Timing Is Everything. Temporal and Spatial Niche Segregation in Curculio spp. (Coleoptera: Curculionidae) Associated with Oak Trees



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experiment 1—Ovipositional Strategies among Curculio Species
- differences between masses of acorns chosen by particular Curculio spp. females;
- differences in the number of eggs laid by Curculio spp. females.
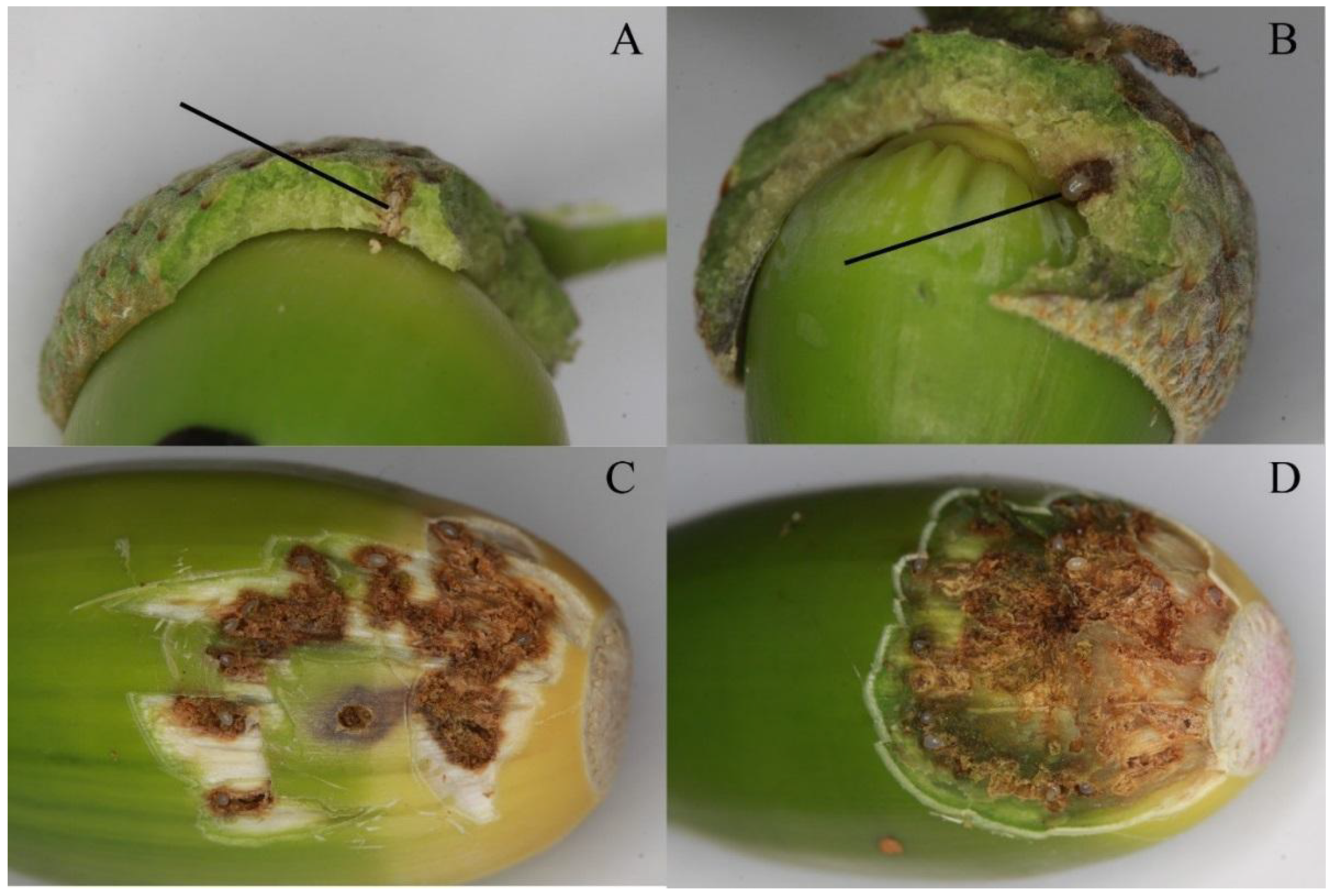
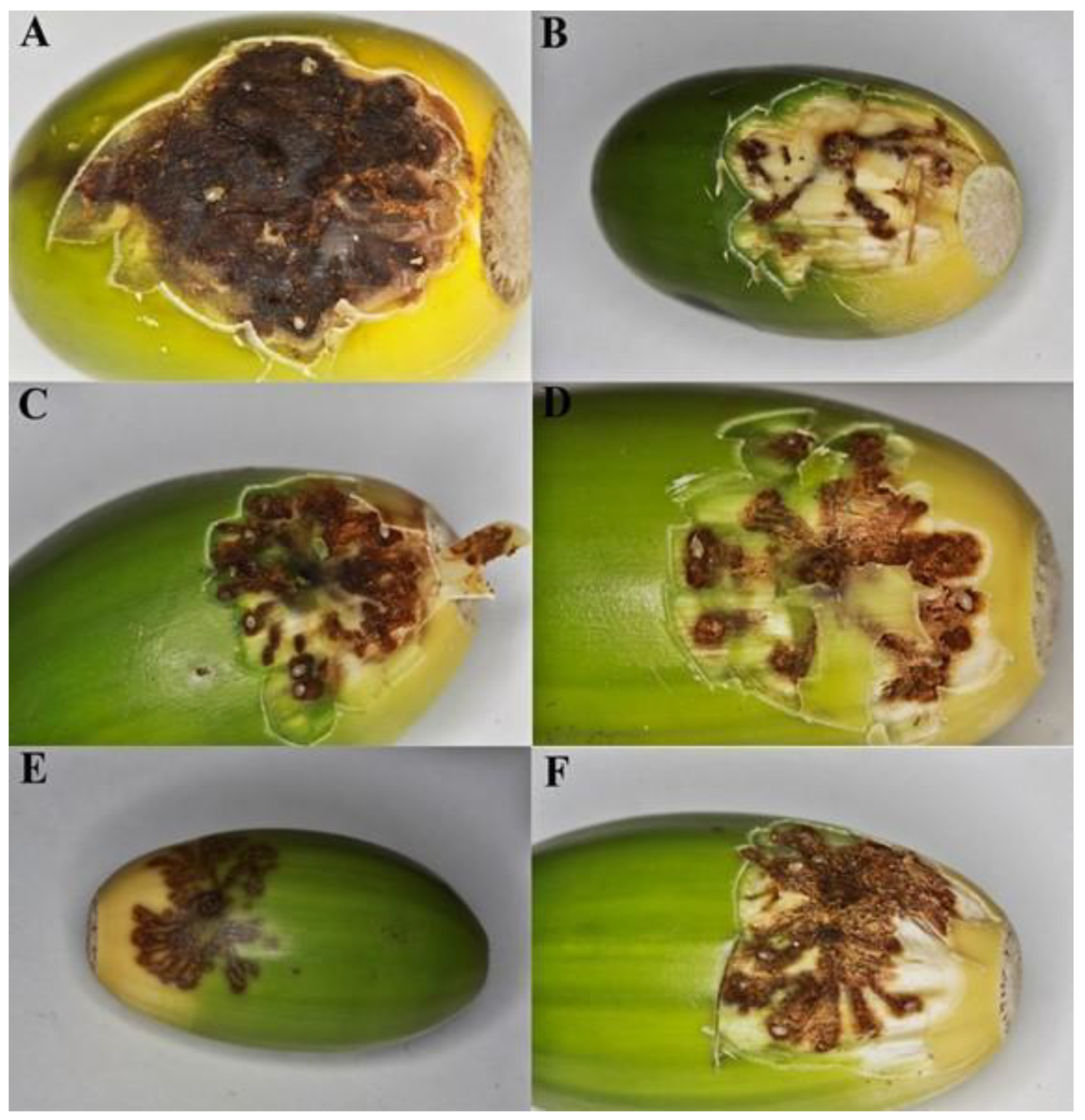
2.2. Experiment 2—Distribution of Eggs and Egg Galleries in Curculio Glandium
3. Results
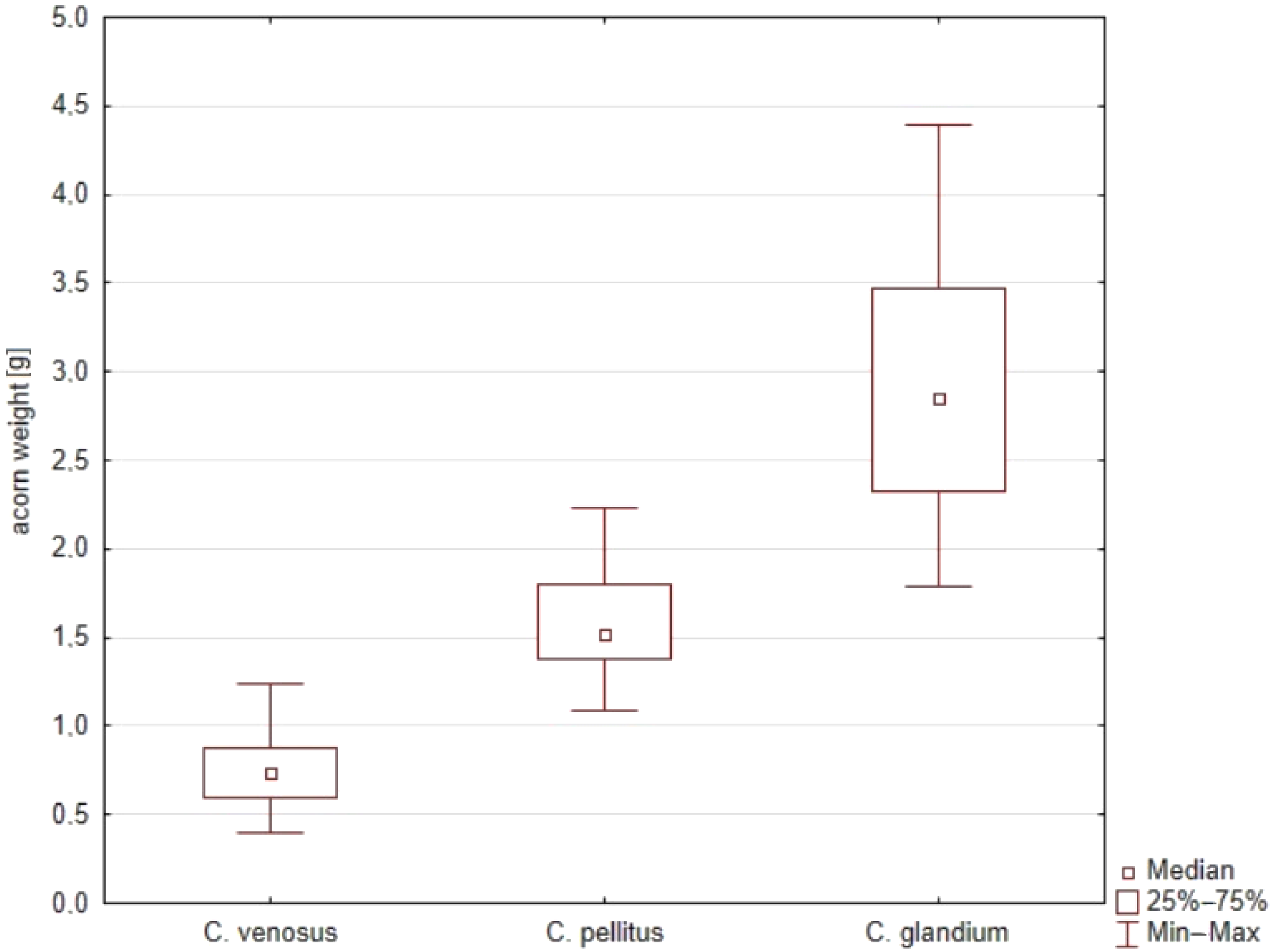
3.1. Ovipositional Strategies among Three Curculio Species
3.2. Distribution of Eggs and Egg Galleries in Curculio Glandium
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Janzen, D.H. Specificity of seed-attacking beetles in a Costa Rican deciduous forest. J. Ecol. 1980, 68, 929–952. [Google Scholar] [CrossRef]
- Novotny, V.; Basset, Y. Host specificity of insect herbivores in tropical forests. Proc. R. Soc. B. 2005, 272, 1083–1090. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.C.; Gao, P.; Zhang, G.Y.; Wen, C.; Wen, J.B. Micro-habitat niche differentiation contributing to coexistence of Eucryptorrhynchus scrobiculatus Motschulsky and Eucryptorrhynchus brandti (Harold). Biocontrol. Sci. Technol. 2017, 27, 1180–1194. [Google Scholar] [CrossRef]
- Muñoz, A.; Bonal, R.; Espelta, J.M. Acorn–weevil interactions in a mixed-oak forest: Outcomes for larval growth and plant recruitment. Forest Ecol. Manag. 2014, 322, 98–105. [Google Scholar] [CrossRef]
- Addicott, F.T. Abscission; University of California Press: Berkeley, CA, USA, 1982; pp. 1–355. [Google Scholar]
- Koenig, W.D.; Knops, J.M. The behavioral ecology of masting in oaks. In Oak Forest Ecosystems: Ecology and Management for Wildlife; Mc Shea, W.J., Healy, W.M., Eds.; Johns Hopkins University Press: Baltimore, MD, USA, 2002; pp. 129–148. [Google Scholar]
- Bogdziewicz, M.; Marino, S.; Bonal, R.; Zwolak, R.; Steele, M.A. Rapid aggregative and reproductive responses of weevils to masting of North American oaks counteract predator satiation. Ecology 2018, 99, 2575–2582. [Google Scholar] [CrossRef]
- Bogdziewicz, M.; Espelta, J.M.; Muñoz, A.; Aparicio, J.M.; Bonal, R. Effectiveness of predator satiation in masting oaks is negatively affected by conspecific density. Oecologia 2018, 186, 983–993. [Google Scholar] [CrossRef] [Green Version]
- Canelo, T.; Gaytán, Á.; Pérez-Izquierdo, C.; Bonal, R. Effects of Longer Droughts on Holm Oak Quercus ilex L. Acorn Pests: Consequences for Infestation Rates, Seed Biomass and Embryo Survival. Diversity 2021, 13, 110. [Google Scholar] [CrossRef]
- Den Ouden, J.P.; Jansen, A.; Smit, R. Jays, mice and oaks: Predation and dispersal of Quercus robur and Q. petraea in North-western Europe. In Seed Fate: Predation, Dispersal and Seedling Establishment; Forget, P.M., Lambert, J.E., Hulme, P.E., Vander Wall, S.B., Eds.; CABI Publishing: Wallingford, UK, 2005; pp. 223–240. [Google Scholar]
- Bonal, R.; Muñoz, A. Multi-trophic effects of ungulate intraguild predation on acorn weevils. Oecologia 2007, 152, 533–540. [Google Scholar] [CrossRef]
- Canelo, T.; Perez-Izquierdo, C.; Gaytan, A.; Bonal, R. Intraguild predation of weevils by livestock reduces acorn pests in oak silvopastoral systems. J. Pest. Sci. 2021, 94, 541–551. [Google Scholar] [CrossRef]
- Bonal, R.J.; Espelta, M.; Vogler, A.P. Complex selection on life-history traits and the maintenance of variation in exaggerated rostrum length in acorn weevils. Oecologia 2011, 167, 1053–1061. [Google Scholar] [CrossRef]
- Wanat, M.; Mokrzycki, T. The Checklist of the Weevils (Coleoptera: Curculionoidea) of Poland Revisited. Ann. Zool. 2018, 68, 1–48. [Google Scholar] [CrossRef]
- Kania, J.; Malkiewicz, A.; Niedojad, K.; Matraj, M. Uwagi o biologii i występowaniu Curculio elephas (Gyllenhal, 1836) (Coleoptera: Curculionidae) w Polsce południowo-zachodniej. Przyroda Sudetów 2013, 16, 119–124. [Google Scholar]
- Hughes, J.; Vogler, A.P. Ecomorphological adaptation of acorn weevils to their oviposition site. Evolution 2004, 58, 1971–1983. [Google Scholar] [CrossRef] [PubMed]
- Pélisson, P.F.; Bel Venner, M.C.; Rey, B.; Burgevin, L.; Martineau, F.; Fourel, F.; Venner, S. Contrasted breeding strategies in four sympatric sibling insect species: When a proovigenic and capital breeder copes with a stochastic environment. Funct. Ecol. 2012, 26, 198–206. [Google Scholar] [CrossRef]
- Aoki, K.; Kato, M.; Murakami, N. Glacial bottleneck and postglacial recolonization of a seed parasitic weevil, Curculio hilgendorfi, inferred from mitochondrial DNA variation. Mol. Ecol. 2008, 17, 3276–3289. [Google Scholar] [CrossRef] [PubMed]
- Aoki, K.; Kato, M.; Murakami, N. Phylogeography of Phytophagous Weevils and Plant Species in Broadleaved Evergreen Forests: A Congruent Genetic Gap between Western and Eastern Parts of Japan. Insects 2011, 2, 128–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sprick, P.; Floren, A. Diversity of Curculionoidea in Humid Rain Forest Canopies of Borneo: A Taxonomic Blank Spot. Diversity 2018, 10, 116. [Google Scholar] [CrossRef] [Green Version]
- Smreczyński, S. The Key for Identification of Polish Insects. Beetles–Coleoptera. Part XIX (98f). Weevils–Curculionidae. Subfamily–Curculioninae; Państwowe Wydawnictwo Naukowe: Warsaw, Poland, 1972; p. 195. [Google Scholar]
- Peguero, G.R.; Bonal, R.; Sol, D.; Muñoz, A.; Sork, V.L.; Espelta, J.M. Tropical insect diversity: Evidence of greater host specialization in seed-feeding weevils. Ecology 2017, 98, 2180–2190. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.-Y.; Hu, X.-H.; Liu, D.-C.; Ouyang, A.; Tong, X.; Wang, Y.-J.; Wang, R.; Chen, X.-Y. High diversity and strong variation in host specificity of seed parasitic acorn weevils. Insect Conserv. Divers. 2020, 14, 367–376. [Google Scholar] [CrossRef]
- Bonal, R.; Muñoz, A. Seed weevils living on the edge: Pressures and conflicts over body size in the endoparasitic Curculio larvae. Ecol. Entomol. 2009, 34, 304–309. [Google Scholar] [CrossRef]
- Addesso, K.M.; Alborn, H.T.; Bruton, R.R.; McAuslane, H.J. A multicomponent marking pheromone produced by the pepper weevil, Anthonomus eugenii (Coleoptera: Curculionidae). Chemoecology 2021, 1–12. [Google Scholar] [CrossRef]
- Addesso, K.A.; McAuslane, H.J.; Stansly, P.A.; Schuster, D.J. Host-marking by female pepper weevils, Anthonomus eugenii. Entomol. Exp. Appl. 2007, 125, 269–327. [Google Scholar] [CrossRef]
- Okamoto, M. Interactions between Camellia japonica and its seed predator Curculio camelliae I. Observations on morphology, phenology and oviposition behaviors in Kinki district, Japan. Bull. Osaka Mus. Nat. Hist. 1988, 43, 15–37. [Google Scholar]
- Haack, R.A.; Wilkinson, R.C.; Foltz, J.L.; Corneil, J.A. Gallery construction and oviposition by Ips calligraphus (Coleoptera: Scolytidae) in relation to slash pine phloem thickness and temperature. Can. Entomol. 1984, 116, 625–632. [Google Scholar] [CrossRef]
- Shaw, M.; Jennings, M.; Quicke, D. The identity of Scambus planatus (Hartig, 1838) and Scambus ventricosus (Tschek, 1871) as seasonal forms of Scambus calobatus (Gravenhorst, 1829) in Europe (Hymenoptera, Ichneumonidae, Pimplinae, Ephialtini). J. Hymenopt. Res. 2011, 23, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Hughes, J.; Vogler, A.P. The phylogeny of acorn weevils (genus Curculio) from mitochondrial and nuclear DNA sequences: The problem of incomplete data. Mol. Phylogenetics Evol. 2004, 32, 601–615. [Google Scholar] [CrossRef] [PubMed]
- Toju, H.; Sota, T. Adaptive divergence of scaling relationships mediates the arms race between a weevil and its host plant. Biol. Lett. 2006, 2, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toju, H. Natural selection drives the fine-scale divergence of a coevolutionary arms race involving a long-mouthed weevil and its obligate host plant. BMC Evol. Biol. 2009, 9, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, X.; Yang, Y. Large acorns benefit seedling recruitment by satiating weevil larvae in Quercus aliena. Plant Ecol. 2010, 209, 291–300. [Google Scholar] [CrossRef]
- Yi, X.; Wang, Z.; Liu, C.; Liu, G.; Zhang, M. Acorn cotyledons are larger than their seedlings’ need: Evidence from artificial cutting experiments. Sci. Rep. 2015, 5, 1–6. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | N | Min | Max | Median | Quartile Range |
|---|---|---|---|---|---|
| C. venosus | 34 | 0.394 | 1.241 | 0.741 a | 0.278 |
| C. pellitus | 19 | 1.083 | 2.231 | 1.519 b | 0.425 |
| C. glandium | 50 | 1.782 | 4.388 | 2.852 c | 1.137 |
| Scheme | N | Min | Max | Median | Sum |
|---|---|---|---|---|---|
| C. venosus | 34 | 1 | 3 | 1 a | 44 |
| C. pellitus | 19 | 1 | 3 | 2 a | 38 |
| C. glandium | 50 | 1 | 8 | 4 b | 194 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reut, M.; Chrabąszcz, M.; Moniuszko, H. Timing Is Everything. Temporal and Spatial Niche Segregation in Curculio spp. (Coleoptera: Curculionidae) Associated with Oak Trees. Insects 2021, 12, 687. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12080687
Reut M, Chrabąszcz M, Moniuszko H. Timing Is Everything. Temporal and Spatial Niche Segregation in Curculio spp. (Coleoptera: Curculionidae) Associated with Oak Trees. Insects. 2021; 12(8):687. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12080687
Chicago/Turabian StyleReut, Michał, Mariusz Chrabąszcz, and Hanna Moniuszko. 2021. "Timing Is Everything. Temporal and Spatial Niche Segregation in Curculio spp. (Coleoptera: Curculionidae) Associated with Oak Trees" Insects 12, no. 8: 687. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12080687