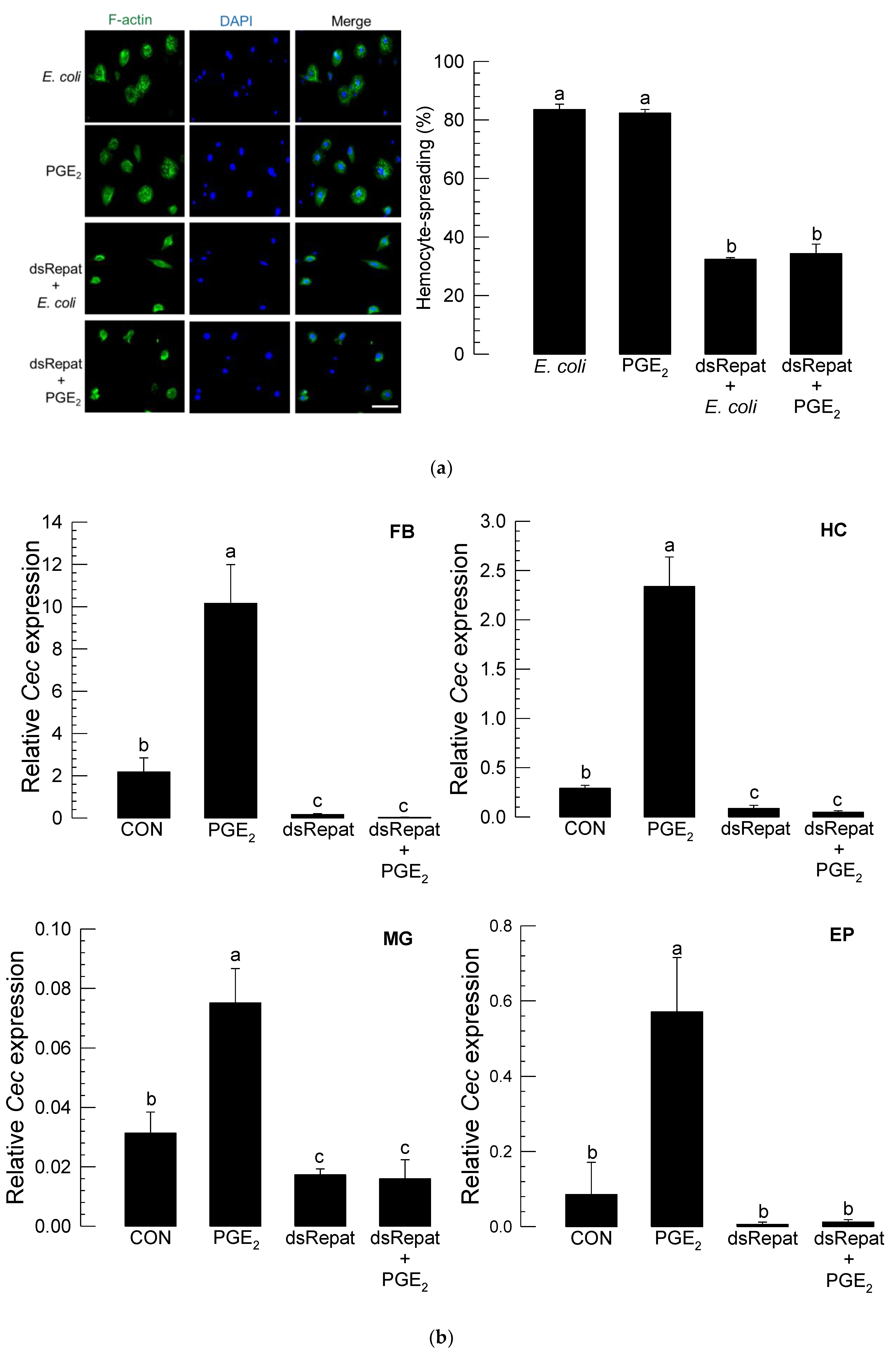
3.5. Repat33 Is a Downstream Component of PGE2 Signaling Pathway
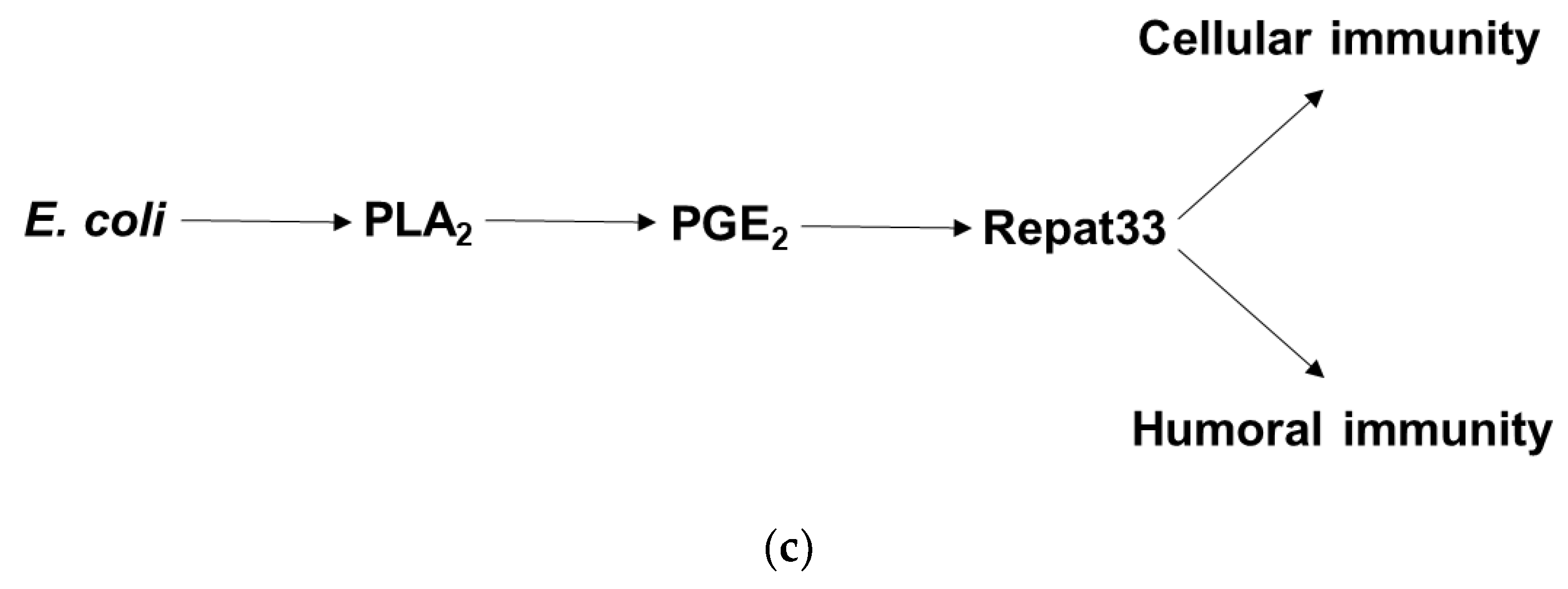
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Lemaitre, B.; Hoffmann, J. The host defense of Drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697–743. [Google Scholar] [CrossRef] [Green Version]
- Strand, M.R.; Clark, K.D. Plasmatocyte spreading peptide induces spreading of plasmatocytes but represses spreading of granulocytes. Arch. Insect Biochem. Physiol. 1999, 42, 213–223. [Google Scholar] [CrossRef]
- Janeway, C.A., Jr.; Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol. 2002, 20, 197–216. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Ahmed, S.; Stanley, D.; An, C. Eicosanoid-mediated immunity in insects. Dev. Comp. Immunol. 2018, 83, 130–143. [Google Scholar] [CrossRef]
- Lavine, M.D.; Strand, M.R. Insect hemocytes and their role in immunity. Insect Biochem. Mol. Biol. 2002, 32, 1295–1309. [Google Scholar] [CrossRef]
- Imler, J.L.; Bulet, P. Antimicrobial peptides in Drosophila: Structures, activities and gene regulation. Chem. Immunol. Allergy 2005, 86, 1–21. [Google Scholar]
- Herrero, S.; Ansems, M.; Van Oers, M.M.; Vlak, J.M.; Bakker, P.L.; de Maagd, R.A. REPAT, a new family of proteins induced by bacterial toxins and baculovirus infection in Spodoptera exigua. Insect Biochem. Mol. Biol. 2007, 37, 1109–1118. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Martínez, P.; Navarro-Cerrillo, G.; Caccia, S.; de Maagd, R.A.; Moar, W.J.; Ferré, J.; Escriche, B.; Herrero, S. Constitutive activation of the midgut response to Bacillus thuringiensis in Bt-resistant Spodoptera exigua. PLoS ONE 2010, 5, e12795. [Google Scholar] [CrossRef]
- Hernández-Rodríguez, C.S.; Ferré, J.; Herrero, S. Genomic structure and promoter analysis of pathogen-induced repat genes from Spodoptera exigua. Insect Mol. Biol. 2009, 18, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Cerrillo, G.; Ferré, J.; de Maagd, R.A.; Herrero, S. Functional interactions between members of the REPAT family of insect pathogen-induced proteins. Insect Mol. Biol. 2012, 21, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Stanley, D. Eicosanoids in Invertebrate Signal Transduction Systems; Princeton University Press: Princeton, NJ, USA, 2000. [Google Scholar]
- Dennis, E.A. The growing phospholipase A2 superfamily of signal transduction enzymes. Trends Biochem. Sci. 1997, 22, 1–2. [Google Scholar] [CrossRef]
- Park, Y.; Stanley, D.; Kim, Y. Eicosanoids up-regulate production of reactive oxygen species by NADPH-dependent oxidase in Spodoptera exigua phagocytic hemocytes. J. Insect Physiol. 2015, 79, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Sadekuzzaman, M.; Gautam, N.; Kim, Y. A novel calcium-independent phospholipase A2 and its physiological roles in development and immunity of a lepidopteran insect, Spodoptera exigua. Dev. Comp. Immunol. 2017, 77, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Vatanparast, M.; Ahmed, S.; Herrero, S.; Kim, Y. A non-venomous sPLA2 of a lepidopteran insect: Its physiological functions in development and immunity. Dev. Comp. Immunol. 2018, 89, 83–92. [Google Scholar] [CrossRef]
- Sajjadian, S.M.; Vatanparast, M.; Stanley, D.; Kim, Y. Secretion of secretory phospholipase A2 into Spodoptera exigua larval midgut lumen and its role in lipid digestion. Insect Mol. Biol. 2019, 28, 773–784. [Google Scholar] [CrossRef]
- Stanley, D.; Kim, Y. Why most insects have very low proportions of C20 polyunsaturated fatty acids: The oxidative stress hypothesis. Arch. Insect Biochem. Physiol. 2020, 103, e21622. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.A.; Ahmed, S.; Mollah, M.M.I.; Lee, D.; Kim, Y. Variation in pathogenicity of different strains of Xenorhabdus nematophila: Differential immunosuppressive activities and secondary metabolite production. J. Invertebr. Pathol. 2019, 166, 107221. [Google Scholar] [CrossRef]
- Ahmed, S.; Stanley, D.; Kim, Y. An insect prostaglandin E2 synthase acts in immunity and reproduction. Front. Physiol. 2018, 9, 1231. [Google Scholar] [CrossRef]
- Sajjadian, S.M.; Kim, Y. PGE2 upregulates gene expression of dual oxidase in a lepidopteran insect midgut via cAMP signalling pathway. Open Biol. 2020, 10, 200197. [Google Scholar] [CrossRef]
- Ahmed, S.; Al Baki, M.A.; Lee, J.; Seo, D.Y.; Lee, D.; Kim, Y. The first report of prostacyclin and its physiological roles in insects. Gen. Comp. Endocrinol. 2021, 301, 113659. [Google Scholar] [CrossRef]
- Al Baki, M.A.; Roy, M.C.; Lee, D.; Stanley, D.; Kim, Y. The prostanoids, thromboxanes, mediate hemocytic immunity to bacterial infection in the lepidopteran Spodoptera exigua. Dev. Comp. Immunol. 2021, 120, 104069. [Google Scholar] [CrossRef] [PubMed]
- Vatanparast, M.; Lee, D.H.; Kim, Y. Biosynthesis and immunity of epoxyeicosatrienoic acids in a lepidopteran insect, Spodoptera exigua. Dev. Comp. Immunol. 2020, 107, 103643. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, S.; Kim, Y. Various eicosanoids modulate the cellular and humoral immune responses of the beet armyworm, Spodoptera exigua. Biosci. Biotechnol. Biochem. 2009, 73, 2077–2084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, H.; Yang, Y.; Kumar, S.; Lee, D.W.; Bajracharya, P.; Calkins, T.L.; Kim, Y.; Pietrantonio, P.V. Characterization of the first insect prostaglandin (PGE2) receptor: MansePGE2R is expressed in oenocytoids and lipoteichoic acid (LTA) increases transcript expression. Insect Biochem. Mol. Biol. 2020, 117, 103290. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Ahmed, S.; Al Baki, M.A.; Kumar, S.; Kim, K.; Park, Y.; Stanley, D. Deletion mutant of PGE2 receptor using CRISPR-Cas9 exhibits larval immunosuppression and adult infertility in a lepidopteran insect, Spodoptera exigua. Dev. Comp. Immunol. 2020, 111, 103743. [Google Scholar] [CrossRef]
- Goh, H.G.; Lee, S.G.; Lee, B.P.; Choi, K.M.; Kim, J.H. Simple mass-rearing of beet armyworm, Spodoptera exigua (Hübner) (Lepidoptera: Noctuidae), on an artificial diet. Korean J. Appl. Entomol. 1990, 29, 180–183. [Google Scholar]
- Kim, H.; Keum, S.; Hasan, A.; Kim, H.; Jung, Y.; Lee, D.; Kim, Y. Identification of an entomopathogenic bacterium, Xenorhabdus ehlersii KSY, from Steinernema longicaudum GNUS101 and its immunosuppressive activity against insect host by inhibiting eicosanoid biosynthesis. J. Invertebr. Pathol. 2018, 159, 6–17. [Google Scholar] [CrossRef]
- Jung, S.; Kim, Y. An entomopathogenic bacterium, Xenorhabdus nematophila K1, enhances baculovirus pathogenicity against Spodoptera exigua and Plutella xylostella. J. Asia Pac. Entomol. 2006, 9, 139–143. [Google Scholar] [CrossRef]
- Park, J.; Kim, Y. Phospholipase A2 inhibitors in bacterial culture broth enhance pathogenicity of a fungus Nomuraea rileyi. J. Microbiol. 2012, 50, 644–651. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- SAS Institute, Inc. SAS/STAT User’s Guide; SAS Institute: Cary, NC, USA, 1989. [Google Scholar]
- Li, F.Q.; Takemaru, K.; Goto, M.; Ueda, H.; Handa, H.; Hirose, S. Transcriptional activation through interaction of MBF2 with TFIIA. Genes Cells 1997, 2, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.X.; Ueda, H.; Hirose, S. MBF2 is a tissue- and stage-specific coactivator that Is regulated at the step of nuclear transport in the silkworm Bombyx mori. Dev. Biol. 2000, 225, 437–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, S.; Kim, Y. PGE2 mediates cytoskeletal rearrangement of hemocytes via Cdc42, a small G protein, to activate actin-remodeling factors in Spodoptera exigua (Lepidoptera: Noctuidae). Arch. Insect Biochem. Physiol. 2019, 102, e21607. [Google Scholar] [CrossRef]
- Sajjadian, S.M.; Ahmed, S.; Al Baki, M.A.; Kim, Y. Prostaglandin D2 synthase and its functional association with immune and reproductive processes in a lepidopteran insect, Spodoptera exigua. Gen. Comp. Endocrinol. 2020, 287, 113352. [Google Scholar] [CrossRef] [PubMed]
- Morishima, I.; Yamano, Y.; Inoue, K.; Matsuo, N. Eicosanoids mediate induction of immune genes in the fat body of the silkworm, Bombyx mori. FEBS Lett. 1997, 419, 83–86. [Google Scholar] [CrossRef] [Green Version]
- Yajima, M.; Tanaka, M.; Tanahashi, N.; Kikuchi, H.; Natori, S.; Oshima, Y.; Kurata, S. A newly established in vitro culture using transgenic Drosophila reveals functional coupling between the phospholipase A2-generated fatty acid cascade and lipopolysaccharide-dependent activation of the immune deficiency (imd) pathway in insect immunity. Biochem. J. 2003, 371, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, S.; Kim, Y. Activation of immune-associated phospholipase A2 is functionally linked to Toll/Imd signal pathways in the red flour beetle, Tribolium castaneum. Dev. Comp. Immunol. 2010, 34, 530–537. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hrithik, M.T.H.; Vatanparast, M.; Ahmed, S.; Kim, Y. Repat33 Acts as a Downstream Component of Eicosanoid Signaling Pathway Mediating Immune Responses of Spodoptera exigua, a Lepidopteran Insect. Insects 2021, 12, 449. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12050449
Hrithik MTH, Vatanparast M, Ahmed S, Kim Y. Repat33 Acts as a Downstream Component of Eicosanoid Signaling Pathway Mediating Immune Responses of Spodoptera exigua, a Lepidopteran Insect. Insects. 2021; 12(5):449. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12050449
Chicago/Turabian StyleHrithik, Md Tafim Hossain, Mohammad Vatanparast, Shabbir Ahmed, and Yonggyun Kim. 2021. "Repat33 Acts as a Downstream Component of Eicosanoid Signaling Pathway Mediating Immune Responses of Spodoptera exigua, a Lepidopteran Insect" Insects 12, no. 5: 449. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12050449