
Development of Monitoring and Mating Disruption against the Chilean Leafroller Proeulia auraria (Lepidoptera: Tortricidae) in Orchards

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Orchards
2.2. Chemicals, Traps, and Pheromone Matrix
2.3. Optimizing Pheromone Load in Septa
2.4. Remaining Pheromone in Septa after Aging under Field Conditions
2.5. Proeulia Auraria Adult Male Phenology
2.6. Proeulia Auraria Mating Disruption Trials
2.7. Remaining Pheromone in SPLAT Matrix after Ageing under Field Conditions
3. Results
3.1. Optimizing Pheromone Load in Septa
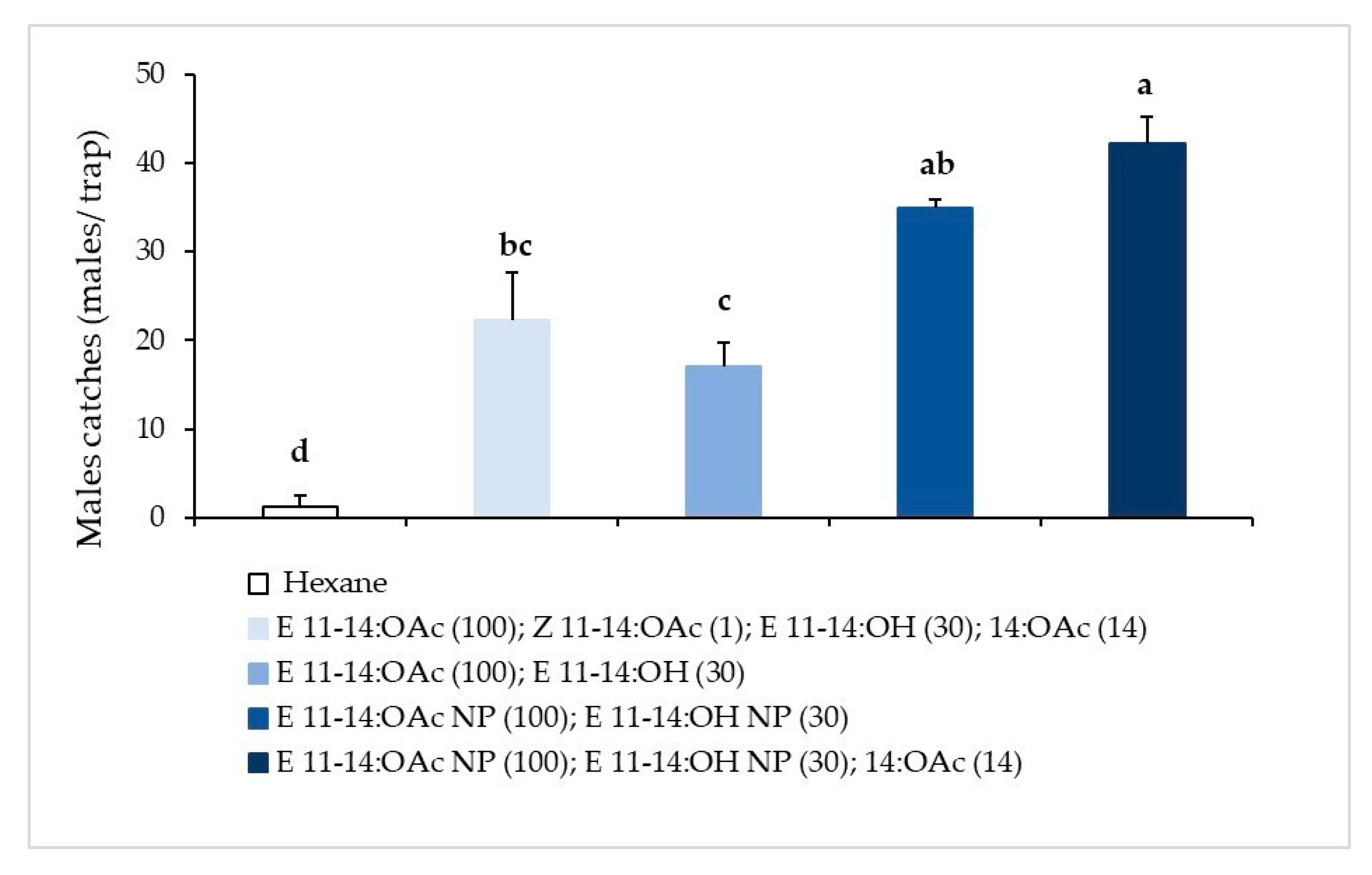
3.1.1. Pheromone Composition Trial
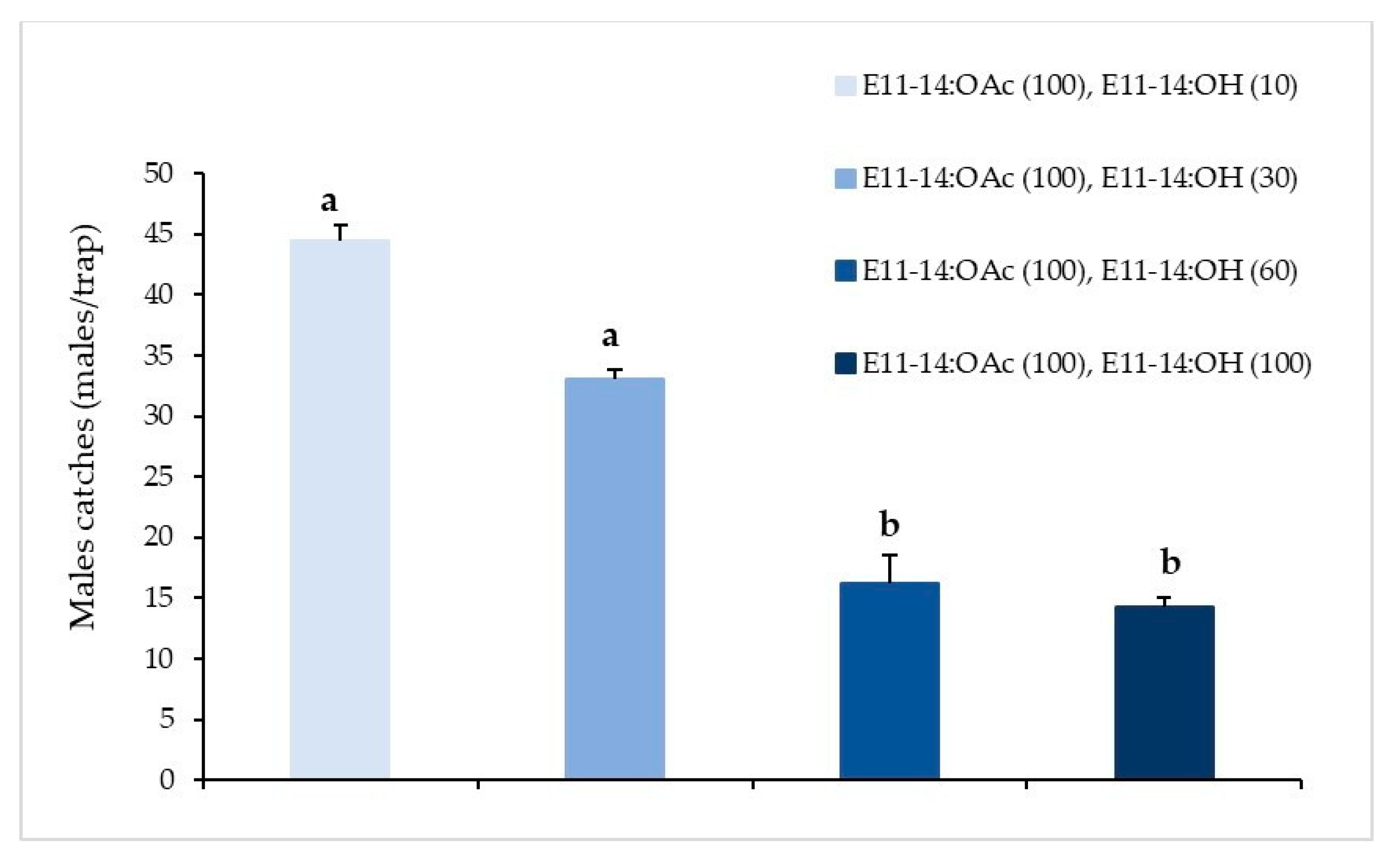
3.1.2. Pheromone Proportion Trial
3.1.3. Septa Longevity
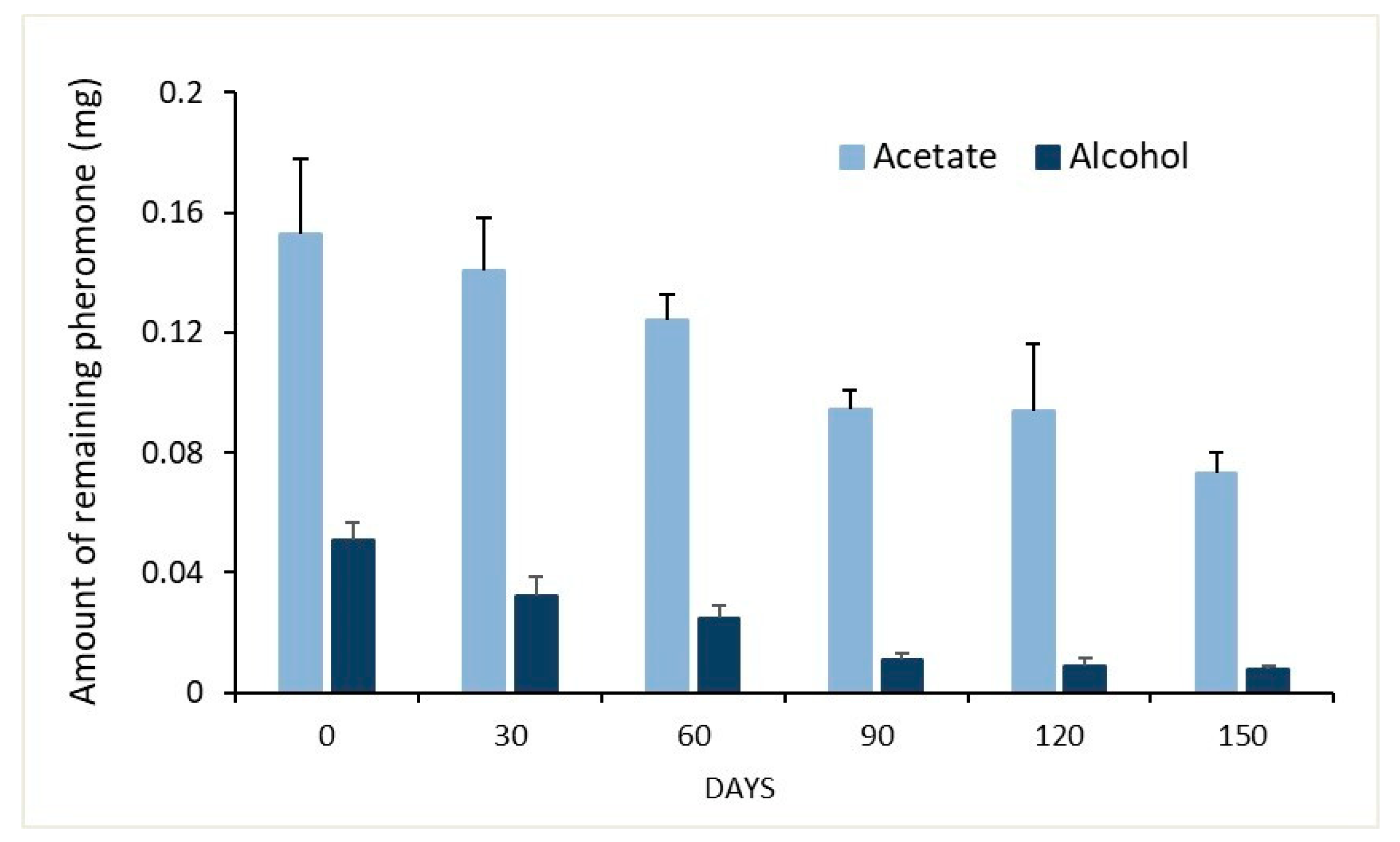
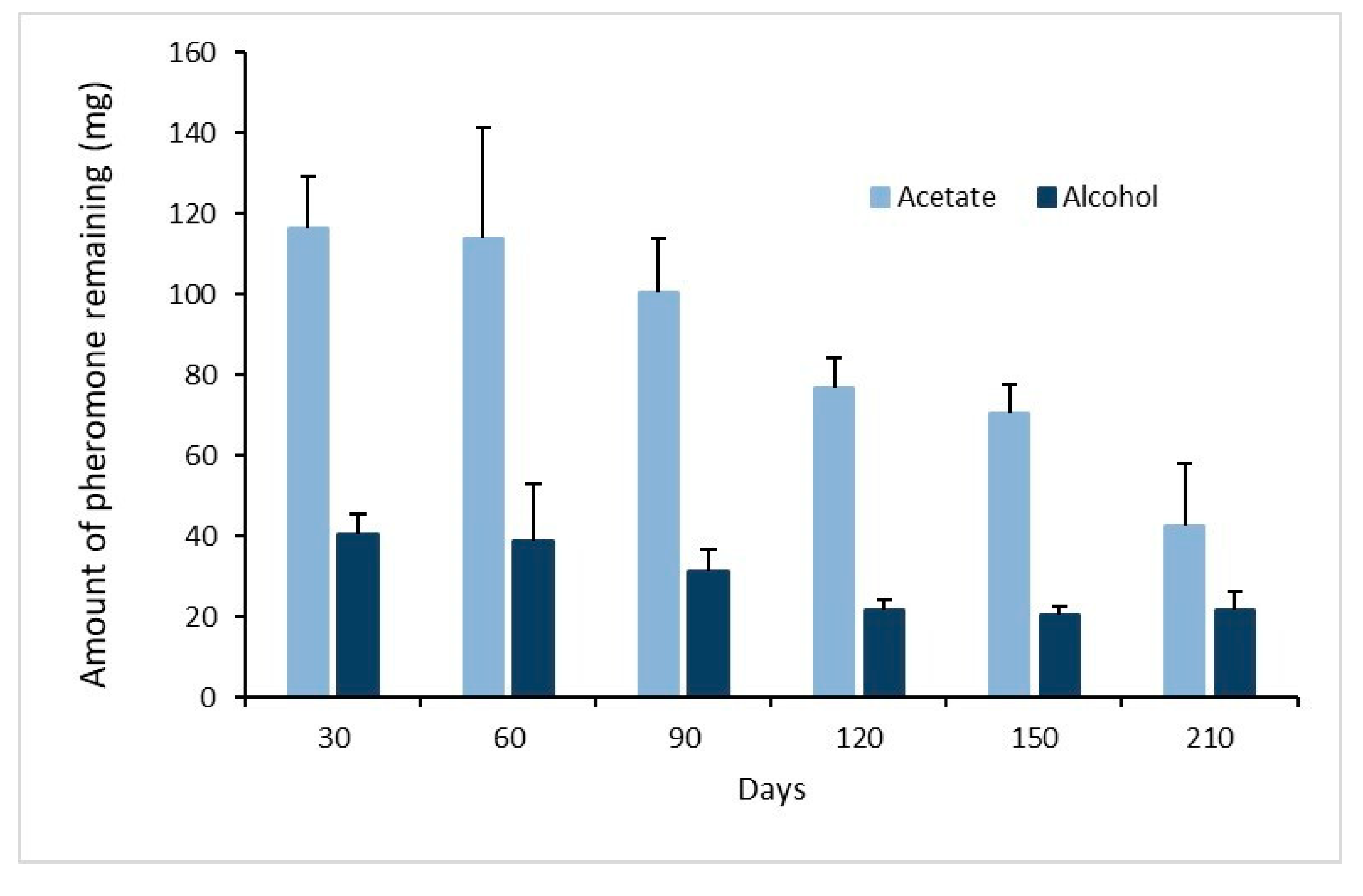
3.2. Remaining Pheromone in Septa after Ageing under Field Conditions
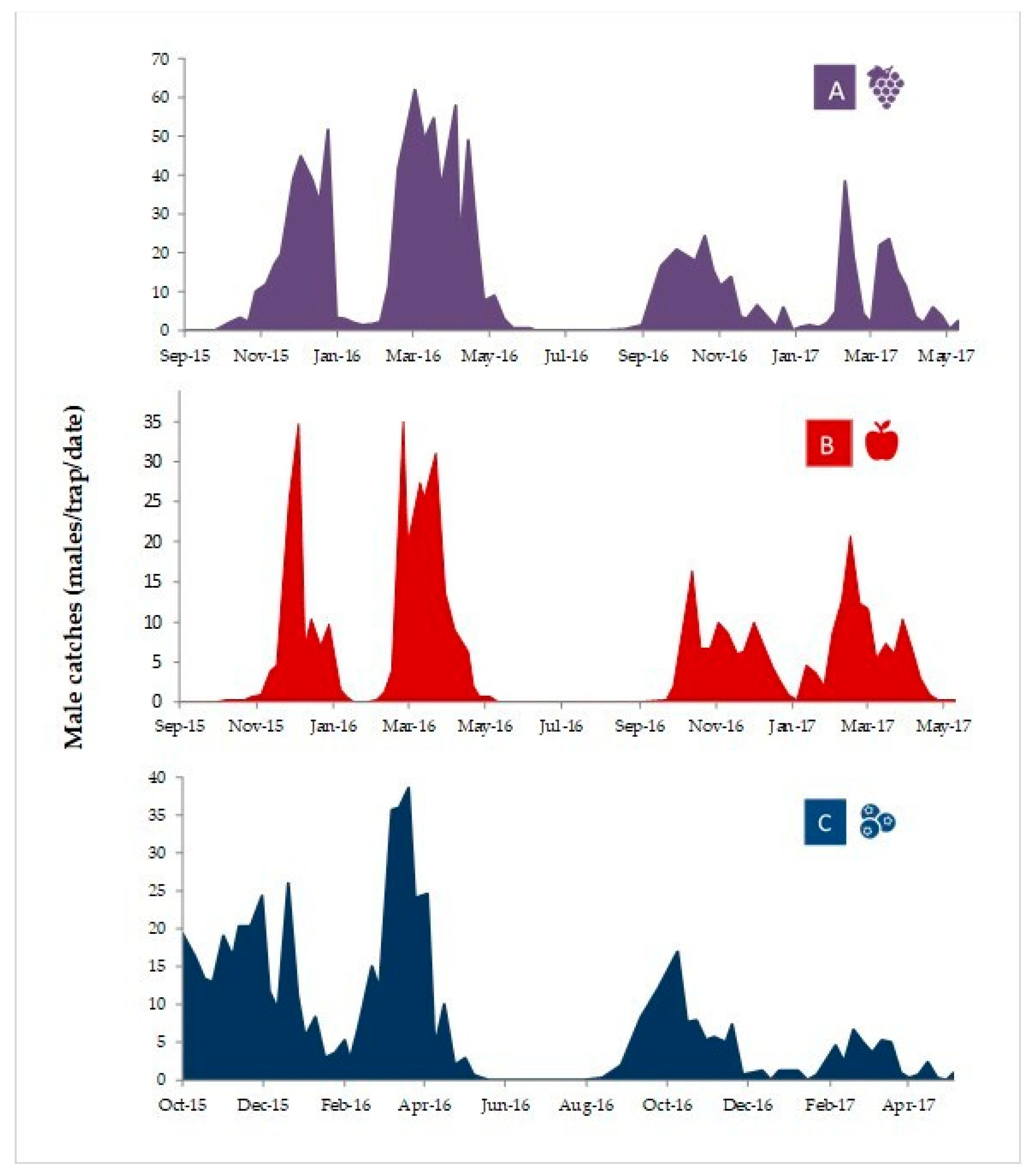
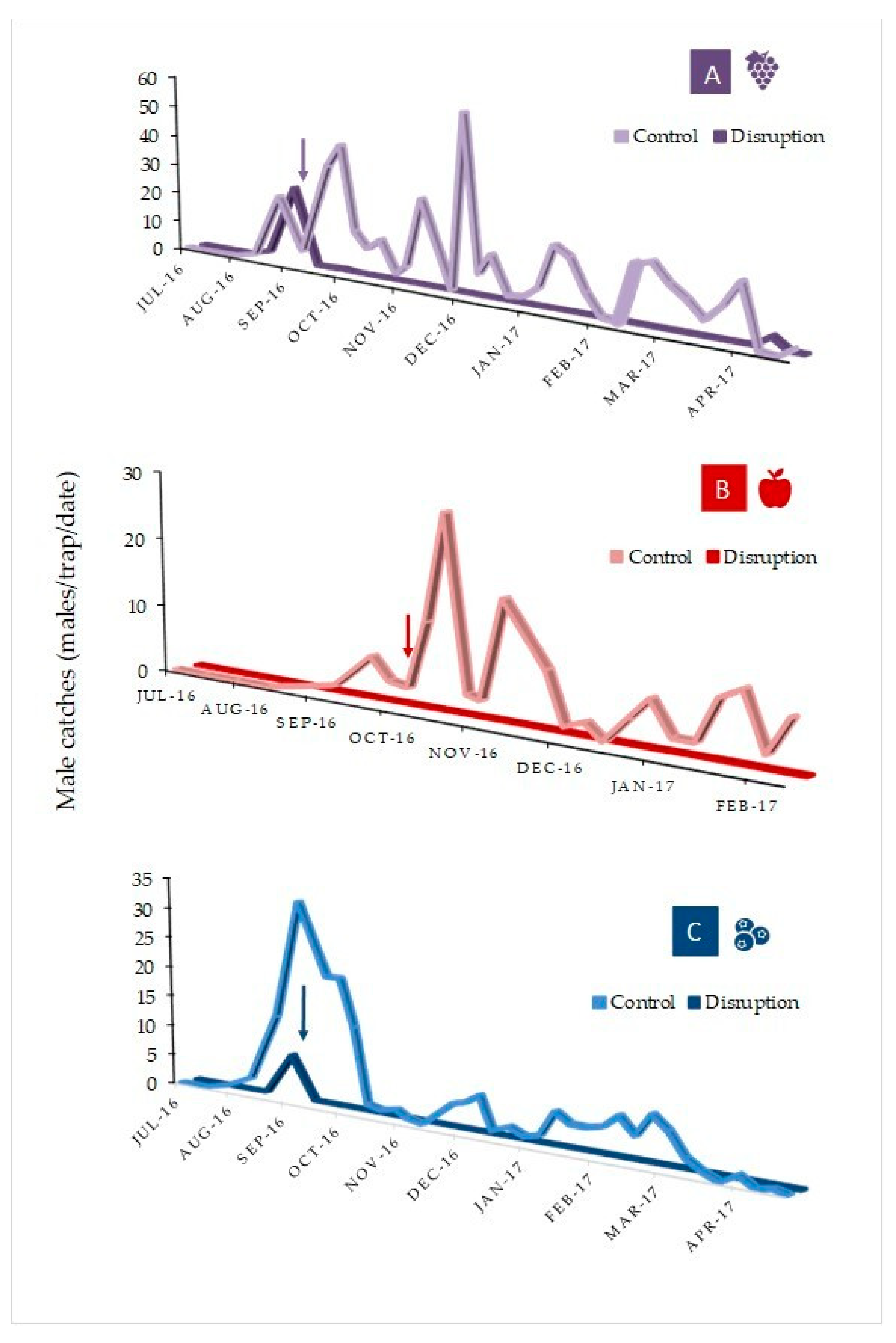
3.3. Proeulia Auraria Adult Male Phenology
3.4. Proeulia auraria Mating Disruption Trials
3.5. Remaining Pheromone in SPLAT Matrix after Aging under Field Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Campos, L.; Faccin, M.; Echeverria, N.; Sazo, L. Distribución y ciclo evolutivo del tortrícido enrollador de la vid Proeulia auraria (Clarke). Agric. Tec. 1981, 41, 249–256. [Google Scholar]
- Álvarez, P.; González, R.H. Biología de la polilla enrolladora del peral Proeulia auraria (Clarke). Rev. Frutic. 1982, 3, 75–80. [Google Scholar]
- Klein Koch, C.; Waterhouse, D.F. The Distribution and Importance of Arthropods Associated with Agriculture and Forestry in Chile; Australian Centre for International Agricultural Research (ACIAR): Canberra, Australia, 2000; p. 231.
- Trematerra, P.; Brown, J.W. Argentine Argyrotaenia (Lepidoptera: Tortricidae): Synopsis and descriptions of two new species. Zootaxa 2004, 574, 1–12. [Google Scholar] [CrossRef]
- CABI (Centre for Agriculture and Bioscience International). Proeulia auraria (Chilean Fruit Tree Leaf Folder). Available online: https://www.cabi.org/isc/datasheet/44569 (accessed on 13 March 2021).
- Razowski, J.; Pelz, V. Tortricidae from Chile (Lepidoptera: Tortricidae). SHILAP Rev. Lepid. 2010, 38, 5–55. [Google Scholar]
- Cepeda, D.; Curkovic, T. A new species of Proeulia Clarke (Lepidoptera: Tortricidae) from Central Chile. Rev. Chil. Entomol. 2020, 46, 493–499. [Google Scholar]
- Cepeda, D.; González, R. Nueva especie del género Proeulia Clarke, con registros adicionales de distribución geográfica para cinco especies (Lepidoptera: Tortricidae). Rev. Chil. Entomol. 2017, 40, 5–12. [Google Scholar]
- Artigas, J.N. Tortricidae. In Entomología Económica. Insectos de Interés Agrícola, Forestal, Médico y Veterinario (Nativos, Introducidos y Susceptibles de ser Introducidos); Universidad de Concepción: Concepción, Chile, 1994; pp. 751–779. ISBN 9562271013. [Google Scholar]
- González, R.H. Las Polillas de la Fruta en Chile (Lepidoptera: Tortricidae; Pyralidae); Serie Ciencias Agronómicas; Universidad de Chile: Santiago, Chile, 2003; pp. 107–138. [Google Scholar]
- Curkovic, T.; Ballesteros, C.; Carpio, C. Manejo integrado de plagas del granado. In Bases Para el Cultivo del Granado en Chile; Henríquez, J.L., Franck, N., Eds.; Serie Ciencias Agronómicas; Universidad de Chile: Santiago, Chile, 2015; pp. 159–231. [Google Scholar]
- Chacón-Fuentes, M.A.; Lizama, M.G.; Parra, L.J.; Seguel, I.E.; Quíroz, A.E. Insect diversity, community composition and damage index on wild and cultivated murtilla. Cienc. Investig. Agrar. 2016, 43, 57–67. [Google Scholar] [CrossRef] [Green Version]
- Biosecurity, N.Z. Biosecurity New Zealand, Standard 155.02.06: Importation of Nursery Stock. Available online: https://www.ippc.int/static/media/files/publications/en/2013/06/05/1125929849079_Biosecurity_New_Zealand.pdf (accessed on 13 March 2021).
- Suffert, M.; Wilstermann, A.; Petter, F.; Schrader, G.; Grousset, F. Identification of new pests likely to be introduced into Europe with the fruit trade. EPPO Bull. 2018, 48, 144–154. [Google Scholar] [CrossRef] [Green Version]
- González, R.; Curkovic, T. Manejo de plagas y degradación de residuos de pesticidas en kiwi. Rev. Frutic. 1994, 15, 5–20. [Google Scholar]
- Ripa, R.; Larral, P. Manejo de plagas en paltos y cítricos. Colecc. Libros Inst. Investig. Agropecu. 2008, 23, 399. [Google Scholar]
- Faccin, M. Feromona Sexual del Enrollador de la vid Proeulia auraria (Clarke) (Lepidoptera: Tortricidae). Bachelor Thesis, Universidad de Chile, Santiago, Chile, 1979. [Google Scholar]
- Roelofs, W.L.; Brown, R.L. Pheromones and evolutionary relationships of Tortricidae. Annu. Rev. Ecol. Syst. 1982, 13, 395–422. [Google Scholar] [CrossRef]
- Reyes-Garcia, L.; Cuevas, Y.; Ballesteros, C.; Curkovic, T.; Löfstedt, C.; Bergmann, J. A 4-component sex pheromone of the Chilean fruit leaf roller Proeulia auraria (Lepidoptera: Tortricidae). Cienc. Investig. Agrar. 2014, 41, 9–10. [Google Scholar] [CrossRef] [Green Version]
- Kahler, E. Evaluación de Cebos Basados en Feromona Sexual para el Monitoreo de Proeulia auraria y Proeulia triquetra en Cultivos de Arándanos. Bachelor Thesis, Universidad Austral de Chile, Valdivia, Chile, 2018. [Google Scholar]
- Valverde-Rodríguez, A. Formulación SPLAT para el control de Proeulia auraria (Lepidoptera: Tortricidae) a través del método de confusión sexual en frutales. Rev. Invest. Agr. 2019, 1, 67–75. [Google Scholar] [CrossRef]
- Stelinski, L.L.; Miller, J.R.; Ledebuhr, R.; Siegert, P.; Gut, L.J. Season-long mating disruption of Grapholita molesta (Lepidoptera: Tortricidae) by one machine application of pheromone in wax drops (SPLAT-OFM). J. Pest Sci. 2007, 80, 109–117. [Google Scholar] [CrossRef]
- Babu, A.; Rodríguez-Saona, C.; Mafra-Neto, A.; Sial, A.A. Efficacy of Attract-and-Kill Formulations Using the Adjuvant Acttra SWD TD for the Management of Spotted-Wing Drosophila in Blueberries, 2020. Arthropod Manag. Tests 2021, 46, 1–2. [Google Scholar] [CrossRef]
- Wallingford, A.K.; Connelly, H.L.; Dore Brind’Amor, G.; Boucher, M.T.; Mafra-Neto, A.; Loeb, G.M. Field evaluation of an oviposition deterrent for management of spotted-wing drosophila, Drosophila suzukii, and potential nontarget effects. J. Econ. Entomol. 2016, 109, 1779–1784. [Google Scholar] [CrossRef] [PubMed]
- Zar, J.H. Biostatistical Analysis, 4th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 1999; p. 929. [Google Scholar]
- Carde, R.T.; Minks, A.K. Control of moth pests by mating disruption: Successes and constraints. Annu. Rev. Entomol. 1995, 40, 559–585. [Google Scholar] [CrossRef]
- Ioriatti, C.; Anfora, G.; Tasin, M.; De Cristofaro, A.; Witzgall, P.; Lucchi, A. Chemical ecology and management of Lobesia botrana (Lepidoptera: Tortricidae). J. Econ. Entomol. 2011, 104, 1125–1137. [Google Scholar] [CrossRef]
- Evenden, M.L.; Judd, G.J.R.; Borden, J.H. Pheromone-mediated mating disruption of Choristoneura rosaceana: Is the most attractive blend really the most effective? Entomol. Exp. Appl. 1999, 90, 37–47. [Google Scholar] [CrossRef]
- Lapointe, S.L.; Stelinski, L.L.; Evens, T.J.; Niedz, R.P.; Hall, D.G.; Mafra-Neto, A. Sensory imbalance as mechanism of orientation disruption in the leafminer Phyllocnistis citrella: Elucidation by multivariate geometric de-signs and response surface models. J. Chem. Ecol. 2009, 35, 896–903. [Google Scholar] [CrossRef] [Green Version]
- Higbee, B.S.; Burks, C.S.; Cardé, R.T. Mating disruption of the navel orangeworm (Lepidoptera: Pyralidae) using widely spaced aerosol dispensers: Is the pheromone blend the most efficacious disruptant? J. Econ. Entomol. 2017, 110, 2056–2061. [Google Scholar] [CrossRef] [PubMed]
- Coracini, M.D.A.; Bengtsson, M.; Reckziegel, A.; Löfqvist, J.; Francke, W.; Vilela, E.F.; Eiras, A.E.; Kovaleski, A.; Witzgall, P. Identification of a Four-Component Sex Pheromone Blend in Bonagota cranaodes (Lepidoptera: Tortricidae). J. Econ. Entomol. 2001, 94, 911–914. [Google Scholar] [CrossRef] [PubMed]
- Knight, A.L. A comparison of gray halo-butyl elastomer and red rubber septa to monitor codling moth (Lepidoptera: Tortricidae) in sex pheromone-treated orchards. J. Entomol. Soc. B. C. 2002, 99, 123–132. [Google Scholar]
- Biever, K.D.; Hostetter, D.L. Phenology and pheromone trap monitoring of the grape berry moth, Endopiza viteana Clemens (Lepidoptera: Tortricidae) in Missouri. J. Entomol. Sci. 1989, 24, 472–481. [Google Scholar] [CrossRef]
- Butler, U.I.; McDonough, L.M. Insect Sex Pheromones: Evaporation Rates of Alcohols and Acetates from Natural Rubber Septa 1. J. Chem. Ecol. 1981, 7, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Fuentes-Contreras, E.; Reyes, M.; Barros, W.; Sauphanor, B. Evaluation of Azinphos-Methyl Resistance and Activity of Detoxifying Enzymes in Codling Moth (Lepidoptera: Tortricidae) from Central Chile. J. Econ. Entomol. 2007, 100, 551–556. [Google Scholar] [CrossRef] [PubMed]
- Lobesia botrana o Polilla del Racimo de la Vid, SAG. Available online: https://www.sag.gob.cl/ambitos-de-accion/lobesia-botrana-o-polilla-del-racimo-de-la-vid (accessed on 19 March 2021).
- Steffan, S.A.; Chasen, E.M.; Deutsch, A.E.; Mafra-Neto, A. Multi-species mating disruption in cranberries (Ericales: Ericaceae): Early evidence using a flowable emulsion. J. Insect Sci. 2017, 17, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Svensson, G.P.; Wang, H.L.; Jirle, E.V.; Rosenberg, O.; Liblikas, I.; Chong, J.M.; Löfstedt, C.; Anderbrant, O. Challenges of pheromone-based mating disruption of Cydia strobilella and Dioryctria abietella in spruce seed orchards. J. Pest Sci. 2018, 91, 639–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovanci, O.B.; Schal, C.; Walgenbach, J.F.; Kennedy, G.G. Comparison of Mating Disruption with Pesticides for Management of Oriental Fruit Moth (Lepidoptera: Tortricidae) in North Carolina Apple Orchards. J. Econ. Entomol. 2005, 98, 1248–1258. [Google Scholar] [CrossRef]
- Light, D.M. Control and Monitoring of Codling Moth (Lepidoptera: Tortricidae) in Walnut Orchards Treated with Novel High-Load, Low-Density “Meso” Dispensers of Sex Pheromone and Pear Ester. Environ. Entomol. 2016, 45, 700–707. [Google Scholar] [CrossRef]
- Vacas, S.; Alfaro, C.; Primo, J.; Navarro-Llopis, V. Studies on the development of a mating disruption system to control the tomato leafminer, Tuta absoluta Povolny (Lepidoptera: Gelechiidae). Pest Manag. Sci. 2011, 67, 1473–1480. [Google Scholar] [CrossRef] [PubMed]
- Arioli, C.J.; Pastori, P.L.; Botton, M.; Garcia, M.S.; Borges, R.; Mafra-Neto, A. Assessment of SPLAT formulations to control Grapholita molesta (Lepidoptera: Tortricidae) in a Brazilian apple orchard. Chil. J. Agric. Res. 2014, 74, 184–190. [Google Scholar] [CrossRef] [Green Version]
- Suckling, D.M.; Baker, G.; Salehi, L.; Woods, B.; Dickens, J.C. Is the Combination of Insecticide and Mating Disruption Synergistic or Additive in Lightbrown Apple Moth, Epiphyas postvittana? PLoS ONE 2016, 11, e0160710. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.R.; Gut, L.J. Mating disruption for the 21st century: Matching technology with mechanism. Environ. Entomol. 2015, 44, 427–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinke, M.D.; Siegert, P.Y.; McGhee, P.S.; Gut, L.J.; Miller, J.R. Pheromone release rate determines whether sexual communication of Oriental fruit moth is disrupted competitively or non-competitively. Entomol. Exp. Appl. 2014, 150, 1–6. [Google Scholar] [CrossRef]
- Curkovic, T.; Sazo, L.; Araya, J.E.; Agurto, L.; Polanco, J. Effect of mating disruption pheromones on Cydia pomonella and other arthropods in pome orchards in Central and Southern Chile. Bol. Sanid. Veg. 2004, 31, 309–318. [Google Scholar]
- Brunner, J.; Welter, S.; Calkins, C.; Hilton, R.; Beers, E.; Dunley, J.; Unruh, T.; Knight, A.; Van Steenwyk, R.; Van Buskirk, P. Mating disruption of codling moth: A perspective from the Western United States. IOBC-WPRS Bull. 2002, 25, 11–20. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| E11-14:OAc + E11-14:OH (μg) | |||
|---|---|---|---|
| Aging Weeks | 50 + 15 | 200 + 60 | 800 + 240 |
| 1 | 13.7 b | 24.7 a | 26.3 a * |
| 2 | 26.3 a | 20.7 a | 20.7 a |
| 3 | 21.7 a | 24.0 a | 11.0 b |
| 4 | 22.7 a | 27.7 a | 24.0 a |
| 5 | 8.0 b | 27.7 a | 23.0 a |
| 6 | 17.7 b | 19.7 b | 30.0 a |
| 7 | 16.0 b | 28.3 a | 23.7 a |
| 8 | 13.3 b | 21.0 a | 22.3 a |
| 9 | 16.7 b | 27.7 a | 17.0 b |
| 10 | 14.7 b | 26.0 a | 21.0 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flores, M.F.; Bergmann, J.; Ballesteros, C.; Arraztio, D.; Curkovic, T. Development of Monitoring and Mating Disruption against the Chilean Leafroller Proeulia auraria (Lepidoptera: Tortricidae) in Orchards. Insects 2021, 12, 625. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12070625
Flores MF, Bergmann J, Ballesteros C, Arraztio D, Curkovic T. Development of Monitoring and Mating Disruption against the Chilean Leafroller Proeulia auraria (Lepidoptera: Tortricidae) in Orchards. Insects. 2021; 12(7):625. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12070625
Chicago/Turabian StyleFlores, M. Fernanda, Jan Bergmann, Carolina Ballesteros, Diego Arraztio, and Tomislav Curkovic. 2021. "Development of Monitoring and Mating Disruption against the Chilean Leafroller Proeulia auraria (Lepidoptera: Tortricidae) in Orchards" Insects 12, no. 7: 625. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12070625