
Drosophila suzukii Susceptibility to the Oral Administration of Bacillus thuringiensis, Xenorhabdus nematophila and Its Secondary Metabolites




Abstract
:Simple Summary
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Chemicals and Instruments
2.2. Target Insect
2.3. Bacterial Strains and Cultures
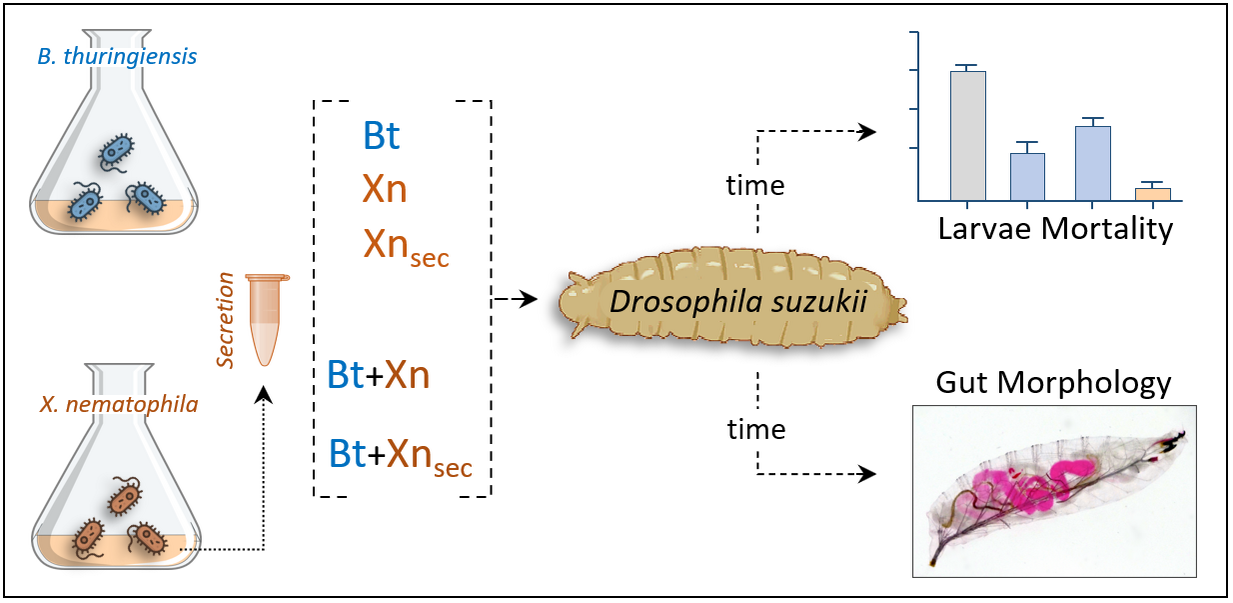
2.4. Oral Administrations: Single and Combined Treatments
2.5. SDS-PAGE Analysis of X. nematophila Secretions
2.6. Hemolymph Collection and Total Hemocytes Count (THC)
2.7. Interference of Bioinsecticide Secretions on Bacterial Growth
2.8. Analysis of the SWD Larvae Gut Morphology
2.9. Data Analysis
3. Results
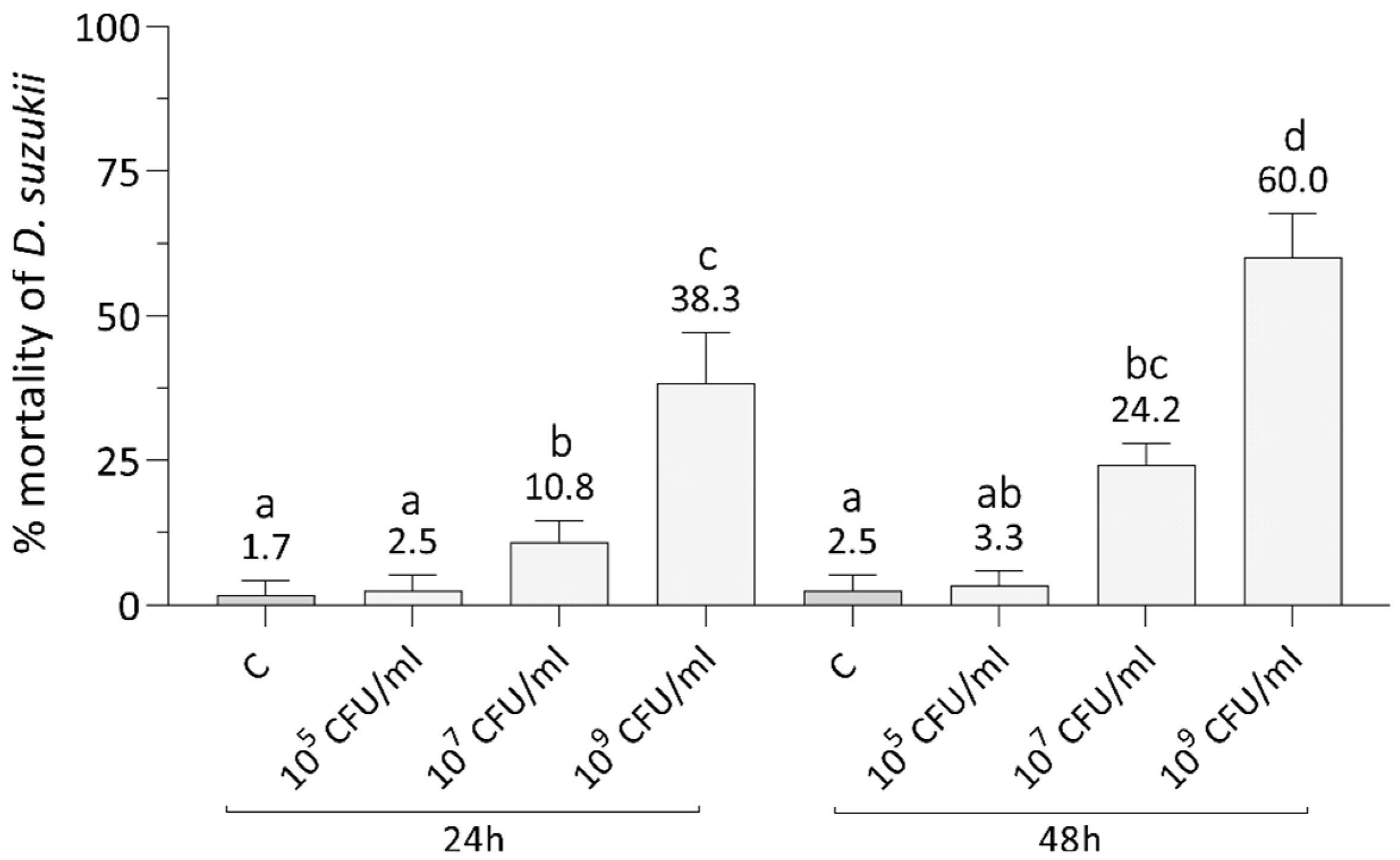
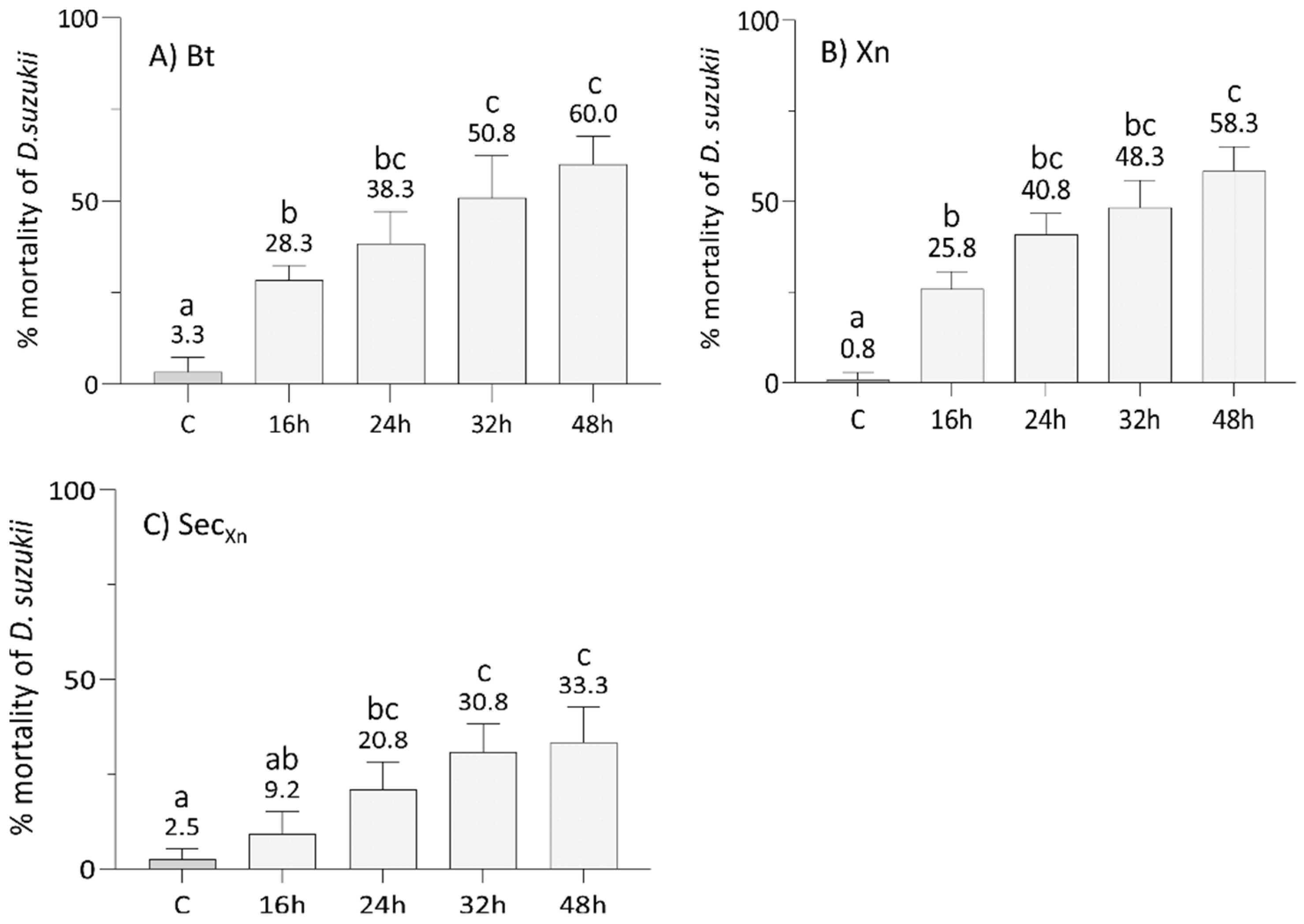
3.1. Bioinsecticides Administration
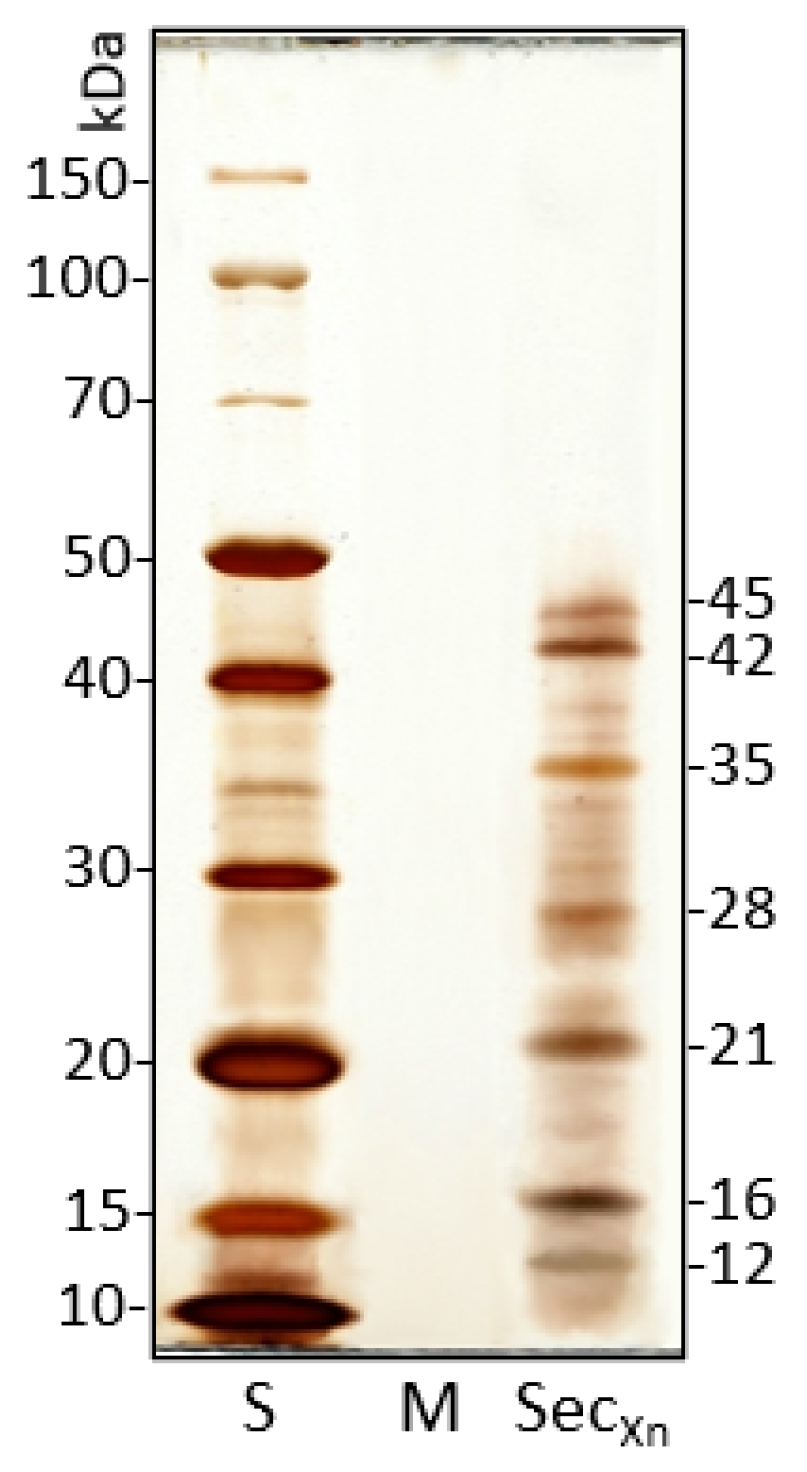
3.2. Secretion Protein Pool of X. nematophila
3.3. Mutual Interference of Secretions on Bacterial Growth
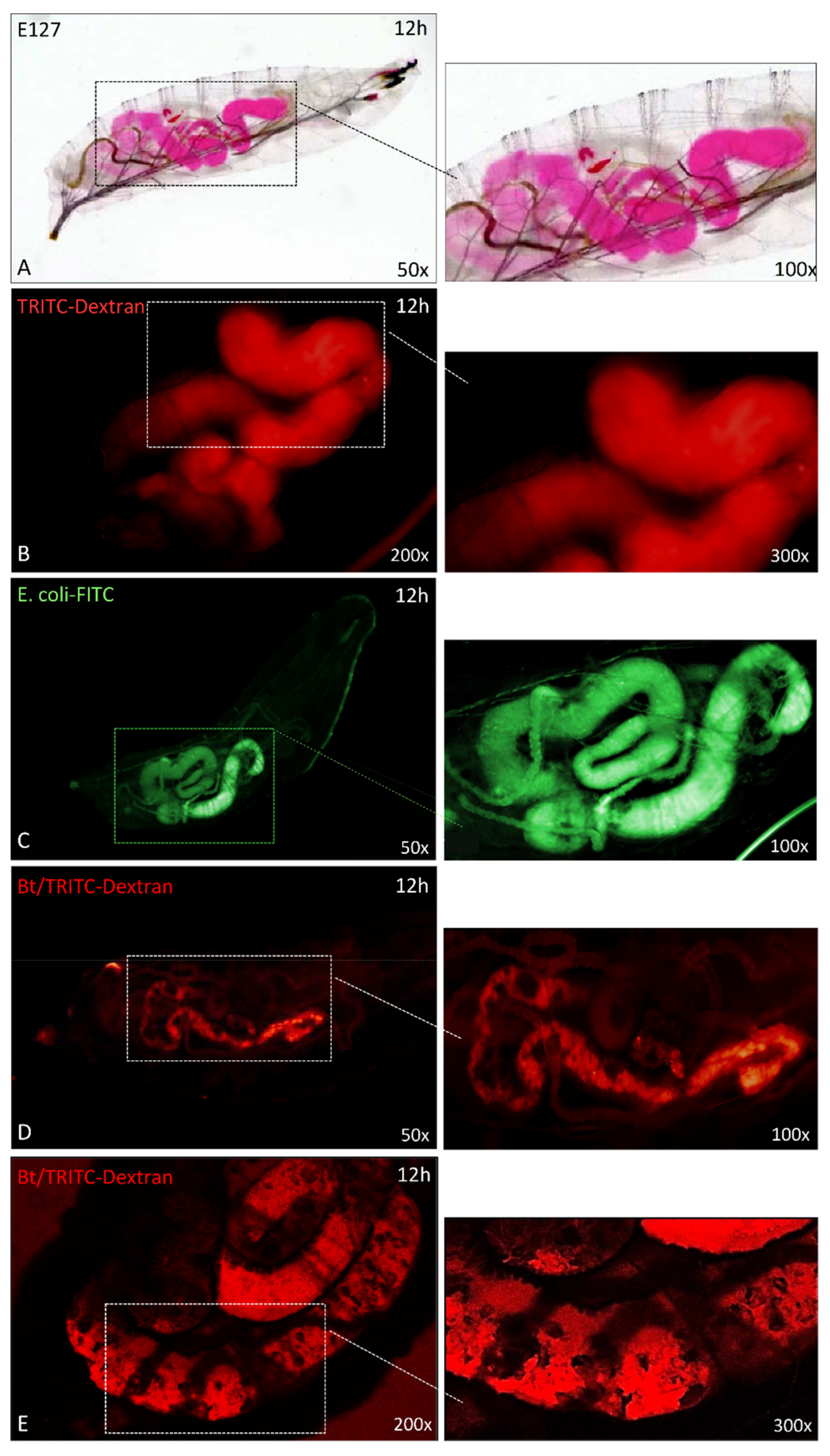
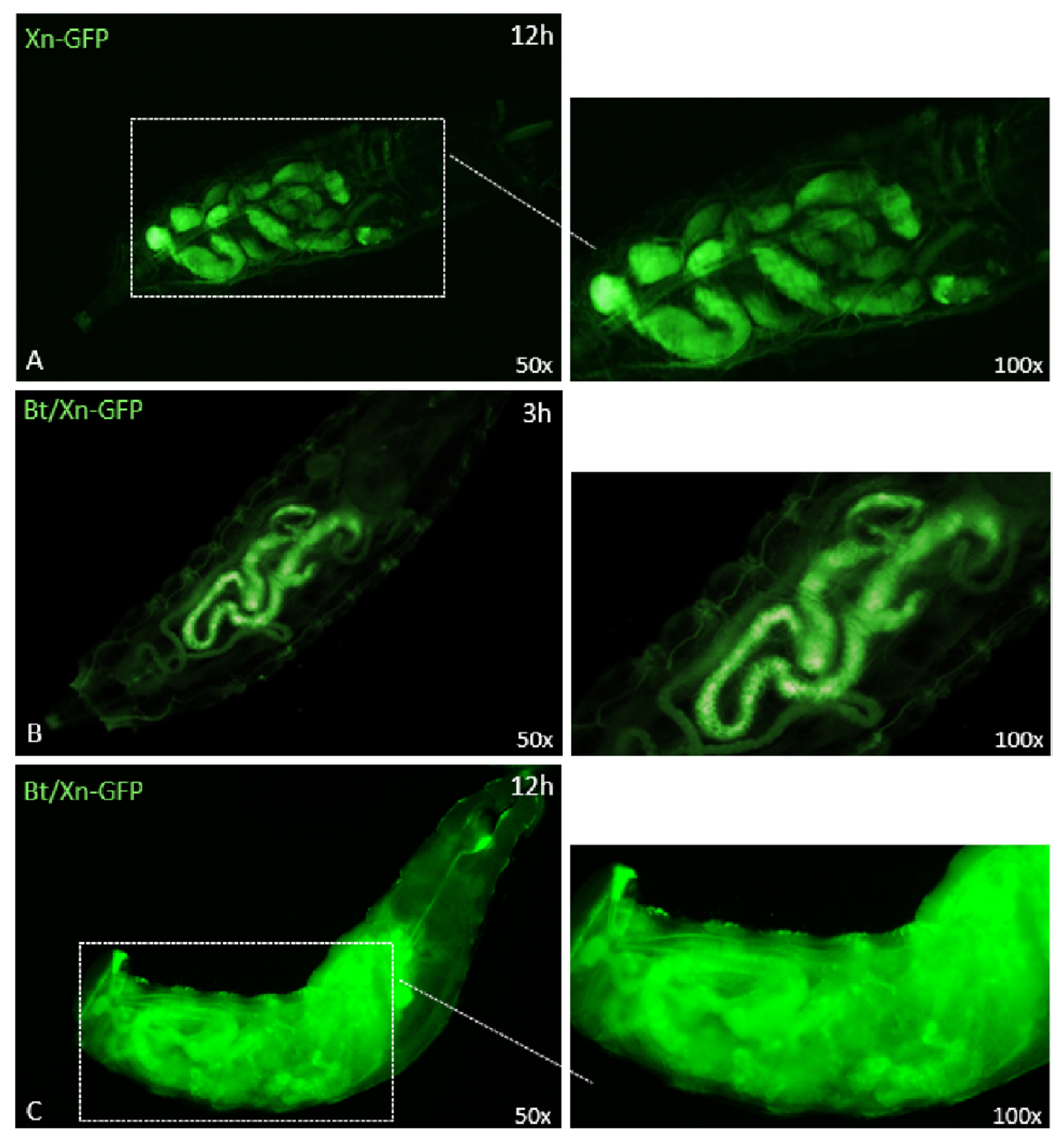
3.4. Effects of Entomopathogenic Bacteria on SWD Gut
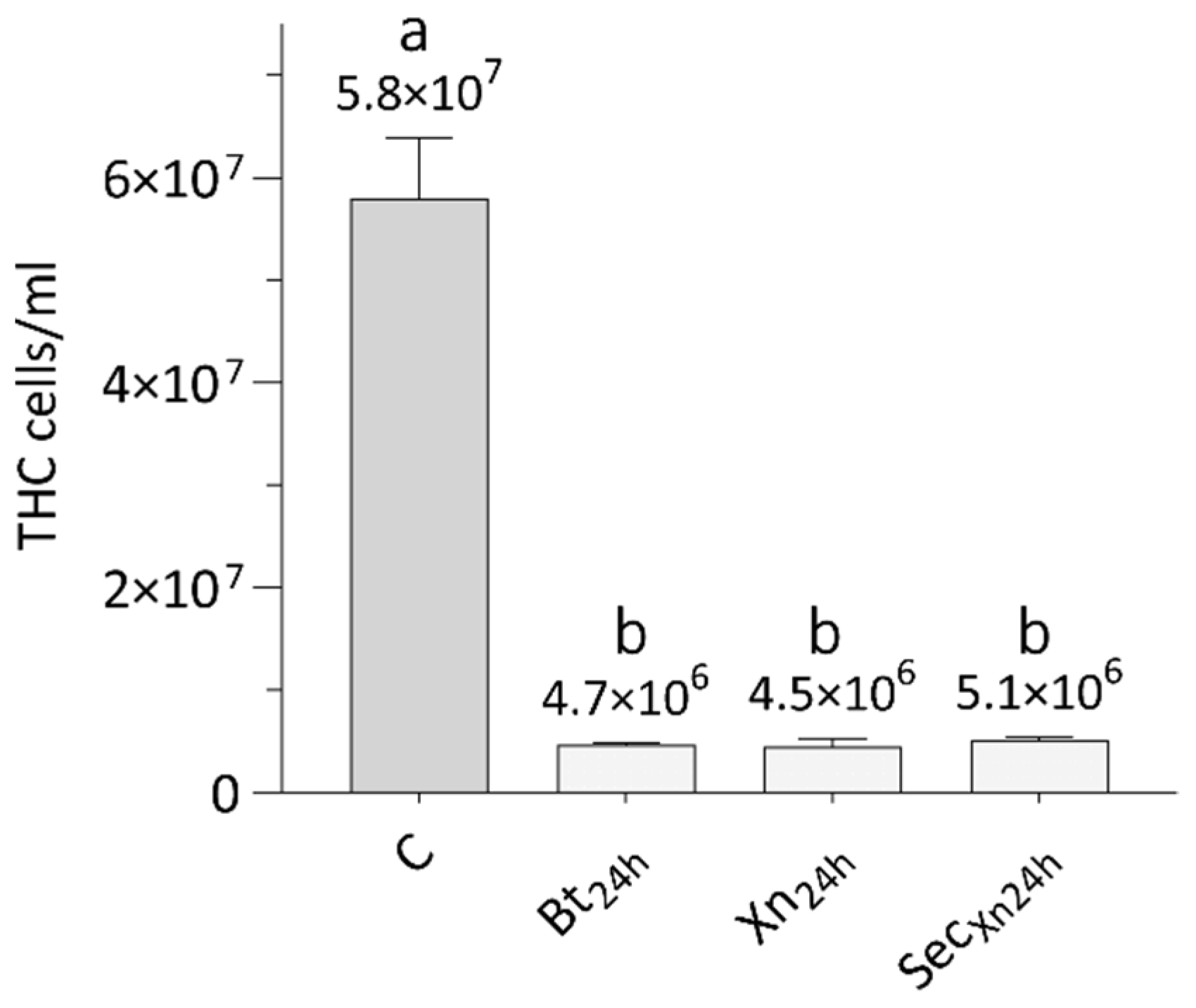
3.5. Effects of Bioinsecticides on Total Hemocytes Count (THC)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Asplen, M.K.; Anfora, G.; Biondi, A.; Choi, D.S.; Chu, D.; Daane, K.M.; Patricia Gibert, P.; Gutierrez, A.P.; Hoelmer, K.A.; Hutchison, W.D.; et al. Invasion biology of spotted wing Drosophila (Drosophila suzukii): A global perspective and future priorities. J. Pest. Sci. 2015, 88, 469–494. [Google Scholar] [CrossRef]
- Shawer, R. Chemical Control of Drosophila suzukii. In Drosophila suzukii Management; Garcia, F.R.M., Ed.; Springer: Cham, Switzerland, 2020; pp. 133–142. [Google Scholar]
- Dam, D.; Molitor, D.; Beyer, M. Natural compounds for controlling Drosophila suzukii: A review. Agron. Sustain. Dev. 2019, 39, 53. [Google Scholar] [CrossRef]
- Wang, X.; Daane, K.M.; Hoelmer, K.A.; Lee, J.C. Biological Control of Spotted-Wing Drosophila: An Update on Promising Agents. In Drosophila suzukii Management; Garcia, F.R.M., Ed.; Springer: Cham, Switzerland, 2020; pp. 143–167. [Google Scholar]
- Lee, J.; Wang, X.G.; Daane, K.M.; Hoelmer, K.A.; Isaacs, R.; Sial, A.; Walton, V.M. Biological control of spotted-wing drosophila (Diptera: Drosophilidae)—Current and pending tactics. J. Integr. Pest Manag. 2019, 10, 1–9. [Google Scholar] [CrossRef]
- Garriga, A.; Morton, A.; Garcia-del-Pino, F. Is Drosophila suzukii as susceptible to entomopathogenic nematodes as Drosophila melanogaster? J. Pest Sci. 2017, 91, 789–798. [Google Scholar] [CrossRef]
- Hübner, A.; Englert, C.; Herz, A. Effect of entomopathogenic nematodes on different developmental stages of Drosophila suzukii in and outside fruits. BioControl 2017, 62, 669–680. [Google Scholar] [CrossRef]
- Garriga, A.; Mastore, M.; Morton, A.; Garcia-del-Pino, F.; Brivio, M.F. Immune Response of Drosophila suzukii Larvae to Infection with the Nematobacterial Complex Steinernema carpocapsae-Xenorhabdus nematophila. Insects 2020, 11, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastore, M.; Quadroni, S.; Brivio, M.F. Susceptibility of Drosophila suzukii larvae to the combined administration of the entomopathogens Bacillus thuringiensis and Steinernema carpocapsae. Sci. Rep. 2021, 11, 8149. [Google Scholar] [CrossRef]
- Labaude, S.; Griffin, C.T. Transmission success of entomopathogenic nematodes used in pest control. Insects 2018, 9, 72. [Google Scholar] [CrossRef] [Green Version]
- Herbert, E.E.; Goodrich-Blair, H. Friend and foe: The two faces of Xenorhabdus nematophila. Nat. Rev. Microbiol. 2007, 5, 634–646. [Google Scholar] [CrossRef]
- Tobias, N.J.; Wolff, H.; Djahanschiri, B.; Grundmann, F.; Kronenwerth, M.; Shi, Y.M.; Simonyi, S.; Grün, P.; David Shapiro-Ilan, D.; Pidot, S.J.; et al. Natural product diversity associated with the nematode symbionts Photorhabdus and Xenorhabdus. Nat. Microbiol. 2017, 2, 1676–1685. [Google Scholar] [CrossRef]
- Mollah, M.M.I.; Kim, Y. Virulent secondary metabolites of entomopathogenic bacteria genera, Xenorhabdus and Photorhabdus, inhibit phospholipase A2 to suppress host insect immunity. BMC Microbiol. 2020, 20, 359. [Google Scholar] [CrossRef]
- da Silva, O.S.; Prado, G.R.; da Silva, J.L.; Silva, C.E.; da Costa, M.; Heermann, R. Oral toxicity of Photorhabdus luminescens and Xenorhabdus nematophila (Enterobacteriaceae) against Aedes aegypti (Diptera: Culicidae). Parasitol. Res. 2013, 112, 2891–2896. [Google Scholar] [CrossRef]
- Hwang, J.; Park, Y.; Kim, Y.; Hwang, J.; Lee, D. An entomopathogenic bacterium, Xenorhabdus nematophila, suppresses expression of antimicrobial peptides controlled by Toll and Imd pathways by blocking eicosanoid biosynthesis. Arch. Insect Biochem. Physiol. 2013, 83, 151–169. [Google Scholar] [CrossRef] [PubMed]
- Brivio, M.F.; Toscano, A.; De Pasquale, S.M.; De Lerma Barbaro, A.; Giovannardi, S.; Finzi, G.; Mastore, M. Surface protein components from entomopathogenic nematodes and their symbiotic bacteria: Effects on immune responses of the greater wax moth, Galleria mellonella (Lepidoptera, Pyralidae). Pest. Manag. Sci. 2018, 74, 2089–2099. [Google Scholar] [CrossRef] [PubMed]
- De Lerma Barbaro, A.; Gariboldi, M.B.; Mastore, M.; Brivio, M.F.; Giovannardi, S. In Vivo Effects of a Pro-PO System Inhibitor on the Phagocytosis of Xenorhabdus Nematophila in Galleria Mellonella Larvae. Insects 2019, 10, 263. [Google Scholar] [CrossRef] [Green Version]
- Cossentine, J.; Robertson, M.; Xu, D. Biological Activity of Bacillus thuringiensis in Drosophila suzukii (Diptera: Drosophilidae). J. Econ. Entomol. 2016, 109, 1071–1078. [Google Scholar] [CrossRef] [PubMed]
- Biganski, S.; Jehle, J.A.; Kleespies, R.G. Bacillus thuringiensis serovar. israelensis has no effect on Drosophila suzukii Matsumura. J. Appl. Entomol. 2018, 142, 33–36. [Google Scholar]
- Mastore, M.; Quadroni, S.; Toscano, A.; Mottadelli, N.; Brivio, M.F. Susceptibility to entomopathogens and modulation of basal immunity in two insect models at different temperatures. J. Thermal Biol. 2019, 79, 15–23. [Google Scholar] [CrossRef]
- Park, Y.; Kyo Jung, J.; Kim, Y. A mixture of Bacillus thuringiensis subsp. israelensis with Xenorhabdus nematophila-cultured broth enhances toxicity against mosquitoes Aedes albopictus and Culex pipiens pallens (Diptera: Culicidae). J. Econ. Entomol. 2016, 109, 1086–1093. [Google Scholar] [CrossRef]
- Yaroslavtseva, O.N.; Dubovskiy, I.M.; Khodyrev, V.P.; Duisembekov, B.A.; Kryukov, V.Y.; Glupov, V.V. Immunological mechanisms of synergy between fungus Metarhizium robertsii and bacteria Bacillus thuringiensis ssp. morrisoni on Colorado potato beetle larvae. J. Insect Physiol. 2017, 96, 14–20. [Google Scholar] [CrossRef]
- Park, Y.; Kim, Y. Eicosanoids rescue Spodoptera exigua infected with Xenorhabdus nematophilus, the symbiotic bacteria to the entomopathogenic nematode Steinernema carpocapsae. J. Insect Physiol. 2000, 46, 1469–1476. [Google Scholar] [CrossRef]
- Hazenbos, W.L.; van den Berg, B.M.; van’t Wout, J.W.; Mooi, F.R.; van Furth, R. Virulence factors determine attachment and ingestion of nonopsonized and opsonized Bordetella pertussis by human monocytes. Infect Immun. 1994, 62, 4818–4824. [Google Scholar] [CrossRef] [Green Version]
- Laemmli UK, Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [CrossRef]
- Palma, L.; Muñoz, D.; Berry, C.; Murillo, J.; Caballero, P. Bacillus thuringiensis toxins: An overview of their biocidal activity. Toxins 2014, 6, 3296–3325. [Google Scholar] [CrossRef] [Green Version]
- Nawrot-Esposito, M.P.; Babin, A.; Pasco, M.; Poirié, M.; Gatti, J.L.; Gallet, A. Bacillus thuringiensis Bioinsecticides Induce Developmental Defects in Non-Target Drosophila melanogaster Larvae. Insects 2020, 11, 697. [Google Scholar] [CrossRef]
- Ben-Dov, E. Bacillus thuringiensis subsp. israelensis and its dipteran-specific toxins. Toxins 2014, 6, 1222–1243. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, P.; Banerjee, G. Recent advancement on chemical arsenal of Bt toxin and its application in pest management system in agricultural field. 3 Biotech 2018, 8, 201. [Google Scholar] [CrossRef] [PubMed]
- Castillo, J.C.; Reynolds, S.E.; Eleftherianos, I. Insect immune responses to nematode parasites. Trends Parasitol. 2011, 27, 537–547. [Google Scholar] [CrossRef]
- Cooper, D.; Eleftherianos, I. Parasitic Nematode Immunomodulatory Strategies: Recent Advances and Perspectives. Pathogens 2016, 5, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brivio, M.F.; Mastore, M. Nematobacterial Complexes and Insect Hosts: Different Weapons for the Same War. Insects 2018, 9, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brivio, M.F.; Mastore, M. When Appearance Misleads: The Role of the Entomopathogen Surface in the Relationship with Its Host. Insects 2020, 11, 387. [Google Scholar] [CrossRef]
- Hinchliffe, S.J.; Hares, M.C.; Dowling, A.J.; ffrench-Constant, R.H. Insecticidal Toxins from the Photorhabdus and Xenorhabdus Bacteria. The Open Toxicol. J. 2010, 3, 83–100. [Google Scholar] [CrossRef]
- Shi, H.; Zeng, H.; Yang, X.; Zhao, J.; Chen, M.; Qiu, D. An insecticidal protein from Xenorhabdus ehlersii triggers prophenoloxidase activation and hemocyte decrease in Galleria mellonella. Curr. Microbiol. 2012, 64, 604–610. [Google Scholar] [CrossRef]
- Ozakman, Y.; Eleftherianos, I. Immune interactions between Drosophila and the pathogen Xenorhabdus. Microbiol. Res. 2020, 240, 126568. [Google Scholar] [CrossRef]
- Park, Y.; Kim, Y.; Stanley, D. The bacterium Xenorhabdus nematophila inhibits phospholipases A2 from insect, prokaryote, and vertebrate sources. Naturwissenschaften 2004, 91, 371–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheets, J.J.; Hey, T.D.; Fencil, K.J.; Burton, S.L.; Ni, W.; Lang, A.E.; Benz, R.; Aktories, K. Insecticidal toxin complex proteins from Xenorhabdus nematophilus: Structure and pore formation. J. Biol. Chem. 2011, 286, 22742–22749. [Google Scholar] [CrossRef] [Green Version]
- Kim, I.H.; Aryal, S.K.; Aghai, D.T.; Casanova-Torres, Á.M.; Hillman, K.; Kozuch, M.P.; Mans, E.J.; Mauer, T.J.; Ogier, J.-C.; Ensign, J.C.; et al. The insect pathogenic bacterium Xenorhabdus innexi has attenuated virulence in multiple insect model hosts yet encodes a potent mosquitocidal toxin. BMC Genom. 2017, 18, 927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva, W.J.; Pilz-Júnior, H.L.; Heermann, R.; da Silva, O.S. The great potential of entomopathogenic bacteria Xenorhabdus and Photorhabdus for mosquito control: A review. Parasit. Vectors 2020, 13, 376. [Google Scholar] [CrossRef]
- ffrench-Constant, R.H.; Bowen, D.J. Novel insecticidal toxins from nematode-symbiotic bacteria. Cell Mol. Life Sci. 2000, 57, 828–833. [Google Scholar] [CrossRef] [PubMed]
- Forst, S.; Dowds, B.; Boemare, N.; Stackebrandt, E. Xenorhabdus and Photorhabdus spp.: Bugs that kill bugs. Annu. Rev. Microbiol. 1997, 51, 47–72. [Google Scholar] [CrossRef]
- Dreyer, J.; Malan, A.P.; Dicks Leon, M.T. Bacteria of the Genus Xenorhabdus, a Novel Source of Bioactive Compounds. Front. Microbiol. 2018, 9, 3177. [Google Scholar] [CrossRef] [Green Version]
- Ruiu, L. Insect Pathogenic Bacteria in Integrated Pest Management. Insects 2015, 6, 352–367. [Google Scholar] [CrossRef] [Green Version]
- Kajla, M.K. Symbiotic Bacteria as Potential Agents for Mosquito Control. Trends Parasitol. 2020, 36, 4–7. [Google Scholar] [CrossRef]
- Valtierra-de-Luis, D.; Villanueva, M.; Berry, C.; Caballero, P. Potential for Bacillus thuringiensis and Other Bacterial Toxins as Biological Control Agents to Combat Dipteran Pests of Medical and Agronomic Importance. Toxins 2020, 12, 773. [Google Scholar] [CrossRef]
- Nielsen-LeRoux, C.; Gaudriault, S.; Ramarao, N.; Lereclus, D.; Givaudan, A. How the insect pathogen bacteria Bacillus thuringiensis and Xenorhabdus/Photorhabdus occupy their hosts. Curr. Opin. Microbiol. 2012, 15, 220–231. [Google Scholar] [CrossRef]
- Belousova, M.E.; Malovichko, Y.V.; Shikov, A.E.; Nizhnikov, A.A.; Antonets, K.S. Dissecting the Environmental Consequences of Bacillus thuringiensis Application for Natural Ecosystems. Toxins 2021, 13, 355. [Google Scholar] [CrossRef] [PubMed]
- Steinigeweg, C.; Alkassab, A.T.; Beims, H.; Eckert, J.H.; Richter, D.; Pistorius, J. Assessment of the impacts of microbial plant protection products containing Bacillus thuringiensis on the survival of adults and larvae of the honeybee (Apis mellifera). Environ. Sci. Pollut. Res. 2021, 28, 29773–29780. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.C.; Kim, Y.G. Potentiating effect of Bacillus thuringiensis subsp. kurstaki on pathogenicity of entomopathogenic bacterium Xenorhabdus nematophila K1 against diamondback moth (Lepidoptera: Plutellidae). J. Econ. Entomol. 2007, 100, 246–250. [Google Scholar] [CrossRef]
- Opisa, S.; du Plessis, H.; Akutse, K.S.; Fiaboe, K.K.M.; Ekesi, S. Effects of Entomopathogenic fungi and Bacillus thuringiensis-based biopesticides on Spoladea recurvalis (Lepidoptera: Crambidae). J. Appl. Entomol. 2018, 142, 617–626. [Google Scholar] [CrossRef]
- Li, E.T.; Zhang, S.; Li, K.B.; Nyamwasaa, I.; Li, J.Q.; Li, X.F.; Qin, J.H.; Yin, J. Efficacy of entomopathogenic nematode and Bacillus thuringiensis combinations against Holotrichia parallela (Coleoptera: Scarabaeidae) larvae. Biol. Control 2021, 152, 104469. [Google Scholar] [CrossRef]
- Fukruksa, C.; Yimthin, T.; Suwannaroj, M.; Muangpat, P.; Tandhavanant, S.; Thanwisai, A.; Vitta, A. Isolation and identification of Xenorhabdus and Photorhabdus bacteria associated with entomopathogenic nematodes and their larvicidal activity against Aedes aegypti. Parasites Vectors 2017, 10, 440. [Google Scholar] [CrossRef] [Green Version]
- Bode, H.B. Entomopathogenic bacteria as a source of secondary metabolites. Curr. Opin. Chem. Biol. 2009, 13, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Koppenhofer, A.M.; Kaya, H.K. Additive and synergistic interaction between entomopathogenic nematodes and Bacillus thuringiensis for scarab grub control. Biol. Control. 1997, 8, 131–137. [Google Scholar] [CrossRef]
- Gassmann, A.J.; Stock, S.P.; Sisterson, M.S.; Carrière, Y.; Tabashnik, B.E. Synergism between entomopathogenic nematodes and Bacillus thuringiensis crops: Integrating biological control and resistance management. J. Appl. Ecol. 2008, 45, 957–966. [Google Scholar] [CrossRef]
- NanGong, Z.; Wang, Q.; Song, P.; Hao, J.; Yang, Q.; Wang, L. Synergism between Bacillus thuringiensis and Xenorhabdus nematophila against resistant and susceptible Plutella xylostella (Lepidoptera: Plutellidae). Biocontrol Sci. Technol. 2016, 26, 1411–1419. [Google Scholar] [CrossRef]
- Park, Y.; Kim, Y.; Tunaz, H.; Stanley, D.W. An entomopathogenic bacterium, Xenorhabdus nematophila, inhibits hemocytic phospholipase A2 (PLA2) in tobacco hornworms Manduca sexta. J. Invertebr. Pathol. 2004, 86, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Ahmed, S.; Stanley, D.; An, C. Eicosanoid-mediated immunity in insects. Dev. Comp. Immunol. 2018, 83, 130–143. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mastore, M.; Caramella, S.; Quadroni, S.; Brivio, M.F. Drosophila suzukii Susceptibility to the Oral Administration of Bacillus thuringiensis, Xenorhabdus nematophila and Its Secondary Metabolites. Insects 2021, 12, 635. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12070635
Mastore M, Caramella S, Quadroni S, Brivio MF. Drosophila suzukii Susceptibility to the Oral Administration of Bacillus thuringiensis, Xenorhabdus nematophila and Its Secondary Metabolites. Insects. 2021; 12(7):635. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12070635
Chicago/Turabian StyleMastore, Maristella, Sara Caramella, Silvia Quadroni, and Maurizio Francesco Brivio. 2021. "Drosophila suzukii Susceptibility to the Oral Administration of Bacillus thuringiensis, Xenorhabdus nematophila and Its Secondary Metabolites" Insects 12, no. 7: 635. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12070635