
Morphological and Ultrastructural Characterization of Hemocytes in an Insect Model, the Hematophagous Dipetalogaster maxima (Hemiptera: Reduviidae)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Hemolymph Collection
2.3. Phase Contrast Microscopy
2.4. Fluorescence Microscopy
2.5. Transmission Electron Microscopy (TEM)
2.6. Flow Cytometry
3. Results
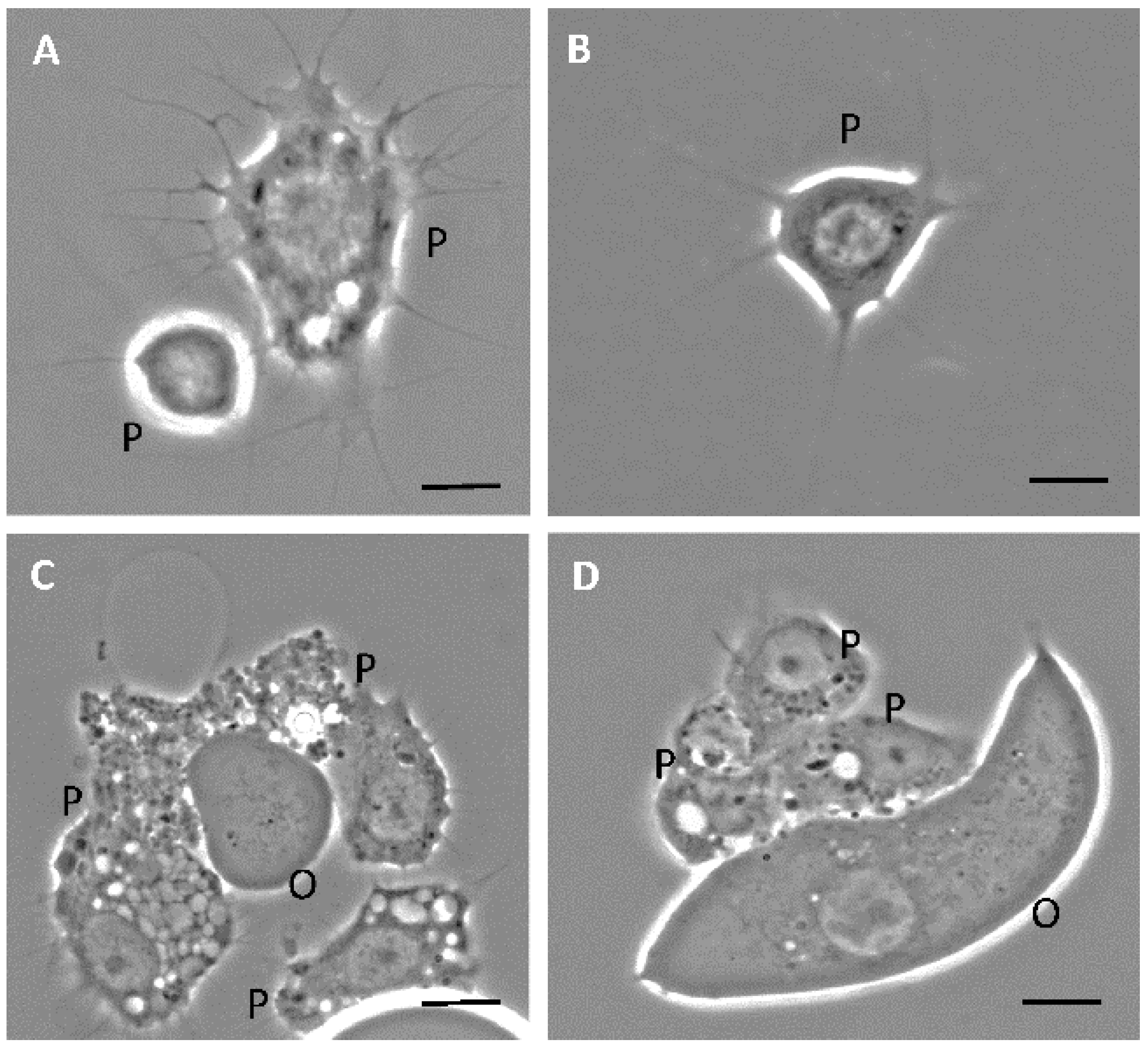
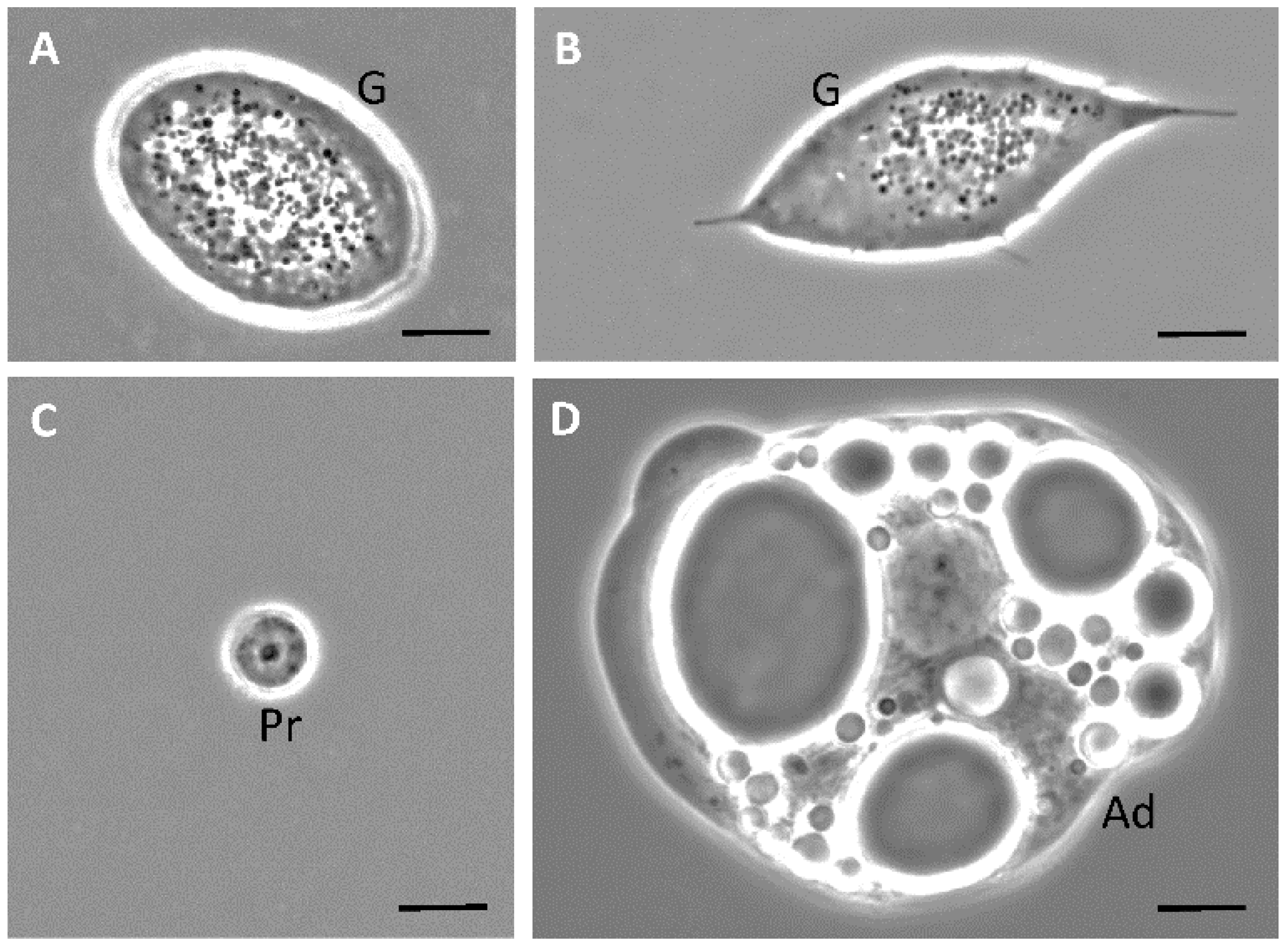
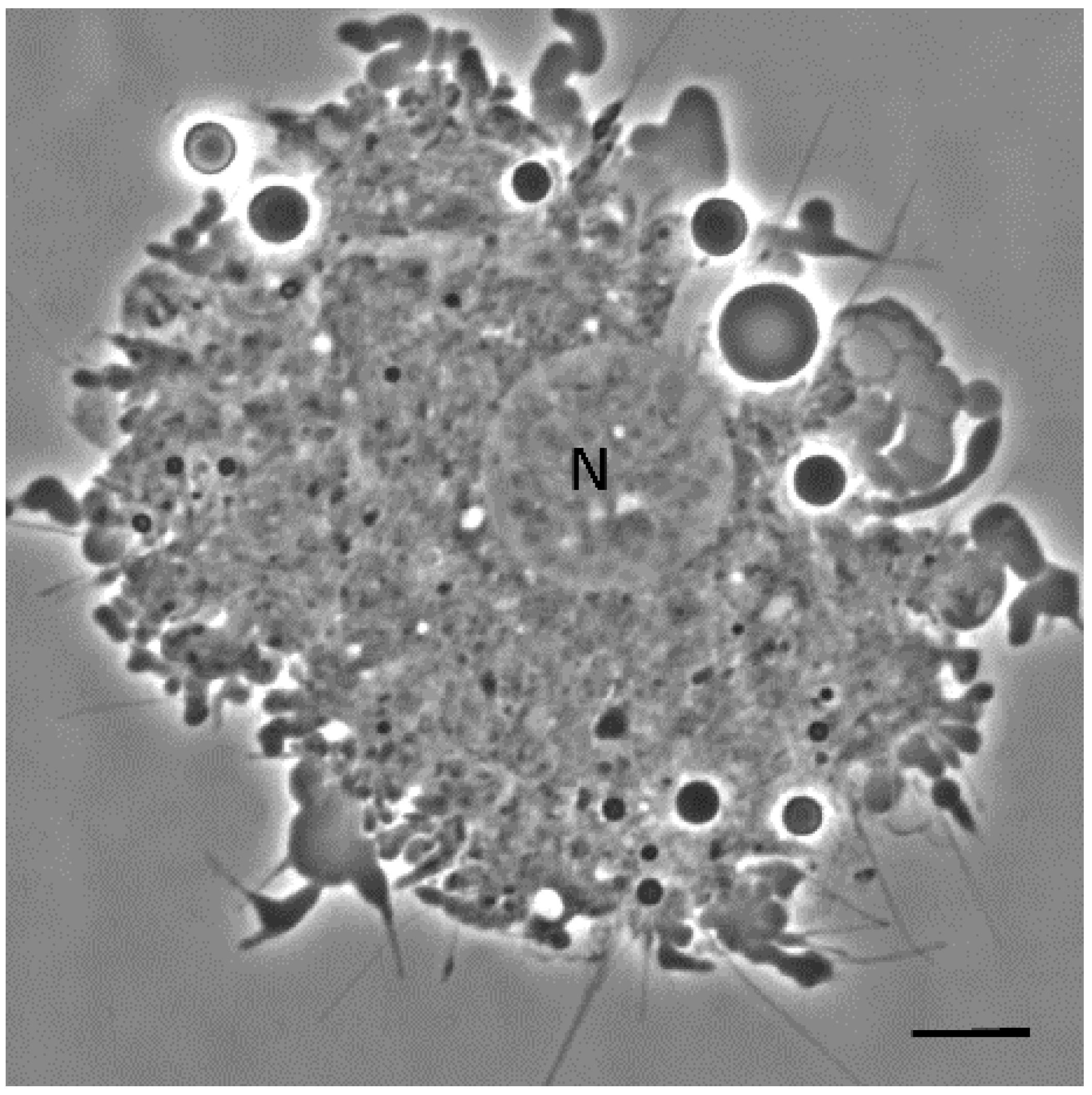
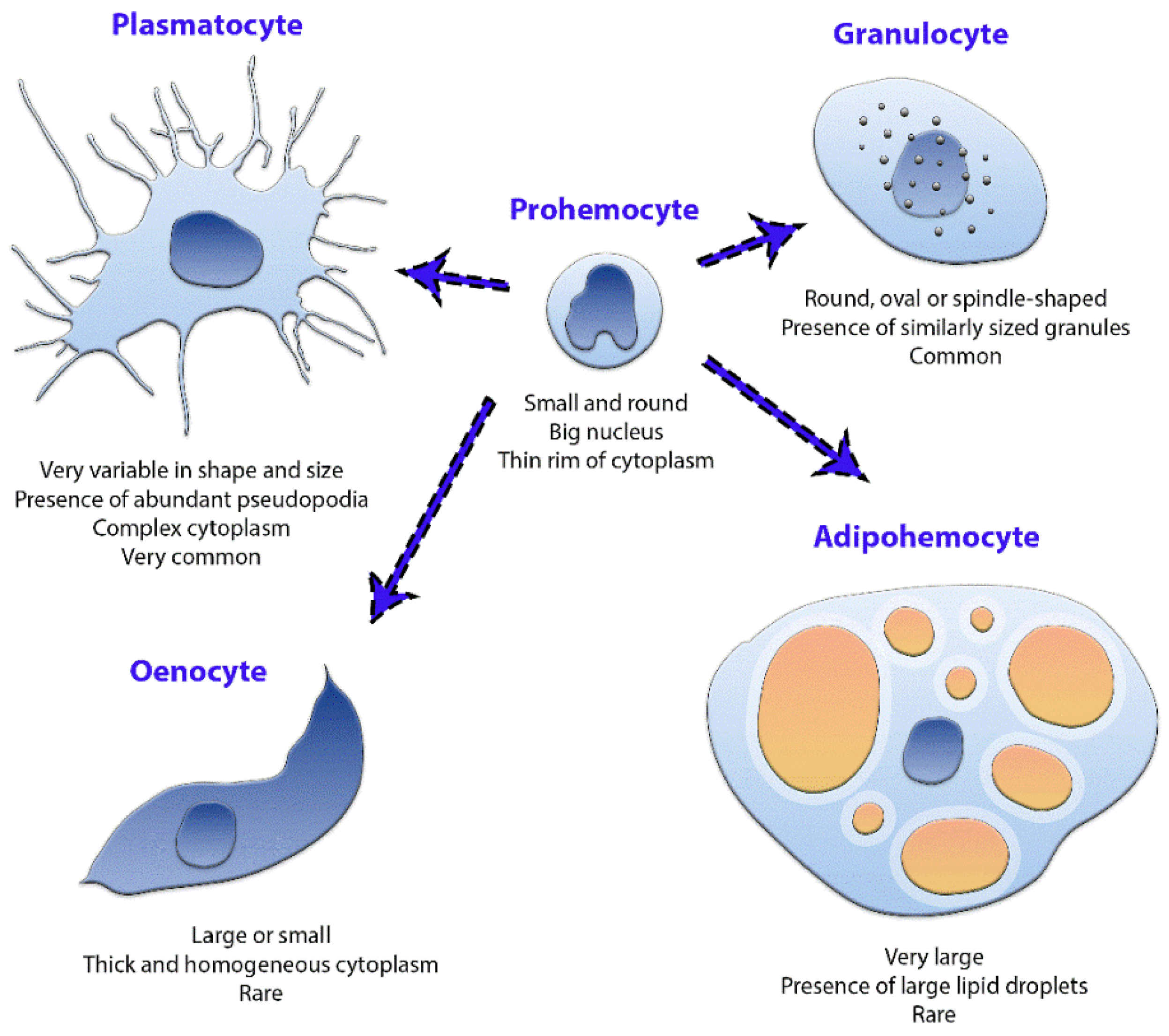
3.1. Phase-Contrast Microscopy
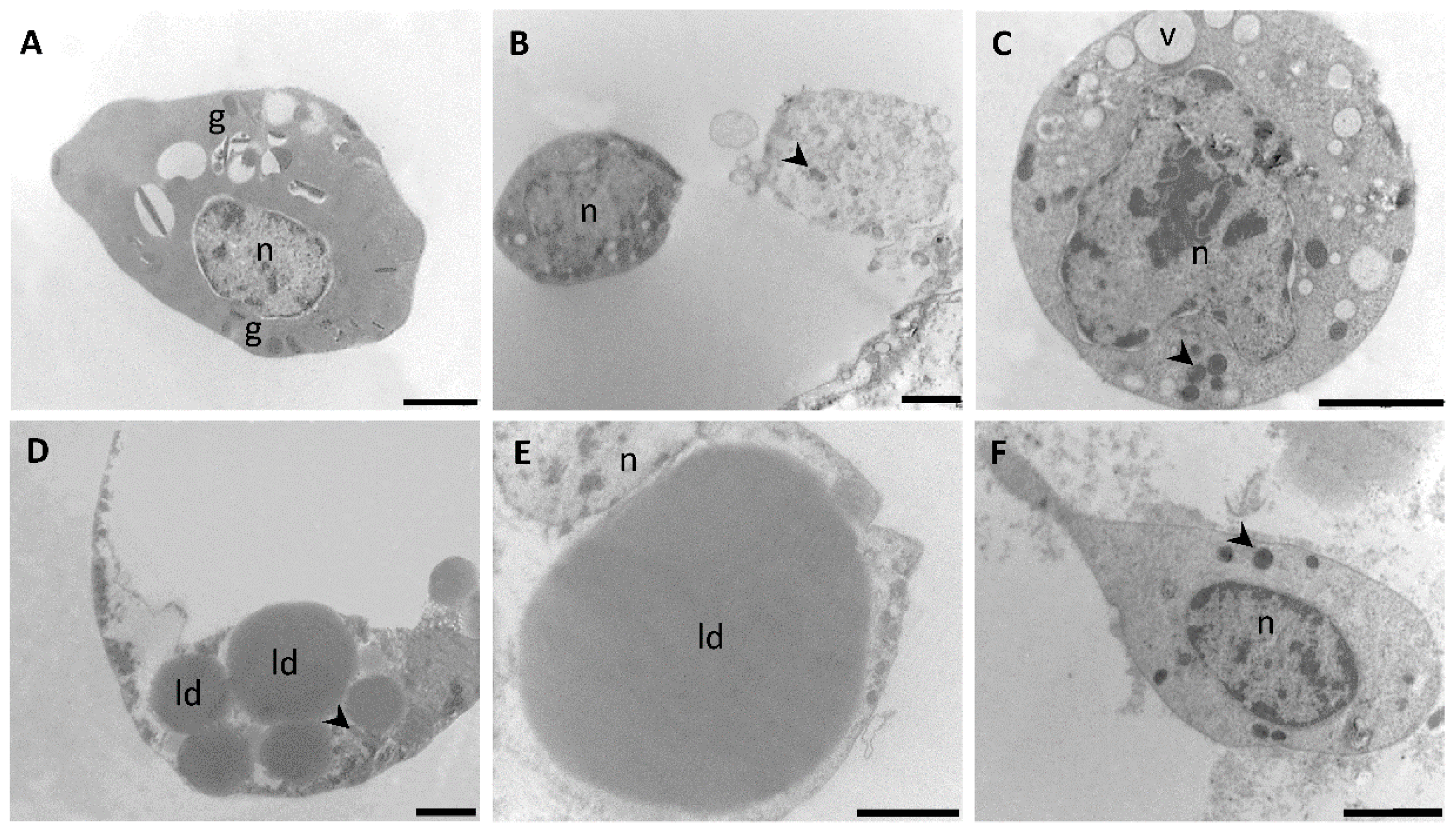
3.2. Transmission Electron Microscopy
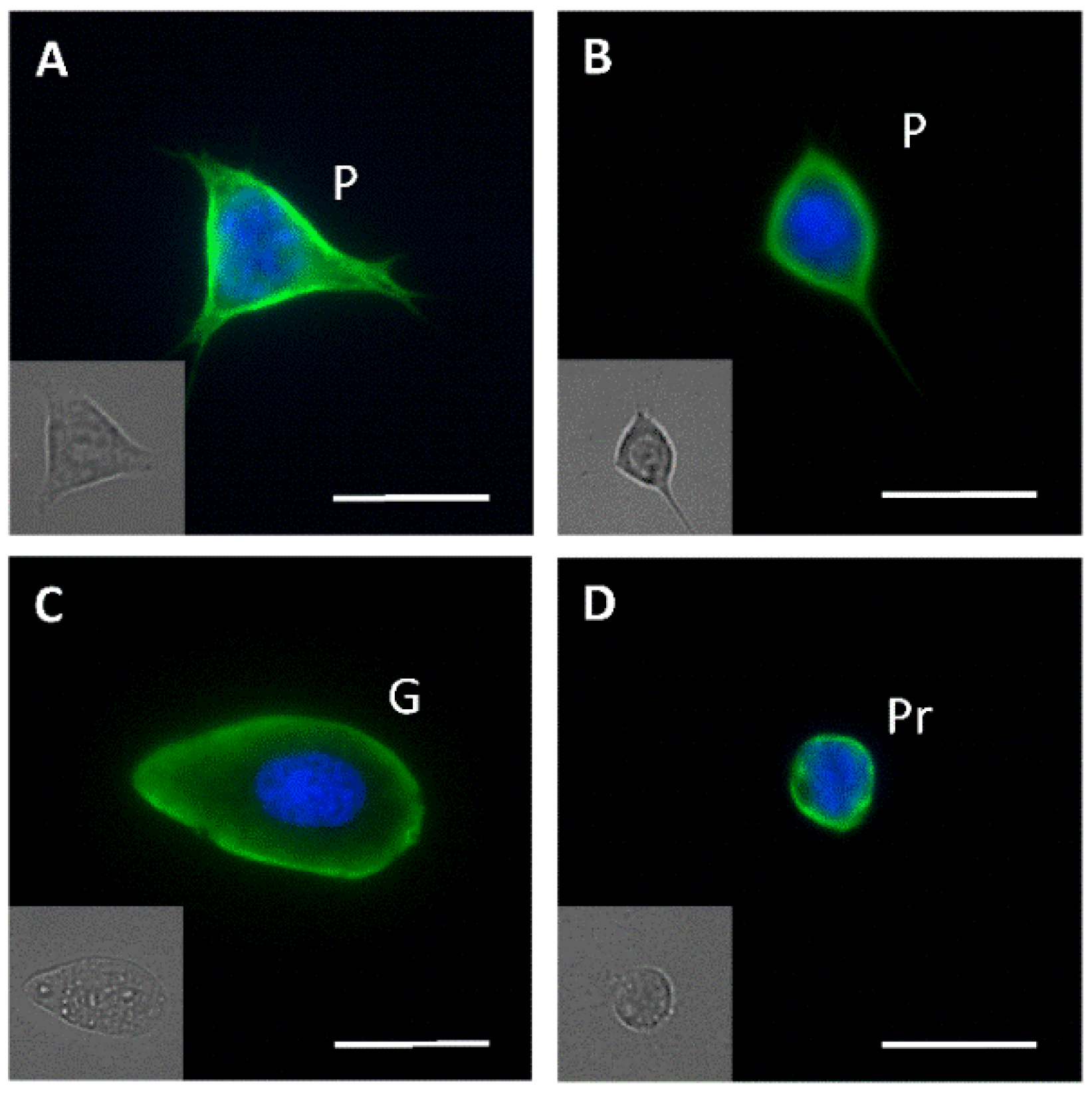
3.3. Fluorescence Microscopy
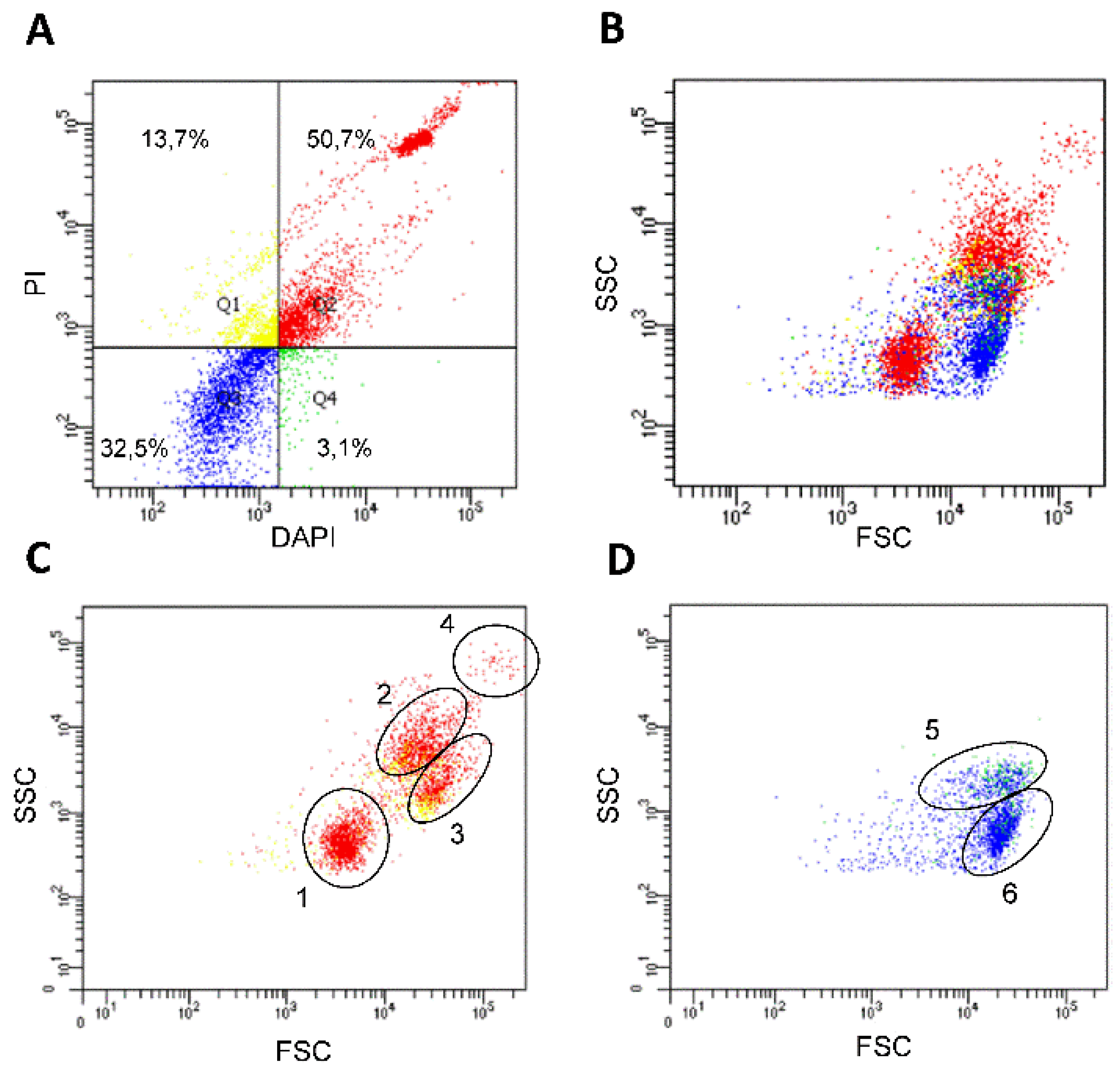
3.4. Flow Cytometry Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Capinera, J.L.; Crist, T.O.; Heppner, J.B.; Tzanakakis, M.E.; Gayubo, S.F.; Tartar, A.; Lawrence, P.O.; Hangay, G.; Shukle, R.H.; Skevington, J.H.; et al. Hemocytes of Insects: Their Morphology and Function. In Encyclopedia of Entomology; Springer: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Gupta, A.P. Cellular Elements in the Hemolymph. In Integument, Respiration and Circulation; Pergamon Press: Oxford, UK, 1985; pp. 402–451. [Google Scholar]
- Pandey, J.P.; Tiwari, R.K. An overview of insect hemocyte science and its future application in applied and biomedical fields. Am. J. Biochem. Mol. Biol. 2012, 2, 82–105. [Google Scholar] [CrossRef] [Green Version]
- Browne, N.; Heelan, M.; Kavanagh, K. An analysis of the structural and functional similarities of insect hemocytes and mammalian phagocytes. Virulence 2013, 4, 597–603. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.; Cho, S. Hemocyte-hemocyte adhesion by granulocytes is associated with cellular immunity in the cricket, Gryllus bimaculatus. Sci. Rep. 2019, 9, 18066. [Google Scholar] [CrossRef] [Green Version]
- Miles, M.A. New World Trypanosomiasis. In Topley & Wilson’s Microbiology and Microbial Infections; Wiley: Hoboken, NJ, USA, 2010. [Google Scholar]
- World Health Organisation. Chagas Disease (also Known as American Trypanosomiasis). Available online: https://www.who.int/news-room/fact-sheets/detail/chagas-disease-(american-trypanosomiasis) (accessed on 19 May 2021).
- Guzmán-Bracho, C. Epidemiology of Chagas disease in Mexico: An update. Trends Parasitol. 2001, 17, 372–376. [Google Scholar] [CrossRef]
- Noireau, F.; Carbajal-De-La-Fuente, A.L.; Lopes, C.M.; Diotaiuti, L. Some considerations about the ecology of Triatominae. An. Acad. Bras. Cienc. 2005, 77, 431–436. [Google Scholar] [CrossRef] [Green Version]
- Bello-Bedoy, R.; Peiro-Nuño, H.; Córdoba-Aguilar, A.; Flores-López, C.A.; Romero-Figueroa, G.; Arteaga, M.C.; Gutiérrez-Cabrera, A.E.; De la Rosa-Conroy, L. Ontogenetic changes in wild chagasic bugs (Dipetalogaster maximus): Exploring morphological adaptations in pre-adult and adult stages. Rev. Mex. Biodivers. 2019, 90, e902664. [Google Scholar] [CrossRef]
- Canavoso, L.E.; Rubiolo, E.R. Interconversions of lipophorin particles by adipokinetic hormone in hemolymph of Panstrongylus megistus, Dipetalogaster maximus and Triatoma infestans (Hemiptera: Reduviidae). Comp. Biochem. Physiol. Part A Physiol. 1995, 112, 143–150. [Google Scholar] [CrossRef]
- Aguirre, S.A.; Fruttero, L.L.; Leyria, J.; Defferrari, M.S.; Pinto, P.M.; Settembrini, B.P.; Rubiolo, E.R.; Carlini, C.R.; Canavoso, L.E. Biochemical changes in the transition from vitellogenesis to follicular atresia in the hematophagous Dipetalogaster maxima (Hemiptera: Reduviidae). Insect Biochem. Mol. Biol. 2011, 41, 832–841. [Google Scholar] [CrossRef] [PubMed]
- Leyria, J.; Fruttero, L.L.; Ligabue-Braun, R.; Defferrari, M.S.; Arrese, E.L.; Soulages, J.L.; Settembrini, B.P.; Carlini, C.R.; Canavoso, L.E. DmCatD, a cathepsin D-like peptidase of the hematophagous insect Dipetalogaster maxima (Hemiptera: Reduviidae): Purification, bioinformatic analyses and the significance of its interaction with lipophorin in the internalization by developing oocytes. J. Insect Physiol. 2018, 105, 28–39. [Google Scholar] [CrossRef]
- Ramos, F.; Leyria, J.; Nouzova, M.; Fruttero, L.; Noriega, F.; Canavoso, L. Juvenile hormone mediates lipid storage in the oocytes of Dipetalogaster maxima. Insect Biochem. Mol. Biol. 2020, 133, 103499. [Google Scholar] [CrossRef]
- Moyetta, N.R.; Fruttero, L.L.; Leyria, J.; Ramos, F.O.; Carlini, C.R.; Canavoso, L. The entomotoxin Jack Bean Urease changes cathepsin D activity in nymphs of the hematophagous insect Dipetalogaster maxima (Hemiptera: Reduviidae). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2021, 251, 110511. [Google Scholar] [CrossRef]
- Leyria, J.; Fruttero, L.L.; Nazar, M.; Canavoso, L.E. The role of DmCatD, a cathepsin D-like peptidase, and acid phosphatase in the process of follicular atresia in Dipetalogaster maxima (Hemiptera: Reduviidae), a vector of chagas’ disease. PLoS ONE 2015, 10, e0130144. [Google Scholar] [CrossRef] [Green Version]
- Padilha, M.H.P.; Pimentel, A.C.; Ribeiro, A.F.; Terra, W.R. Sequence and function of lysosomal and digestive cathepsin D-like proteinases of Musca domestica midgut. Insect Biochem. Mol. Biol. 2009, 39, 782–791. [Google Scholar] [CrossRef] [PubMed]
- Borges, E.C.; Machado, E.M.M.; Garcia, E.S.; Azambuja, P. Trypanosoma cruzi: Effects of infection on cathepsin D activity in the midgut of Rhodnius prolixus. Exp. Parasitol. 2006, 112, 130–133. [Google Scholar] [CrossRef] [PubMed]
- Saikhedkar, N.; Summanwar, A.; Joshi, R.; Giri, A. Cathepsins of lepidopteran insects: Aspects and prospects. Insect Biochem. Mol. Biol. 2015, 64, 51–59. [Google Scholar] [CrossRef] [PubMed]
- de Azambuja, P.; Garcia, E.S.; Ratcliffe, N.A. Aspects of classification of Hemiptera hemocytes from six triatomine species. Mem. Inst. Oswaldo Cruz 1991, 86, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barracco, M.A.; De Oliveira, R.; Schlemper Júnior, B. The hemocytes of Panstrongylus megistus (Hemiptera: Reduviidae). Mem. Inst. Oswaldo Cruz 1987, 82, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Barracco, M.A.; Loch, C.T. Ultrastructural studies of the hemocytes of Panstrongylus megistus (Hemiptera: Reduviidae). Mem. Inst. Oswaldo Cruz 1989, 84, 171–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.C. The hemocytes of Rhodnius prolixus Stal. Biol. Bull. 1965, 129, 282–294. [Google Scholar] [CrossRef]
- Wigglesworth, V.B. Memoirs: The physiology of the cuticle and of ecdysis in Rhodnius prolixus (Triatomidae, Hemiptera); with special reference to the function of the oenocytes and of the dermal glands. J. Cell Sci. 1933, 2, 269–318. [Google Scholar] [CrossRef]
- Wigglesworth, V.B. The Principles of Insect Physiology; Springer: Berlin/Heidelberg, Germany, 1972. [Google Scholar]
- Wigglesworth, V.B. The role of the haemocytes in the growth and moulting of an insect, Rhodnius prolixus (Hemiptera). J. Exp. Biol. 1955, 32, 649–663. [Google Scholar] [CrossRef]
- Wigglesworth, V.B. The function of the amoebocytes during moulting in Rhodnius. Ann. Des. Sci. Nat. Zool. 1956, 18, 139–144. [Google Scholar]
- Wigglesworth, V.B. The haemocytes and connective tissue formation in an insect, Rhodnius prolixus (Hemiptera). J. Cell Sci. 1956, 3, 87–98. [Google Scholar]
- Nuñez, J.A.; Segura, E.L. Rearing of Triatominae. In Chagas’ Disease Vectors. Volume II. Anatomic and Physiological Aspects; Springer: Berlin/Heidelberg, Germany, 1987; pp. 31–40. [Google Scholar]
- Moyetta, N.R.; Broll, V.; Perin, A.P.A.; Uberti, A.F.; Coste Grahl, M.V.; Staniscuaski, F.; Carlini, C.R.; Fruttero, L.L. Jaburetox-induced toxic effects on the hemocytes of Rhodnius prolixus (Hemiptera: Reduviidae). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017, 200, 17–26. [Google Scholar] [CrossRef]
- Bergin, D.; Reeves, E.P.; Renwick, J.; Wientjes, F.B.; Kavanagh, K. Superoxide production in Galleria mellonella hemocytes: Identification of proteins homologous to the NADPH oxidase complex of human neutrophils. Infect Immun. 2005, 73, 4161–4170. [Google Scholar] [CrossRef] [Green Version]
- Senior, N.J.; Titball, R.W. Isolation and primary culture of Galleria mellonella hemocytes for infection studies. F1000Res. 2020, 9, 1392. [Google Scholar] [CrossRef] [PubMed]
- Fruttero, L.L.; Leyria, J.; Moyetta, N.R.; Ramos, F.O.; Settembrini, B.P.; Canavoso, L.E. The fat body of the hematophagous insect, Panstrongylus megistus (Hemiptera: Reduviidae): Histological features and participation of the β-chain of ATP synthase in the lipophorin-mediated lipid transfer. J. Insect Sci. 2019, 19, 16. [Google Scholar] [CrossRef] [PubMed]
- Lane, N.J.; Leslie, R.A.; Swales, L.S. Insect peripheral nerves: Accessibility of neurohaemal regions to lanthanum. J. Cell Sci. 1975, 18, 179–197. [Google Scholar] [CrossRef]
- Price, C.D.; Ratcliffe, N.A. A reappraisal of insect haemocyte classification by the examination of blood from fifteen insect orders. Zeitschrift Für Zellforsch. Mikroskopische Anat. 1974, 147, 537–549. [Google Scholar] [CrossRef]
- Ratcliffe, N.A.; Rowley, A.F.; Fitzgerald, S.W.; Rhodes, C.P. Invertebrate Immunity: Basic Concepts and Recent Advances. Int. Rev. Cytol. 1985, 97, 183–350. [Google Scholar] [CrossRef]
- Lai-Fook, J. Haemocytes in the repair of wounds in an insect (Rhodnius prolixus). J. Morphol. 1970, 130, 297–313. [Google Scholar] [CrossRef] [PubMed]
- Lavine, M.D.; Strand, M.R. Insect hemocytes and their role in immunity. Insect Biochem. Mol. Biol. 2002, 32, 1295–1309. [Google Scholar] [CrossRef]
- Jiang, H.; Vilcinskas, A.; Kanost, M.R. Immunity in lepidopteran insects. Adv. Exp. Med. Biol. 2010, 708, 181–204. [Google Scholar] [CrossRef]
- Marringa, W.J.; Krueger, M.J.; Burritt, N.L.; Burritt, J.B. Honey bee hemocyte profiling by flow cytometry. PLoS ONE 2014, 9, e108486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Léry, X.; Charpentier, G.; Belloncik, S. DNA content analysis of insect cell lines by flow cytometry. Cytotechnology 1999, 29, 103–113. [Google Scholar] [CrossRef]
- Panzera, F.; Ferrandis, I.; Ramsey, J.; Salazar-Schettino, P.M.; Cabrera, M.; Monroy, C.; Bargues, M.D.; Mas-Coma, S.; O’Connor, J.E.; Angulo, V.M.; et al. Genome size determination in Chagas disease transmitting bugs (Hemiptera-Triatominae) by flow cytometry. Am. J. Trop. Med. Hyg. 2007, 76, 516–521. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moyetta, N.R.; Ramos, F.O.; Leyria, J.; Canavoso, L.E.; Fruttero, L.L. Morphological and Ultrastructural Characterization of Hemocytes in an Insect Model, the Hematophagous Dipetalogaster maxima (Hemiptera: Reduviidae). Insects 2021, 12, 640. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12070640
Moyetta NR, Ramos FO, Leyria J, Canavoso LE, Fruttero LL. Morphological and Ultrastructural Characterization of Hemocytes in an Insect Model, the Hematophagous Dipetalogaster maxima (Hemiptera: Reduviidae). Insects. 2021; 12(7):640. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12070640
Chicago/Turabian StyleMoyetta, Natalia R., Fabián O. Ramos, Jimena Leyria, Lilián E. Canavoso, and Leonardo L. Fruttero. 2021. "Morphological and Ultrastructural Characterization of Hemocytes in an Insect Model, the Hematophagous Dipetalogaster maxima (Hemiptera: Reduviidae)" Insects 12, no. 7: 640. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12070640