
Molecular Identification of Trissolcus japonicus, Parasitoid of the Brown Marmorated Stink Bug, by Species-Specific PCR

 , , and
, , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Primer Design
2.2. Sample Collection and Processing
2.3. T. japonicus and BMSB Rearing Conditions
2.4. Genomic DNA Extractions
2.5. Polymerase Chain Reaction Conditions
2.6. Primer Specificity
2.7. Primer Sensitivity
2.8. Application to Parasitized Eggs
3. Results
3.1. Species-Specific Primer Design and Optimization of PCR Conditions
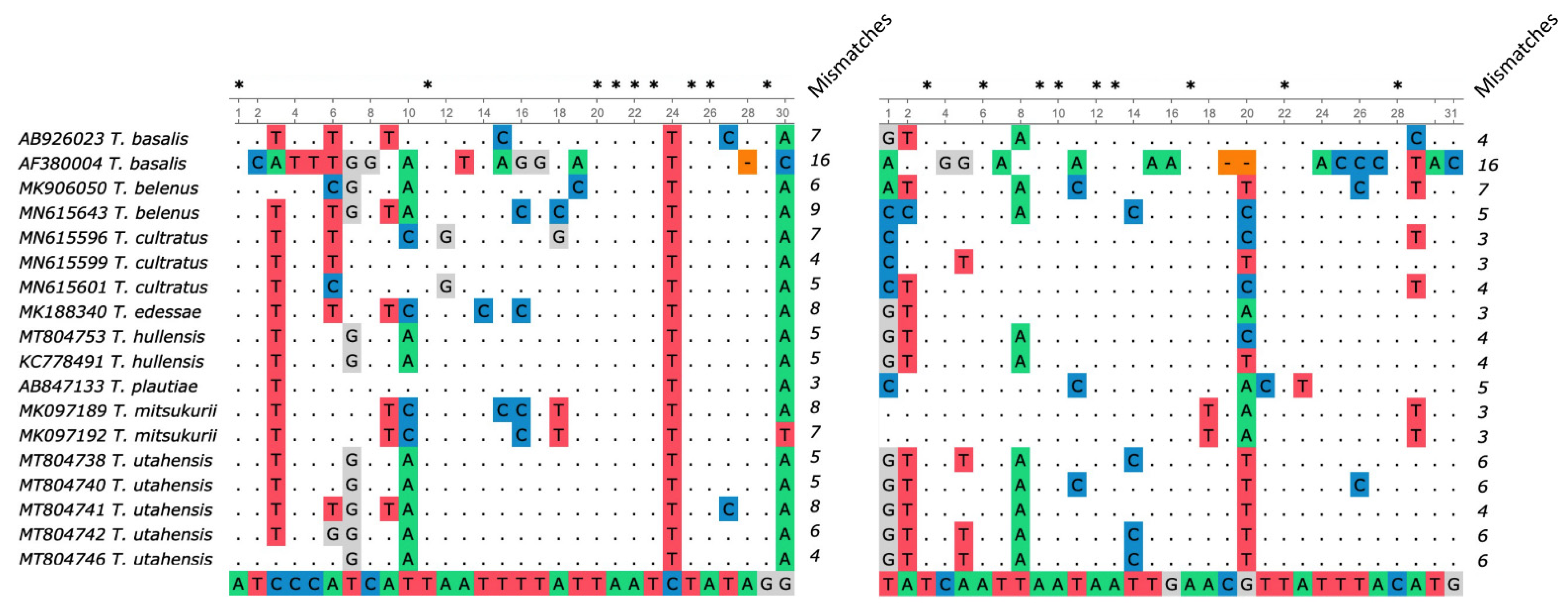
3.2. Primer Specificity
3.3. PCR Validation on Parasitized Eggs
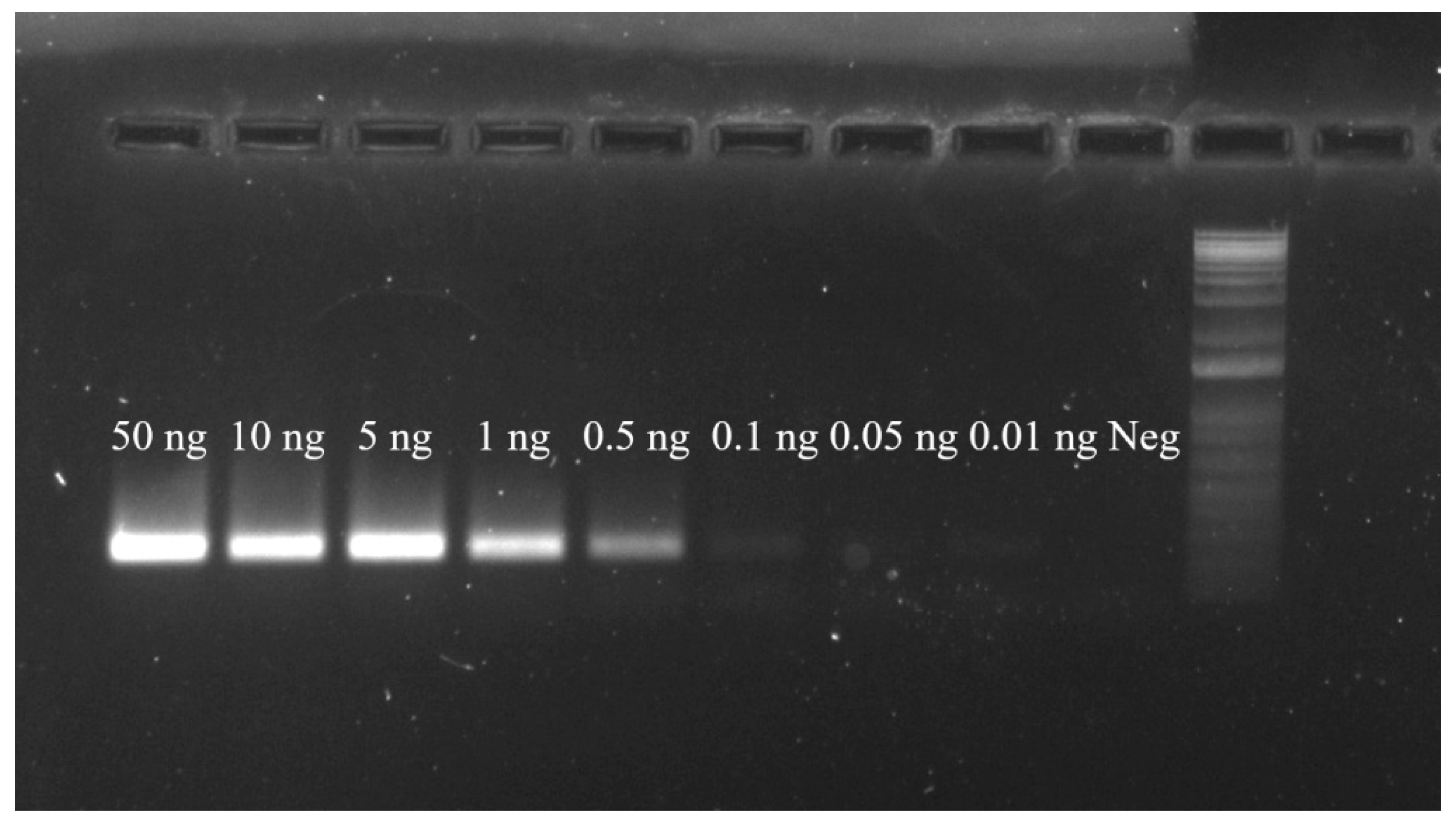
3.4. Primer Sensitivity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hoebeke, E.R.; Carter, M.E. Halyomorpha halys (Stål) (Heteroptera: Pentatomidae): A polyphagous plant pest from Asia newly detected in North America. Proc. Entomol. Soc. Wash. 2005, 105, 225–237. [Google Scholar]
- Fogain, R.; Graff, S. First records of the invasive pest, Halyomorpha halys (Hemiptera: Pentatomidae), in Ontario and Quebec. J. Entomol. Soc. Ont. 2011, 142, 45–48. [Google Scholar]
- Wermelinger, B.; Wyniger, D.; Forster, B. First records of an invasive bug in Europe: Halyomorpha halys Stål (Heteroptera: Pentatomidae), a new pest on woody ornamentals and fruit trees? Alp. Entomol. 2008, 81, 1–8. [Google Scholar]
- Maistrello, L.; Dioli, P.; Bariselli, M.; Mazzoli, G.L.; Giacalone-Forini, I. Citizen science and early detection of invasive species: Phenology of first occurrences of Halyomorpha halys in Southern Europe. Biol. Invasions 2016, 18, 3109–3116. [Google Scholar] [CrossRef]
- Nielsen, A.L.; Hamilton, G.C. Seasonal occurrence and impact of Halyomorpha halys (Hemiptera: Pentatomidae) in tree fruit. J. Econ. Entomol. 2009, 102, 1133–1140. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, E.; Bernhard, K.M.; Bernon, G.; Bickterton, M.; Gill, S.; Gonzales, C.; Hamilton, G.C.; Hedstrom, C.; Kamminga, K.; Koplinka-Loehr, C.; et al. Host Plants of the Brown Marmorated Stink Bug in the U.S. Northeastern IPM Center. Available online: https://www.stopbmsb.org/where-is-bmsb/host-plants/ (accessed on 19 April 2021).
- Leskey, T.C.; Nielsen, A.L. Impact of the invasive brown marmorated stink bug in north America and Europe: History, biology, ecology, and management. Annu. Rev. Entomol. 2018, 63, 599–618. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Bu, W.; Gao, Y.; Liu, G. Potential geographic distribution of brown marmorated stink bug invasion (Halyomorpha halys). PLoS ONE 2012, 7, e31246. [Google Scholar] [CrossRef]
- Haye, T.; Fischer, S.; Zhang, J.; Gariepy, T. Can native egg parasitoids adopt the invasive brown marmorated stink bug, Halyomorpha Halys (Heteroptera: Pentatomidae), in Europe? J. Pest. Sci. 2015, 88, 693–705. [Google Scholar] [CrossRef]
- Jones, A.L.; Jennings, D.E.; Hooks, C.R.R.; Shrewsbury, P.M. Field surveys of egg mortality and indigenous egg parasitoids of the brown marmorated stink bug, Halyomorpha Halys, in ornamental nurseries in the Mid-Atlantic region of the USA. J. Pest. Sci. 2017, 90, 1159–1168. [Google Scholar] [CrossRef]
- Dieckhoff, C.; Tatman, K.M.; Hoelmer, K.A. Natural biological control of Halyomorpha Halys by native egg parasitoids: A multi-year survey in Northern Delaware. J. Pest. Sci. 2017, 90, 1143–1158. [Google Scholar] [CrossRef]
- Yang, Z.-Q.; Yao, Y.-X.; Qiu, L.-F.; Li, Z.-X. A new species of Trissolcus (Hymenoptera: Scelionidae) parasitizing eggs of Halyomorpha Halys (Heteroptera: Pentatomidae) in China with comments on its biology. Ann. Entomol. Soc. 2009, 102, 39–47. [Google Scholar] [CrossRef]
- Lee, D.H.; Short, B.D.; Joseph, S.V.; Bergh, J.C.; Leskey, T.C. Review of the biology, ecology, and management of Halyomorpha Halys (Hemiptera: Pentatomidae) in China, Japan, and the Republic of Korea. Environ. Entomol. 2013, 42, 627–641. [Google Scholar] [CrossRef]
- Talamas, E.J.; Herlihy, M.V.; Dieckhoff, C.; Hoelmer, K.A.; Buffington, M.; Bon, M.-C.; Weber, D.C. Trissolcus japonicus (Ashmead) (Hymenoptera, Scelionidae) emerges in North America. J. Hymenopt. Res. 2015, 43, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Jentsch, P. Expanding the range of the samurai wasp, Trissolcus japonicus, in New York orchards. N. Y. Fruit Q. 2017, 25, 31–35. [Google Scholar]
- Herlihy, M.V.; Talamas, E.J.; Weber, D.C. Attack and success of native and exotic parasitoids on eggs of Halyomorpha halys in three Maryland habitats. PLoS ONE 2016, 11, e0150275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zhang, F.; Gariepy, T.; Mason, P.; Gillespie, D.; Talamas, E.; Haye, T. Seasonal parasitism and host specificity of Trissolcus japonicus in Northern China. J. Pest. Sci. 2017, 90, 1127–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedstrom, C.; Lowenstein, D.; Andrews, H.; Bai, B.; Wiman, N. Pentatomid host suitability and the discovery of introduced populations of Trissolcus japonicus in Oregon. J. Pest. Sci. 2017, 90, 1169–1179. [Google Scholar] [CrossRef]
- Charles, J.G.; Avila, G.A.; Hoelmer, K.A.; Hunt, S.; Gardner-Gee, R.; MacDonald, F.; Davis, V. Experimental assessment of the biosafety of Trissolcus japonicus in New Zealand, prior to the anticipated arrival of the invasive pest Halyomorpha halys. BioControl 2019, 64, 367–379. [Google Scholar] [CrossRef]
- Botch, P.S.; Delfosse, E.S. Host-acceptance behavior of Trissolcus japonicus (Hymenoptera: Scelionidae) Reared on the invasive Halyomorpha halys (Heteroptera: Pentatomidae) and nontarget species. Environ. Entomol. 2018, 47, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Jentsch, P.; Brown, L.; Canino, L.; Chen, M.; Jaeger, J.; Stewart, M.; Woelfersheim, A.; Kent, P. Results of 2020 Insecticide and Acaricide Studies in Eastern New York; Report for Hudson Valley Laboratory; Hudson Valley Laboratory: Highland, NY, USA, 2020. [Google Scholar]
- Venerate® XC Bioinsecticide Improves Insect Control. Available online: https://marronebio.com/products/venerate/ (accessed on 22 March 2021).
- Venerate® XC. Available online: http://www.cdms.net/LabelsSDS/home/ (accessed on 29 March 2021).
- Rutgers University New Jersey Agricultural Experiment Center. Pesticide Control for the Brown Marmorated Stink Bug. Available online: https://njaes.rutgers.edu/stink-bug/pesticides.php (accessed on 29 March 2021).
- National Pesticide Information Center. Bifenthrin. Available online: http://npic.orst.edu/factsheets/bifgen.html (accessed on 22 March 2021).
- National Pesticide Information Center. Cyfluthrin. Available online: http://www.npic.orst.edu/factsheets/cyfluthringen.html (accessed on 29 March 2021).
- Drape Net Pty Ltd. Drape Net®. Available online: https://drapenet.com/drape-net/# (accessed on 22 March 2021).
- Baker, B.P.; Green, T.A.; Loker, A.J. Biological control and integrated pest management in organic and conventional systems. Biol. Control 2020, 140, 104095. [Google Scholar] [CrossRef]
- Mandal, S.D.; Chhakchhuak, L.; Gurusubramanian, G.; Kumar, N.S. Mitochondrial markers for identification and phylogenetic studies in insects—A review. DNA Barcodes 2014, 2, 1–9. [Google Scholar] [CrossRef]
- Talamas, E.J.; Bon, M.-C.; Hoelmer, K.A.; Buffington, M.L. Molecular phylogeny of Trissolcus wasps (Hymenoptera, Scelionidae) associated with Halyomorpha halys (Hemiptera, Pentatomidae). J. Hymenopt. Res. 2019, 73, 201–217. [Google Scholar] [CrossRef]
- Jentsch, P.; Agnello, A.; Tee, E.; Aćimović, D.; Brown, L. Biological Control of Brown Marmorated Stink Bug, Halyomorpha Halys Stål (Hemiptera: Pentatomidae) in NYS; Report for Hudson Valley Laboratory; Hudson Valley Laboratory: Highland, NY, USA, 2020. [Google Scholar]
- Kwok, S.; Kellogg, D.E.; McKinney, N.; Spasic, D.; Goda, L.; Levenson, C.; Sninsky, J.J. Effects of primer-template mismatches on the polymerase chain reaction: Human immunodeficiency virus type 1 model studies. Nucleic Acids Res. 1990, 18, 999–1005. [Google Scholar] [CrossRef]
- Simsek, M.; Adnan, H. Effect of single mismatches at 3′–End of primers on polymerase chain reaction. J. Sci. Res. Med. Sci. 2000, 2, 11–14. [Google Scholar]
- Lefever, S.; Pattyn, F.; Hellemans, J.; Vandesompele, J. Single-nucleotide polymorphisms and other mismatches reduce performance of quantitative PCR assays. Clin. Chem. 2013, 59, 1470–1480. [Google Scholar] [CrossRef] [Green Version]
- Bon, M.C.; United States Department of Agriculture-ARS, European Biological Control Laboratory, Montferrier-sur-Lez, France. Personal communication, March 2021.
- Okonechnikov, K.; Golosova, O.; Fursov, M.; UGENE team. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. Bioinformatics 2012, 13, 134. [Google Scholar] [CrossRef] [Green Version]
- Talamas, E.J.; Johnson, N.F.; Buffington, M. Key to nearctic species of Trissolcus Ashmead (Hymenoptera, Scelionidae), natural enemies of native and invasive stink bugs (Hemiptera, Pentatomidae). J. Hymenopt. Res. 2015, 43, 45–110. [Google Scholar] [CrossRef] [Green Version]
- Borror, D.J.; Johnson, N.F.; Triplehorn, C.A. An Introduction to the Study of Insects; Saunders College Pub.: Philadelphia, PA, USA, 1989. [Google Scholar]
- Stahl, J.M.; Gariepy, T.D.; Beukeboom, L.W.; Haye, T. A Molecular tool to identify anastatus parasitoids of the brown marmorated stink bug. Entomol. Exp. Appl. 2019, 167, 692–700. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, H.; Lowenstein, D.M.; Wiman, N.G.; Wong, J.S.; Lee, J.C. Parasitism of frozen Halyomorpha halys eggs by Trissolcus japonicus: Applications for rearing and experimentation. Biocontrol Sci. Technol. 2019, 29, 478–493. [Google Scholar] [CrossRef]
- Taekul, C.; Valerio, A.A.; Austin, A.D.; Klompen, H.; Johnson, N.F. Molecular phylogeny of Telenomine egg parasitoids (Hymenoptera: Platygastridae s.l.: Telenominae): Evolution of host shifts and implications for classification. Syst. Entomol. 2014, 39, 24–35. [Google Scholar] [CrossRef] [Green Version]
- Sabbatini-Peverieri, G.; Talamas, E.; Bon, M.-C.; Marianelli, L.; Bernardinelli, I.; Malossini, G.; Benvenuto, L.; Roversi, P.F.; Hoelmer, K. Two asian egg parasitoids of Halyomorpha Halys (Stål) (Hemiptera, Pentatomidae) emerge in Northern Italy: Trissolcus Mitsukurii (Ashmead) and Trissolcus japonicus (Ashmead) (Hymenoptera, Scelionidae). J. Hymenopt. Res 2018, 67, 37–53. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.B.; Hoeh, W.; Lutz, R.; Vrijenhoek, R.C. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit i from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Lara, J.; Pickett, C.; Ingels, C.; Haviland, D.R.; Grafton-Cardwell, E.; Doll, D.; Bethke, J.; Faber, B.; Dara, S.K.; Hoddle, M. Biological control program is being developed for brown marmorated stink bug. Calif. Agric. 2016, 70, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Gariepy, T.D.; Haye, T.; Zhang, J. A molecular diagnostic tool for the preliminary assessment of host–parasitoid associations in biological control programmes for a new invasive pest. Mol. Ecol. 2014, 23, 3912–3924. [Google Scholar] [CrossRef]
- Summy, K.R.; Gilstrap, F.E.; Hart, W.G.; Caballero, J.M.; Saenz, I. Biological control of citrus blackfly (Homoptera: Aleyrodidae) in Texas. Environ. Entomol. 1983, 12, 782–786. [Google Scholar] [CrossRef]
- Machtinger, E.T.; Geden, C.J.; Leppla, N.C. Linear dispersal of the filth fly parasitoid Spalangia cameroni (Hymenoptera: Pteromalidae) and parasitism of hosts at increasing distances. PLoS ONE 2015, 10, e0129105. [Google Scholar] [CrossRef]
- Huber, J.T.; Read, J.D.; Triapitsyn, S.V. Illustrated key to genera and catalogue of Mymaridae (Hymenoptera) in America North of Mexico. Zootaxa 2020, 4773, 1–411. [Google Scholar] [CrossRef]
- Trjapitzin, V.A. A review of encyrtid wasps (Hymenoptera, Chalcidoidea, Encyrtidae) of macaronesia. Entomol. Rev. 2008, 88, 218–232. [Google Scholar] [CrossRef]
- Kosheleva, O.V.; Kostjukov, V.V. Types of Parasitism in Eulophid Wasps (Hymenoptera, Eulophidae). Entmol. Rev. 2014, 94, 1202–1217. [Google Scholar] [CrossRef]
- Krzyżyński, M.; Ulrich, W. Ceraphronidae and Megaspilidae (Hymenoptera: Ceraphronoidea) of Poland: Current state of knowledge with corrections to the Polish checklist. Pol. J. Entomol. 2015, 84, 191–200. [Google Scholar] [CrossRef]
- Ronquist, F.; Nieves-Aldrey, J.-L.; Buffington, M.L.; Liu, Z.; Liljeblad, J.; Nylander, J.A.A. Phylogeny, evolution and classification of gall wasps: The plot thickens. PLoS ONE 2015, 10, e0123301. [Google Scholar] [CrossRef]
- Matsuo, K.; Hirose, Y.; Johnson, N.F. A Taxonomic issue of two species of Trissolcus (Hymenoptera: Platygastridae) Parasitic on eggs of the brown-winged green bug, Plautia stali (Hemiptera: Pentatomidae): Resurrection of T. plautiae, a cryptic species of T. japonicus revealed by morphology, reproductive isolation and molecular evidence. Appl. Entomol. Zool. 2014, 49, 385–394. [Google Scholar]
- Matsuo, K.; Honda, T.; Itoyama, K.; Toyama, M.; Hirose, Y. Discovery of three egg parasitoid species attacking the shield bug Glaucias subpunctatus (Hemiptera: Pentatomidae). Jpn. J. Appl. Entomol. Zool. 2016, 60, 43–45. [Google Scholar] [CrossRef] [Green Version]
- Stahl, J.; Tortorici, F.; Pontini, M.; Bon, M.-C.; Hoelmer, K.; Marazzi, C.; Tavella, L.; Haye, T. First discovery of adventive populations of Trissolcus japonicus in Europe. J. Pest. Sci. 2019, 92, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Leigh, J.W.; Bryant, D. PopART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Moraglio, S.T.; Tortorici, F.; Pansa, M.G.; Castelli, G.; Pontini, M.; Scovero, S.; Visentin, S.; Tavella, L. A 3-year survey on parasitism of Halyomorpha halys by egg parasitoids in Northern Italy. J. Pest. Sci. 2020, 93, 183–194. [Google Scholar] [CrossRef]
- Gariepy, T.D.; Bruin, A.; Konopka, J.; Scott-Dupree, C.; Fraser, H.; Bon, M.-C.; Talamas, E. A modified DNA barcode approach to define trophic interactions between native and exotic pentatomids and their parasitoids. Mol. Ecol. 2019, 28, 456–470. [Google Scholar] [CrossRef]
- Paparini, A.; Gofton, A.; Yang, R.; White, N.; Bunce, M.; Ryan, U.M. Comparison of sanger and next generation sequencing performance for genotyping cryptosporidium isolates at the 18S RRNA and actin loci. Exp. Parasitol. 2015, 151–152, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Schlatter, R.; Matte, U.; Polanczyk, C.; Koehler-Santos, P.; Ashton-Prolla, P. Costs of genetic testing: Supporting Brazilian public policies for the incorporating of molecular diagnostic technologies. Genet. Mol. 2015, 38, 332–337. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Specimen ID | Family | Accession # |
|---|---|---|
| Trissolcus japonicus voucher TJ272 | Scelionidae | MK188350 |
| Trissolcus japonicus voucher TJ262 | Scelionidae | MK188360 |
| Trissolcus japonicus voucher Scel-0945 | Scelionidae | MK188349 |
| Trissolcus euschisti voucher Scel-0548 | Scelionidae | MK188347 |
| Trissolcus euschisti voucher FEMS015-09 | Scelionidae | MG939533 |
| Telenomus podisi isolate OH23 | Scelionidae | KR870966 |
| Trissolcus plautiae isolate: 121219-21 | Scelionidae | AB847133 |
| Trissolcus basalis voucher TSP228 | Scelionidae | MK188338 |
| Trissolcus kozlovi isolate T4 | Scelionidae | MH521283 |
| Trissolcus corai voucher USNM:ENT:01223976 | Scelionidae | MN615613 |
| Trissolcus cultratus voucher USNM:ENT:00977540 | Scelionidae | MN615602 |
| Telenomus podisi voucher BIOUG10919-B03 | Scelionidae | KR374962 |
| Telenomus sp. BIOUG24593-G02 | Scelionidae | MG498324 |
| Platygastridae sp. BOLD:AAZ4474 voucher BIOUG03482-G06 | Platygastridae | KM556113 |
| Mymaridae sp. INDOBIOSYS-CCDB25313-E11 | Mymaridae | MH926817 |
| Ceraphronidae sp. BIOUG32185-E11 | Ceraphronidae | MG442991 |
| Pteromalidae sp. BOLD:ADS5523 voucher CHARS00074-B06 | Pteromalidae | MN669960 |
| Encyrtidae sp. BIOUG32169-A04 | Encyrtidae | MG447654 |
| Cynipidae sp. BIOUG23888-C07 | Cynipidae | MG442460 |
| Species | n | Accession # |
|---|---|---|
| Trissolcus basalis | 5 | MK188338 AB926023–AB926024 MN615660 AF380004 |
| Trissolcus belenus | 8 | MN615643–MN615644 MN603802–MN603806 MK906050 |
| Trissolcus cultratus | 8 | MN615596–MN615603 |
| Trissolcus edessae | 2 | MK188340 MN615575 |
| Trissolcus hullensis | 4 | MT804753–MT804755 KC778491 |
| Trissolcus plautiae | 23 | MN615614–MN615623 AB908183–AB908186 AB847147 AB847138–AB847142 AB847133–AB847135 |
| Trissolcus mitsukurii | 25 | MK097189–MK097202 MN615586–MN615595 AB971831 |
| Trissolcus utehensis | 15 | MT804738–MT804752 |
| n a | Sample Type and Specimen Identification b | Location | Collection Year | Control PCR c | Tj Specific PCR d |
|---|---|---|---|---|---|
| 29 | Field-collected insects | ||||
| Trissolcus japonicus | Poughkeepsie, NY | 2019 | + | + | |
| Trissolcus japonicus | Poughkeepsie, NY | 2019 | + | + | |
| Trissolcus japonicus | Campbell Hall, NY | 2018 | + | + | |
| Trissolcus japonicus | Campbell Hall, NY | 2018 | + | + | |
| Trissolcus japonicus | Campbell Hall, NY | 2018 | + | + | |
| Trissolcus japonicus | Campbell Hall, NY | 2018 | + | + | |
| Trissolcus japonicus | Campbell Hall, NY | 2018 | + | + | |
| Trissolcus euschisti | Monroe County, NY | 2018 | + | − | |
| Trissolcus euschisti | Highland, NY | 2020 | + | − | |
| Trissolcus brochymenae | Huron, NY | 2018 | + | − | |
| Trissolcus brochymenae | Poughkeepsie, NY | 2019 | + | − | |
| Telenomus podisi | New Paltz, NY | 2018 | + | − | |
| Telenomus podisi | Milton, NY | 2019 | + | − | |
| Telenomus sp. | Campbell Hall, NY | 2020 | + | − | |
| Telenomus sp. | Poughkeepsie, NY | 2018 | + | − | |
| Amitus sp. | New Paltz, NY | 2019 | + | − | |
| Amitus sp. | Monroe County, NY | 2019 | + | − | |
| Pteromalidae sp. | Marlboro, NY | 2019 | + | − | |
| Pteromalidae sp. | Warwick, NY | 2020 | + | − | |
| Pteromalidae sp. or Myrmaridae sp. e | Walden, NY | 2019 | + | − | |
| Encyrtidae sp. | Monroe County, NY | 2019 | + | − | |
| Encyrtidae sp. | Campbell Hall, NY | 2018 | + | − | |
| Eulophidae sp. | Monroe County, NY | 2019 | + | − | |
| Eulophidae sp. | Monroe County, NY | 2019 | + | − | |
| Eulophidae sp. | Huron, NY | 2018 | + | − | |
| Ceraphronidae sp. | New Paltz, NY | 2018 | + | − | |
| Ceraphronidae sp. | Holley, NY | 2018 | + | − | |
| Cynipidae sp. | New Paltz, NY | 2019 | + | − | |
| Cynipidae sp. | Williamson, NY | 2019 | + | − | |
| 13 | Laboratory-reared specimens | ||||
| Trissolcus japonicus | HVRL | 2020 | + | + | |
| 2 | Trissolcus japonicus | HVRL | 2020 | + | + |
| 8 | Halyomorpha halys parasitized eggs f | HVRL | 2020 | + | + |
| 3 | Halyomorpha halys g | HVRL | 2020 | − | − |
| Geographic Origin | Haplotype a | Representative Sequences b | n c | Geographic Origin | Haplotype a | Representative Sequences b | n c |
|---|---|---|---|---|---|---|---|
| Europe | 11 | Asia | 56 | ||||
| Switzerland | H1 | MH919753–MH919758 | China | ||||
| Italy | H1 *, H3 *, H4 * | MK097184–MK097188 | H2 | MH919759 | |||
| North America | 36 | H2 * | MK188348 | ||||
| Canada | H2 * | MK188349 | MK188354–MK188355 | ||||
| MW97094 | MK188358–MK188359 | ||||||
| US | H2 * | MK188361–MK188362 | |||||
| California | MW97096 | MN615624–MN615625 | |||||
| District of Columbia | MW97073 | H7 * | MK188357 | ||||
| Delaware | MW97079 | H11 # | MK188363 | ||||
| Maryland | MW97074 | MN615630 | |||||
| MW97076 | H12 # | MK188353 | |||||
| MW97078 | MN615632 | ||||||
| MW97093 | Japan | ||||||
| New Jersey | MW97085–MW97086 | H1 | MH919743 | ||||
| MW97089 | H1 *, H3 *, H4 * | MK188351 | |||||
| MW97091 | MK188356 | ||||||
| New York | MW97082 | MN615628 | |||||
| MK188350 | H3 | MH919744–MH919752 | |||||
| MW97084 | H4 | AB847131–AB847132 | |||||
| Ohio | MW97097–MW97101 | AB847136 | |||||
| MW97103 | H5 | AB908179–AB908182 | |||||
| Oregon | MW97077 | AB847144–AB847145 | |||||
| MW97087 | AB894834–AB894835 | ||||||
| MK188360 | AB894838–AB894839 | ||||||
| Pennsylvania | MW97092 | H5 * | MN615626 | ||||
| Utah | MW97102 | H6 | AB847129–AB847130 | ||||
| Virginia | MW97071–MW97072 | AB847137 | |||||
| MW97088 | AB847143 | ||||||
| Washington | MW97075 | AB847146 | |||||
| MW97083 | AB894836–AB894837 | ||||||
| MW97090 | AB894840–AB894841 | ||||||
| MW97095 | H8 # | MN615627 | |||||
| West Virginia | MW97080–MW97081 | H9 # | MN615633 | ||||
| H10 # | MN615631 | ||||||
| South Korea | |||||||
| H2 * | MK188352 | ||||||
| H7 # | MN615629 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, M.N.; Santander, R.D.; Talamas, E.J.; Jentsch, P.J.; Bon, M.-C.; Aćimović, S.G. Molecular Identification of Trissolcus japonicus, Parasitoid of the Brown Marmorated Stink Bug, by Species-Specific PCR. Insects 2021, 12, 467. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12050467
Chen MN, Santander RD, Talamas EJ, Jentsch PJ, Bon M-C, Aćimović SG. Molecular Identification of Trissolcus japonicus, Parasitoid of the Brown Marmorated Stink Bug, by Species-Specific PCR. Insects. 2021; 12(5):467. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12050467
Chicago/Turabian StyleChen, Maple N., Ricardo D. Santander, Elijah J. Talamas, Peter J. Jentsch, Marie-Claude Bon, and Srđan G. Aćimović. 2021. "Molecular Identification of Trissolcus japonicus, Parasitoid of the Brown Marmorated Stink Bug, by Species-Specific PCR" Insects 12, no. 5: 467. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12050467