
Design and Testing of Effective Primers for Amplification of the orf7 Gene of Phage WO Associated with Andricus hakonensis


1
Laboratory of Insect Behavior and Evolutionary Ecology, College of Life Science and Technology, Central South University of Forestry & Technology (CSUFT), Changsha 410004, China
2
College of Life Science, Hunan Normal University, Changsha 410081, China
*
Author to whom correspondence should be addressed.
Insects 2021, 12(8), 713; https://0-doi-org.brum.beds.ac.uk/10.3390/insects12080713
Submission received: 11 May 2021
/
Revised: 17 July 2021
/
Accepted: 24 July 2021
/
Published: 9 August 2021
Abstract
:Simple Summary
Andricus hakonensis is thought to contain the most complex and diverse phage types known and should be an ideal model material for studying interactions among bacteriophages, bacteria and eukaryotes. As shown in previous studies, existing primers are not effective enough to amplify all virus groups in A. hakonensis. Based on a comprehensive analysis of all virus groups reported to date, we designed a relatively conservative primer for virus detection. This primer can accurately and efficiently detect the presence of phage WO in arthropod hosts. Using gene alignment, clear evidence was provided for the existence of hitherto unreported base deletions, which are an important cause of diversity in phage WO associated with A. hakonensis.
Abstract
Phage WO was first characterized in Wolbachia, an obligate intracellular Rickettsiales known for its ability to regulate the reproduction of arthropod hosts. In this paper, we focus on the study of virus diversity in Andricus hakonensis and the development of highly effective primers. Based on the existing Wolbachia genome sequence, we designed primers (WO-TF and WO-TR) to amplify the full-length orf7 gene of phage WO. Surprisingly, sequencing results showed a high abundance of other phage WO groups in A. hakonensis, in addition to the four groups previously identified. The results also showed that A. hakonensis contained most of the known types of orf7 genes (I, III, IV, V and VI) and the level of diversity of harbored phage WO was very high. Therefore, we speculated that existing primers were not specific enough and that new primers for the detection of phage WO were needed. Based on the existing orf7 gene sequence, we designed specific detection primers (WO-SUF and WO-SUR). Sequencing results showed that the primers effectively amplified all known types of phage WO. In addition to amplifying most of the known sequences, we also detected some new genotypes in A. hakonensis using the new primers. Importantly, all phage WO groups could be efficiently detected. Combined with the results of previous studies, our results suggest that A. hakonensis contains the largest number of phage types (up to 36 types). This study is novel in that it provides practical molecular evidence supporting base deletions, in addition to gene mutations and genetic recombination, as an important cause of phage WO diversity.
1. Introduction
Wolbachia are obligated intracellular symbionts that are found in many filarial nematodes and arthropods [1] and are considered to infect about half of arthropod species in the biosphere [2,3]. Following phylogenetic analysis based on the Wolbachia surface protein (wsp) gene and the conserved coding gene 16S ribosomal RNA, strains are classified into 18 different supergroups (A–R) [4]. The high prevalence of Wolbachia is closely related to its ability to induce a variety of phenotypes, from regulating the reproduction of its arthropod hosts through mechanisms such as cytoplasmic incompatibility [5], parthenogenesis [6], male-killing [7] and feminization [8], to mutualism in nematodes [1]. Bacteriophages are thought to be the most abundant group of organisms in the biosphere [9]. Phage WO, the bacteriophage of obligated intracellular symbionts Wolbachia, plays a significant role in Wolbachia genomic evolution. Several recent studies have confirmed that the phage WO gene segment is crucial for cytoplasmic incompatibility of Drosophila melanogaster, induced by the Wolbachia associated with it [10,11,12]. The putative capsid protein encoded orf7 gene is often used to identify bacteriophage infections and for phylogenetic analysis. Polymerase chain reaction (PCR) amplification using the universal primers (WO-F and WO-R) has shown that the phage WO gene segment occurs in most cases of the parasitic A and B Wolbachia supergroups [13,14]. Moreover, 85% of phage-infected Wolbachia strains harbors less than two phage types [14,15]. To the best of our knowledge, and ricus hakonensis contains the largest number of phage types; phage WO detection using WOF/R primers revealed 27 phage types [16].
Gall wasps (Hymenoptera: Cynipidae) are a phytophagous population of the superfamily Cynipoidea that represent the most spectacular branch of gall-inducing insects, with about 1400 species described [17]. It contains twelve tribes, i.e., Cynipini, Aylacini, Eschatocerin, Diplolepidini, Pediaspidili, Qwaqwaiini, Paraulacini, Aulacideini, Phanacidini, Diastrophini, Ceroptresini and Synergini [17]. A. hakonensis of the tribe Cynipini is widely distributed in southern China and it induces galls on Quercus fabri buds and leaves, respectively. We collected them from five places in Anhui, Hunan, Zhejiang and Fujian provinces in early May [16]. Several studies have detected high rates of Wolbachia infections in diverse cynipid species [18,19,20,21,22,23,24]. Moreover, some cynipid species are infected with multiple Wolbachia strains [22].
The tripartite temperate bacteriophage WO, the intracellular bacterium Wolbachia and their eukaryotic hosts are an ideal model system for studying interactions among bacteriophages, bacteria and eukaryotes [25]. In the Wolbachia genome, phage WO is a uniquely described transposable element [26] and prophage genomic regions can occupy more than twenty percent of mobile DNA genes. Moreover, prophage regions play a dominant role in the absent/divergent genes between closely related Wolbachia strains [27]. The relationships among the tripartite arthropod–Wolbachia–phage WO system are extremely intricate. Researchers have tried to explore the relationship between phage WO and Wolbachia associated with insect species such as Drosophila melanogaster, Drosophila simulans, Culex pipiens, Nasonia vitripennis, Ephestia kuehniella and Gryllus pennsylvanicus [10,11,12]. No evidence of phylogenetic congruence between phage WO and Wolbachia suggesting that the cross infection of phage WO in different Wolbachia strains is relatively common [13,27,28]. The CG content and codon preferences of phage WO and its Wolbachia host are similar; all the evidence suggests that phage WO have coevolved within Wolbachia for more than 100 million years [28]. Therefore, there must be a complicated relationship among the tripartite arthropod–Wolbachia–phage WO, which is worth exploring with further research.
The distribution of phage WO and the number of phage types in host arthropods may be greater than current estimates, as the primers WO-F and WO-R used to detect the presence of phage WO are not degenerate enough to detect every orf7 gene variant. For instance, D. simulans was initially reported to have a single phage WO haplotype [14], but the genome sequence of the Wolbachia strain wRi contains four confirmed prophage copies, three of which are unique [29]. Therefore, it is necessary to develop new detection primers of phage WO. By synthetically analyzing the published genome sequences of Wolbachia, we designed specific primers to amplify the full-length sequence of the phage orf7 gene. With our newly designed primers (WO-TF and WO-TR), we detected viral orf7 gene sequences in A. hakonensis that were not detectable using general WO-F and WO-R primers. The results showed that the gall wasp A. hakonensis contained most of the known orf7 gene types (I, III, IV, V, VI and VII) and displayed a very high level of phage WO diversity, which is inconsistent with previous results [14,15]. Given the results we obtained, we decided to develop more versatile primers for the detection of phage WO. The experimental results show that our primers WO-SUF and WO-SUR are more effective than the conventional primers WO-F and WO-R in detecting the bacteriophage WO.
2. Materials and Methods
2.1. Insects and Total Genomic DNA Extraction
Galls of A. hakonensis were collected in early May between 2012 and 2021 from four provinces in southern China. All galls were reared under the appropriate humidity conditions in the zoology laboratory of Central South University of Forestry and Technology (CSUFT) and adult emergence was observed daily. Within 4 days of emergence, adults were directly preserved in 99.5% ethanol at −80 °C until DNA extraction. Adult gall wasps used for RNA extraction were preserved directly in liquid nitrogen.
Adult A. hakonensis were randomly selected for each location and all the samples were washed twice with sterile water before DNA extraction. Total genomic DNA was extracted from each insect using the phenol–chloroform extraction and ethanol precipitation method as previously described [21]. Suitably dried DNA samples were immediately resuspended in 50 µL of sterile ddH2O and deposited in 4 °C refrigerator for later use. We evaluated the quality of each A. hakonensi template by PCR using the nuclear ribosomal DNA internal transcribed spacer 2 (rDNA ITS2) gene [30]. Low quality templates were disposed responsibly to ensure the accuracy of infection rates.
2.2. PCR and Sequencing
All the samples were first screened for Wolbachia infection using the universal primers 81F and 691R (to amplify about 600 bp of Wolbachia surface protein gene) (Table 1) [31]. Phage WO of Wolbachia was then detected using the universal primers WO-F and WO-R (Table 1) to amplify about 350 bp of the Phage WO capsid protein gene orf7 [28]. A blank control (ddH2O) was used for all amplifications. The reaction mixture was composed of 0.5 µL of each primer, 1 µL of template DNA, 0.2 µL of Pyrobest DNA Polymerase (5 U/μL)(Takara Biomedical Technology (Beijing) Co., Ltd, Dalian, China), 2.5 µL of 10×Pyrobest Buffer II, 2 µL of dNTP Mixture (2.5 mM each) and 18.3 µL of sterile ddH2O. The cycling conditions were as follows: 98 °C for 120 s, 30–40 cycles of 98 °C for 10 s, 50–57 °C for 30 s and 68 °C for 1–2 min, using a C1000 Touch Thermal Cycler (Bio-Rad, Hercules, CA, USA).
The PCR products (2.5 μL) were run on a 0.8% Invitrogen agarose gel electrophoresis with 0.3 μL of 10 × Loading Buffer, under 1× TBE buffer. The gels were soaked in GelRed diluent for 30 min and a picture was taken using a gel imaging system (Bio-Rad, Hercules, CA, USA). All positive PCR products were then purified using an Agarose Gel DNA Extraction Kit (Takara Biomedical Technology (Beijing) Co., Ltd.). The purified PCR products were sequenced using the corresponding PCR primers. If the initial sequencing results showed multiple peaks, this sample was considered as multiple phage WO infections. The multiple phage WO infections PCR products were then purified and ligated into the Mighty TA-cloning Reagent Set for PrimeSTAR pMD20-T vector (Takara Biomedical Technology (Beijing) Co., Ltd.) following the manufacturer’s procedures. Firstly, for each adult, 10–15 positive colonies were randomly selected and cultured in a lysogeny broth medium supplemented with ampicillin. Secondly, plasmids were carefully extracted. Finally, we sequenced the plasmid in both directions with two primers (M13F and M13R) using an ABI 3730XL DNA Sequencer (Applied Biosystems, Foster City, CA, USA) at Icongene Co., Ltd. (Wuhan, China).
2.3. Raw Sequence Treatment
Firstly, we performed the sequence homology analysis using the online BLAST program. Secondly, genetic distances between all established sequence pairs were calculated using MEGA 7.0 under Kimura 2-parameter distance models [32]. All the sequencing results in the current study were deposited in GenBank under the following accession numbers: MZ687122-MZ687127; MZ687129-MZ687140; MZ687810-MZ687824.
2.4. Phylogenetic Analyses
The maximum likelihood (ML) phylogenetic tree was carried out using IQ-Tree 2.1.1 via the online CIPRES Science Gateway V. 3.3 using the general time-reversible (GTR) model, in which the invariant sites and gamma distribution were estimated (GTR + I + G) [33]. Model Test v3.7 was used to choose the best evolutionary model. Bootstrap values were generated from 10,000 bootstrap replicates.
2.5. RNA Extraction
We assessed three samples composed of pooled females and three samples of pooled males. The number of individuals in each sample ranged from 10 to 25 according to body size (approximately 10–12 females per sample and 18–25 males per sample). Before RNA extraction, adult A. hakonensis were washed twice with sterile water. We used RNAiso Plus (Takara Biomedical Technology Co., Ltd.) to extract total RNA for each sample and suitably dried RNA samples were resuspended in 50 µL of RNase-free ddH2O. The concentration and purity of each RNA sample were assessed by measuring their optical density (OD) values using a Thermo Scientific NanoDrop One. PCR tests were performed on all RNA samples using the mitochondrial cytochrome c oxidase 1 (cox1) gene [34] and nuclear ribosomal DNA internal transcribed spacer 2 gene [30] as negative controls to affirm that the RNA samples did not contain high levels of A. hakonensis genomic DNA contamination. cDNA was then synthesized using a PrimeScript RT Reagent Kit and remaining genomic DNA contamination was removed using gDNA Eraser (Takara Biomedical Technology Co., Ltd.). The reaction mixture was incubated at 37–50 °C for 40–90 min using a C1000 Touch Thermal Cycler.
3. Results
3.1. Acquisition and Typing of the Full-Length Sequence of the Phage WO orf7 Gene
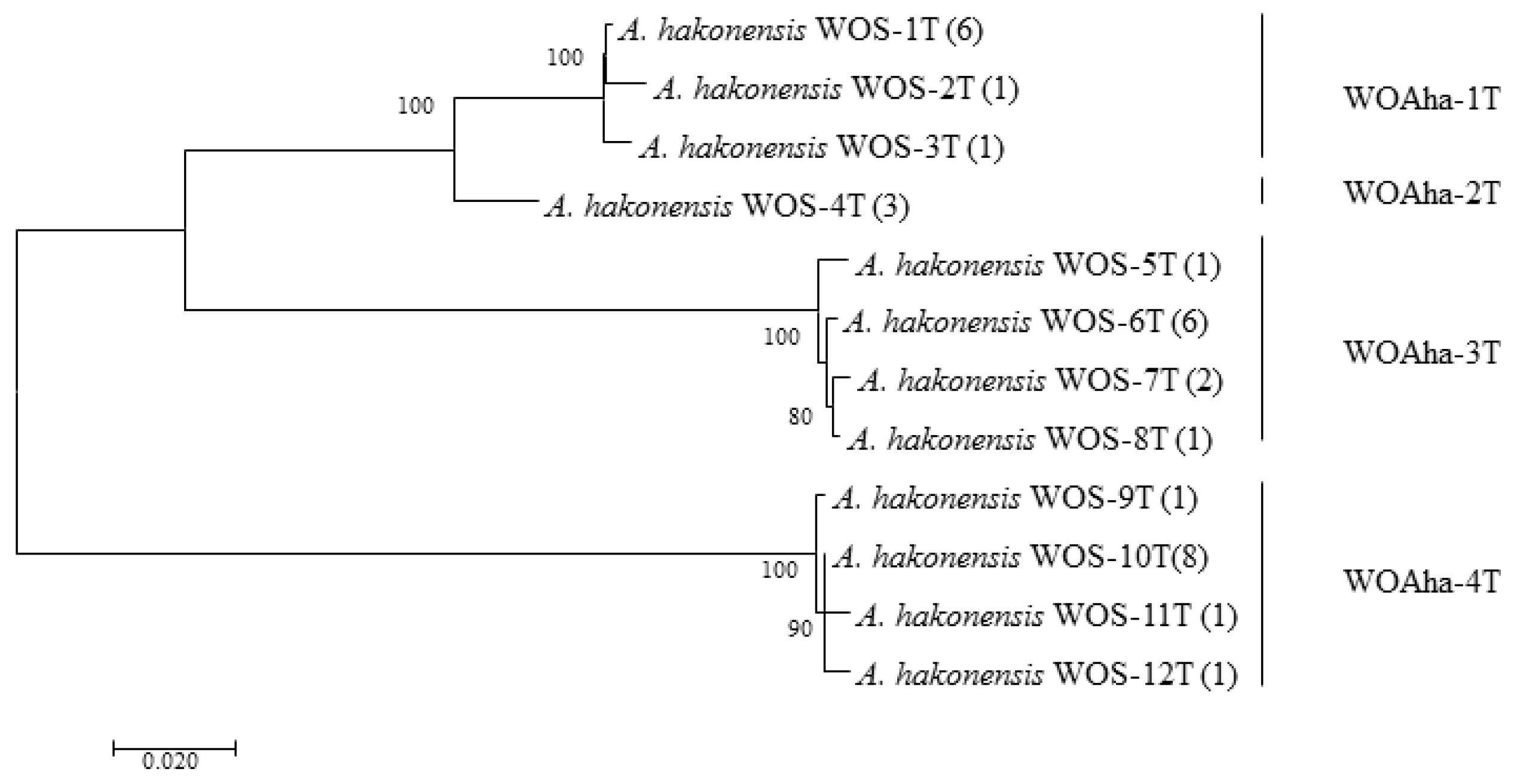
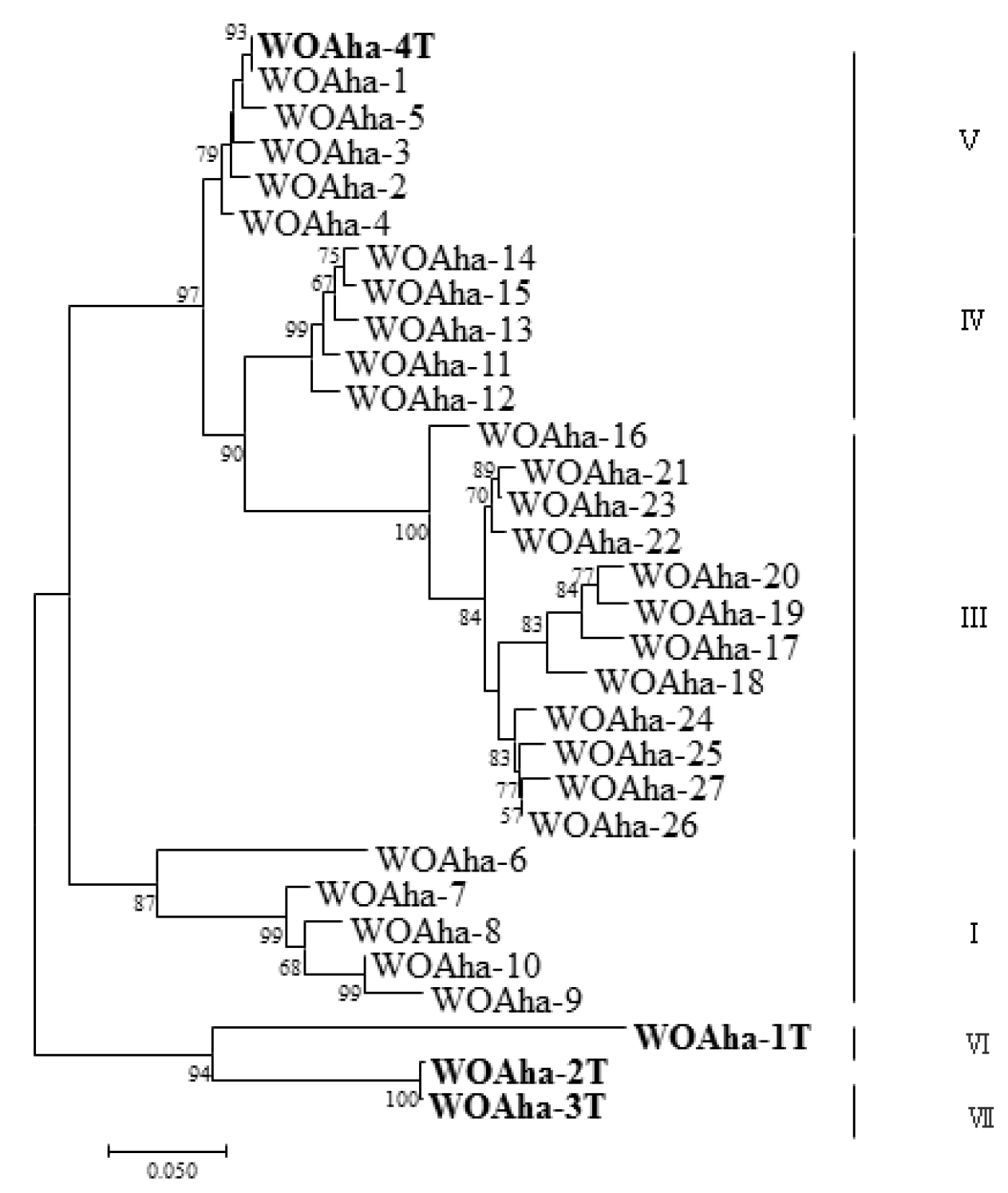
In order to obtain the full-length sequence of the phage WO orf7 gene, we first used inverse PCR. Unfortunately, we were unable to obtain the target gene sequence despite trying various combinations of restriction enzymes. Owing to the failure of the inverse PCR experiment, we were only able to obtain specific viral amplification primers by comparative analysis of virus-related gene sequences from published Wolbachia genomes (Table S1). We downloaded Wolbachia genomes containing the viral gene fragments and obtained the specific primers WO-TF and WO-TR (Table 1) by comparing the genes in the conserved region. We obtained a total of 32 positive clones that could be divided into four types, from WOAha-1T to WOAha-4T. Surprisingly, phylogeny results showed that A. hakonensis was infected with a large number of other phage WO groups, in addition to the four groups identified using WO-F and WO-R (Figure 1). In fact, only WOAha-4T genotypes had previously been detected, whereas WOAha-1T, WOAha-2T and WOAha-3T are newly discovered genotypes [16]. Phylogeny results based on WO-TF and WO-TR showed that three genotypes were undetectable using the primers WO-F/R. It is possible that these results may be due to amplification bias of the primers we used. However, it is indisputable that universal primers, such as WO-F and WO-R, cannot efficiently amplify all phage WO genotypes. Combined with results of the previously identified phage WO genotypes, the phylogeny results show that A. hakonensis contained most of the known orf7 gene types (I, III, IV, V, VI and VII) (Figure 2). Most of the orf7 genotypic differences associated with A. hakonensis result in changes in protein sequences (Figure S3).
3.2. Design of Highly Effective Primers for virus Amplification
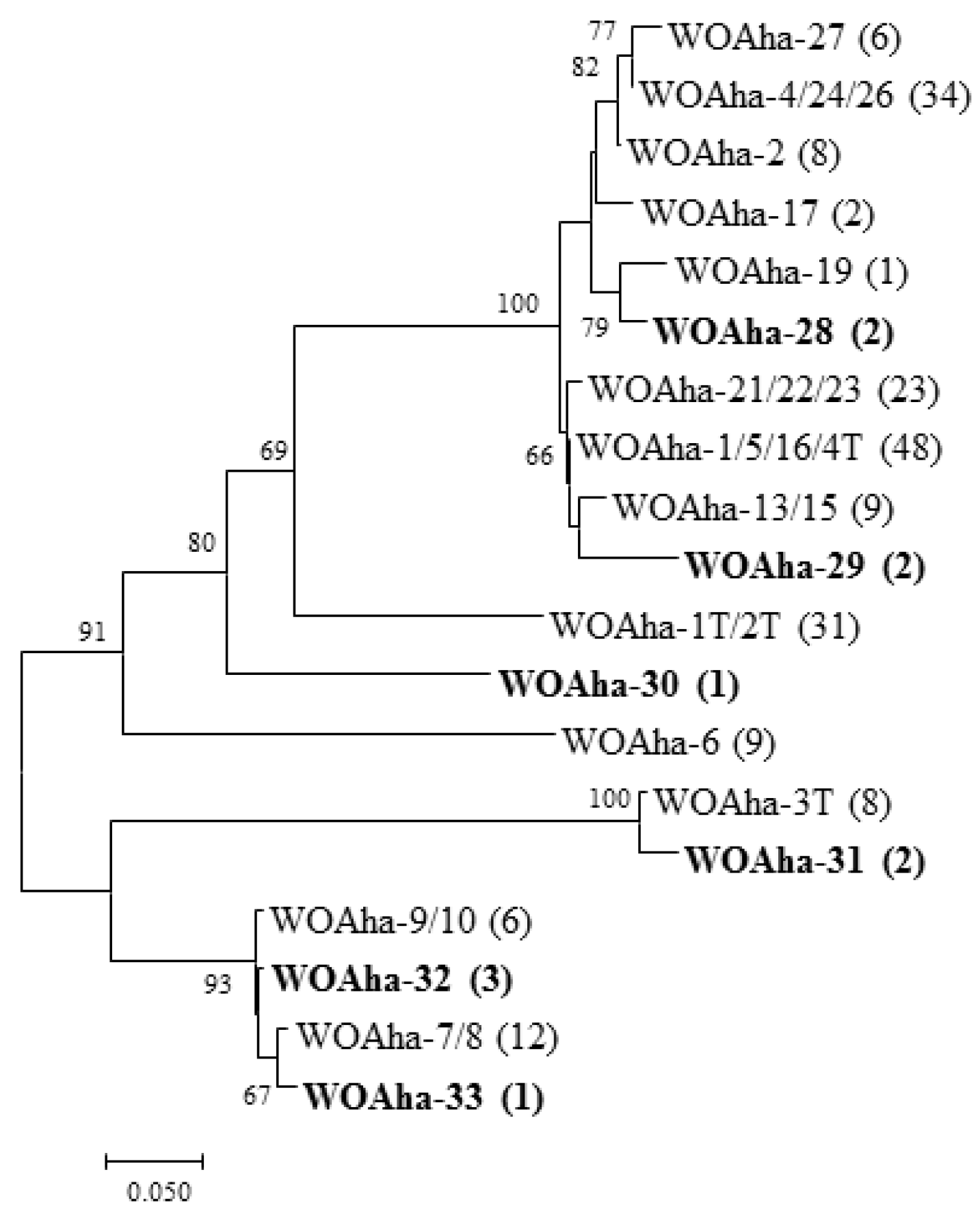
In view of our results, we speculate that existing primers are not specific enough to amplify all phage WO types. We found that primers WO-F and WO-R resulted in a large amount of nonspecific amplification, including around 350 bp, that could interfere with the measurement of infection rates. These results support the urgent need to design new detection primers for phage WO. Based on the existing orf7 gene sequence, we designed specific detection primers (WO-SUF and WO-SUR) (Table 1) to obtain the target band of around 250 bp. The relative positions of all primers used in this article are shown in Figure S4. We obtained a total of 202 positive clones that could be divided into 19 types. In addition to amplifying most of the known sequences, we also detected some new genotypes (Figure 3). Most importantly, all phage WO groups could be efficiently detected. Thus, to the best of our knowledge, A. hakonensis contains the largest number of phage types (up to 36 types) (Figure 1, Figure 2 and Figure 3). We also applied the primers to ten other gall wasps, fifteen crickets, three locusts, two aphids and sixteen butterflies and were able to obtain specific amplification bands for all (Figure S2).
3.3. Base Deletions in orf7
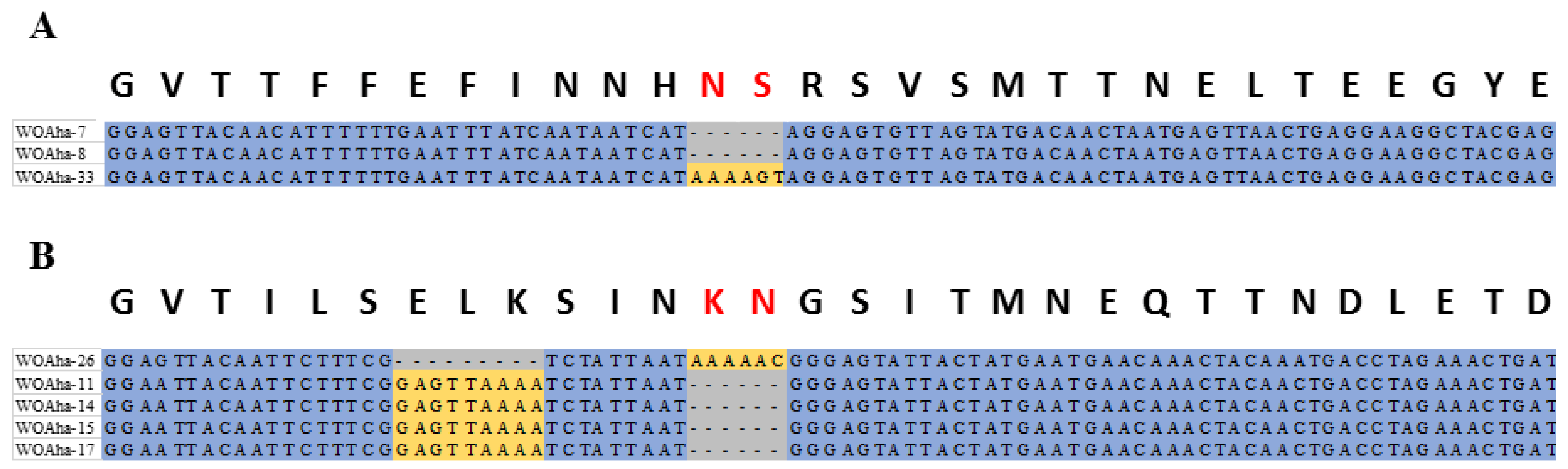
Diverse phage WO types in A. hakonensis provided a perfect sample for obtaining evidence of genetic evolution. This article is novel in providing actual molecular evidence supporting that base deletions are also an important cause of phage WO diversity associated with A. hakonensis, in addition to gene mutations and genetic recombination, which have been reported previously [16]. When the gene sequences were aligned in the way that amino acids are encoded, we saw that all the base deletions caused amino acid deletions (Figure 4). The WOAha-33 deletion “TAAAAG” resulted in WOAha-7 and WOAha-8 and two base deletions were found among WOAha-26, WOAha-11, WOAha-14, WOAha-15 and WOAha-17.
3.4. A High Proportion of Phage WO Types Are Actively Transcribed in A. hakonensis
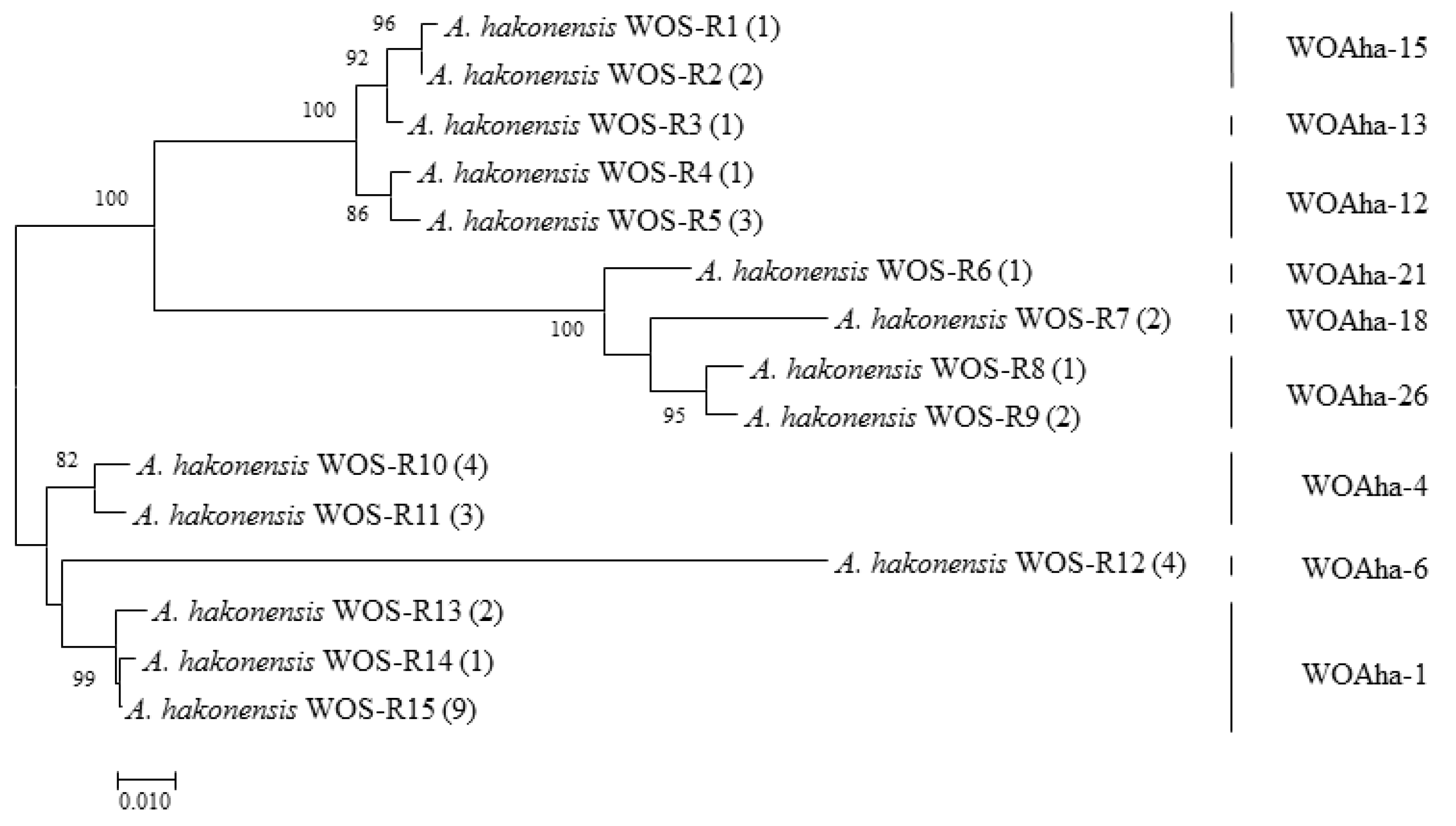
We verified the transcriptional activity of phage WO in A. hakonensis using reverse transcription PCR (RT-PCR). We obtained a total of 37 positive clones, which could be summarized as nine types belonging to WOAha-1/4/6/12/13/15/18/21 and WOAha-26 (Figure 5). These results strongly suggest that most phage WO in A. hakonensis are transcribed normally and are able to perform normal biological functions.
4. Discussion
Although bacteriophages are typically thought to be rare or absent in obligate intracellular bacteria [35,36], phage WO is widely distributed in a variety of Wolbachia-infected insect groups [13,14,27,37,38,39,40]. Moreover, multiple phage infections (Wolbachia strains containing more than one phage type) can be found in several Wolbachia strains [14,40].
Our phylogeny results reveal a high level of phage WO diversity in A. hakonensis and show that phage WO can be divided into four groups [16]. Phylogeny results based on WO-TF and WO-TR show that most of the genotypes were undetectable using the original universal primer WO-F /R, indicating that existing primers are not specific enough to amplify all phage WO types. Therefore, there is an urgent need to design a new detection primer for phage WO. In A. hakonensis, the primers WO-SUF and WO-SUR efficiently detected all known phage WO groups, demonstrating the high efficiency of the primers. These results confirm that A. hakonensis contains the largest number of phage types (up to 36 types), which can be divided into six groups (I, III, IV, V, VI and VII). However, phage-infected Wolbachia strains are thought to harbor low numbers of phage types (with 85% harboring only one or two phage WO types) [14,15]. One explanation for the large number of phage types in A. hakonensis is genetic mutations. The results of Zhu et al. (2021) [16] strongly suggest that intragenic recombination was an important evolutionary dynamic promoting a high level of phage WO diversity in gall wasps. Moreover, the present study is novel in providing practical molecular evidence supporting the fact that base deletions are also an important cause of phage WO diversity associated with A. hakonensis. We also applied the primers to the other 46 species of insects, belonging to Hymenoptera, Orthoptera, Hemiptera and Lepidoptera, and all of them were able to obtain specific amplification bands. The results fully demonstrated the specificity and effectiveness of the newly designed primers. Phage WO phylogenetic relationships inferred from only a portion of the orf7 gene sequence may not be highly reliable. This is demonstrated by our comparison of WOAha-1T (1089 bp) with WOAha-2T (1068 bp). The first 1016 bases of the sequences of these two genotypes are almost identical, but the remainder of the sequence is completely different (Figure S1). Given the frequency of intragenic recombination, more genetic information is needed to accurately reconstruct phylogenetic relationships of phage WO.
5. Conclusions
Given the results of our experiments, the distribution and types of phage WO in host arthropods may be greater than previous estimates, as the primers used to screen for the presence of phage WO are not sufficiently degenerate to detect all orf7 variants. Our newly designed specific primers (WO-SUF and WO-SUR) may be more suitable for detecting the presence of phage WO in arthropods. Most phage types in A. hakonensis are transcribed normally, which reveals that they are able to perform normal biological functions. Therefore, A. hakonensis may be an ideal model system for studying interactions among eukaryotes, bacteria and bacteriophages.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/insects12080713/s1, Figure S1: Gene sequence alignment between WOAha-1T and WOAha-2T, Figure S2: PCR products of the orf7 gene fragments using WO-SUF and WO-SUR, Figure S3: Maximum likelihood tree for phage WO types of A. hakonensis based on orf7 protein sequence, Figure S4: The relative positions of all primers used in this article. Table S1: Genomes used for primer design in this study.
Author Contributions
Conceptualization, C.-Y.S. and D.-H.Z.; methodology, C.-Y.S. and D.-H.Z.; validation, C.-Y.S., D.-H.Z. and X.-H.Y.; formal analysis, C.-Y.S., D.-H.Z. and X.-H.Y.; investigation, X.-H.Y.; resources, C.-Y.S.; data curation, C.-Y.S.; writing—original draft preparation, C.-Y.S. and D.-H.Z.; writing—review and editing, C.-Y.S. and D.-H.Z.; visualization, X.-H.Y.; supervision, D.-H.Z.; project administration, D.-H.Z.; funding acquisition, D.-H.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the National Key Research and Development Program of China (no. 2016YFE0128200).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
All the sequencing results in the current study were deposited in GenBank under the following accession numbers: MZ687122-MZ687127; MZ687129-MZ687140; MZ687810-MZ687824.
Acknowledgments
We thank Yoshihisa Abe of Kyushu University for the identification of Andricus hakonensis. The host plant Quercus fabri Hance was kindly identified by Xun-Lin Yu 433 of Central South University of Forestry and Technology.
Conflicts of Interest
The authors have no conflicts of interest to declare.
References
- Stouthamer, R.; Breeuwer, J.A.; Hurst, G.D. Wolbachia pipientis: Microbial manipulator of arthropod reproduction. Annu. Rev. Microbiol. 1999, 53, 71–102. [Google Scholar] [CrossRef]
- Hilgenboecker, K.; Hammerstein, P.; Schlattmann, P.; Telschow, A.; Werren, J.H. How many species are infected with Wolbachia? A statistical analysis of current data. FEMS Microbiol. Lett. 2008, 281, 215–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenton, A.; Johnson, K.N.; Brownlie, J.C.; Hurst, G.D.D. Solving the Wolbachia paradox: Modeling the tripartite interaction between host, Wolbachia, and a natural enemy. Am. Nat. 2011, 178, 333–342. [Google Scholar] [CrossRef] [Green Version]
- Landmann, F. The Wolbachia endosymbionts. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef]
- O’Neill, S.L.; Karr, T.L. Bidirectional incompatibility between conspecific populations of Drosophila simulans. Nature 1990, 348, 178–180. [Google Scholar] [CrossRef]
- Stouthamer, R.; Luck, R.F.; Hamilton, W.D. Antibiotics cause parthenogenetic Trichogramma (Hymenoptera/Trichogrammatidae) to revert to sex. Proc. Natl. Acad. Sci. USA 1990, 87, 2424–2427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiggins, F.M.; Hurst, G.D.; Schulenburg, J.H.; Majerus, M.E. Two male-killing Wolbachia strains coexist within a population of the butterfly Acraea encedon. Heredity 2001, 86, 161–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouchon, D.; Rigaud, T.; Juchault, P. Evidence for widespread Wolbachia infection in isopod crustaceans: Molecular identification and host feminization. Proc. R. Soc. Lond. B Biol. Sci. 1998, 265, 1081–1090. [Google Scholar] [CrossRef] [Green Version]
- Plett, J.M.; Kemppainen, M.; Kale, S.D.; Kohler, A.; Legué, V.; Brun, A.; Tyler, B.M.; Pardo, A.G.; Martin, F. A secreted effector protein of Laccaria bicolor is required for symbiosis development. Curr. Biol. 2011, 21, 1197–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saridaki, A.; Sapountzis, P.; Harris, H.L.; Batista, P.D.; Biliske, J.A.; Pavlikaki, H.; Oehler, S.; Savakis, C.; Braig, H.R.; Bourtzis, K. Wolbachia Prophage DNA Adenine Methyltransferase Genes in Different Drosophila-Wolbachia Associations. PLoS ONE 2011, 6, e19708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LePage, D.P.; Metcalf, J.A.; Bordenstein, S.R.; On, J.; Perlmutter, J.I.; Shropshire, J.D.; Layton, E.M.; Funkhouser-Jones, L.J.; Beckmann, J.F.; Bordenstein, S.R. Prophage WO genes recapitulate and enhance Wolbachia induced cytoplasmic incompatibility. Nature 2017, 543, 243–247. [Google Scholar] [CrossRef] [Green Version]
- Shropshire, J.D.; On, J.; Layton, E.M.; Zhou, H.; Bordenstein, S.R. One prophage WO gene rescues cytoplasmic incompatibility in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2018, 115, 4987–4991. [Google Scholar] [CrossRef] [Green Version]
- Bordenstein, S.R.; Wernegreen, J.J. Bacteriophage flux in endosymbionts (Wolbachia): Infection frequency, lateral transfer, and recombination rates. Mol. Biol. Evol. 2004, 21, 1981–1991. [Google Scholar] [CrossRef] [Green Version]
- Gavotte, L.; Henri, H.; Stouthamer, R.; Charif, D.; Charlat, S.; Boulétreau, M.; Vavre, F. A survey of the bacteriophage WO in the endosymbiotic bacteria Wolbachia. Mol. Biol. Evol. 2007, 24, 427–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, K.; Furukawa, S.; Nikoh, N.; Sasaki, T.; Fukatsu, T. Complete WO phage sequences reveal their dynamic evolutionary trajectories and putative functional elements required for integrationin to the Wolbachia genome. Appl. Environ. Microbiol. 2009, 75, 5676–5686. [Google Scholar] [CrossRef] [Green Version]
- Zhu, D.H.; Su, C.Y.; Yang, X.H.; Abe, Y. A case of intragenic recombination dramatically impacting the phage WO genetic diversity in gall wasps. Front. Microbiol. 2021, 12, 694115. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Nieves-Aldrey, J.L.; Buffington, M.L.; Liu, Z.; Liljeblad, J.; Nylander, J.A.A. Phylogeny, evolution and classification of gall wasps: The plot thickens. PLoS ONE 2015, 10, e0123301. [Google Scholar] [CrossRef] [PubMed]
- Plantard, O.; Rasplus, J.Y.; Mondor, G.; Le Clainche, I.; Solignac, M. Wolbachia induced thelytoky in the rose gallwasp Diplolepis spinosissimae (Giraud) (Hymenoptera: Cynipidae), and its consequences on the genetic structure of its host. Proc. R. Soc. Lond. B Biol. Sci. 1998, 265, 1075–1080. [Google Scholar] [CrossRef]
- Abe, Y.; Miura, K. Doses Wolbachia induce unisexuality in oak gall wasps? (Hymenoptera: Cynipidae). Ann. Entomol. Soc. Am. 2002, 95, 583–586. [Google Scholar] [CrossRef]
- Rokas, A.; Atkinson, R.J.; Nieves-Aldrey, J.L.; West, S.A.; Stone, G.N. The incidence and diversity of Wolbachia in gallwasps (Hymenoptera; Cynipidae) on oak. Mol. Ecol. 2002, 11, 1815–1829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, D.H.; He, Y.Y.; Fan, Y.S.; Ma, M.Y.; Peng, D.L. Negative evidence of parthenogenesis induction by Wolbachia in a gallwasp species, Dryocosmus kuriphilus. Entomol. Exp. Appl. 2007, 124, 279–284. [Google Scholar] [CrossRef]
- Yang, X.H.; Zhu, D.H.; Liu, Z.; Zhao, L.; Su, C.Y. High levels of multiple infections, recombination and horizontal transmission of Wolbachia in the Andricus mukaigawae (Hymenoptera; Cynipidae) communities. PLoS ONE 2013, 8, e78970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, H.Q.; Zhao, G.Z.; Su, C.Y.; Zhu, D.H. Wolbachia prevalence patterns: Horizontal transmission, recombination, and multiple infections in chestnut gall wasp–parasitoid communities. Entomol. Exp. Appl. 2020, 168, 752–765. [Google Scholar] [CrossRef]
- Zhao, G.Z.; Zhu, T.R.; Zeng, Y.; Zhu, D.H. Wolbachia infection in six species of gall wasps and their parasitoids. J. Asia-Pac. Entomol. 2021, 24, 21–25. [Google Scholar] [CrossRef]
- Bordenstein, S.R.; Marshall, M.L.; Fry, A.J.; Kim, U.; Wernegreen, J.J. The tripartite associations between bacteriophage, Wolbachia, and arthropods. PLoS Pathog. 2006, 2, e43. [Google Scholar]
- Metcalf, J.A.; Bordenstein, S.R. The complexity of virus systems: The case of endosymbionts. Curr. Opin. Microbiol. 2012, 15, 546–552. [Google Scholar] [CrossRef] [Green Version]
- Chafee, M.E.; Funk, D.J.; Harrison, R.G.; Bordenstein, S.R. Lateral phage transfer in obligate intracellular bacteria (Wolbachia): Verification from natural populations. Mol. Biol. Evol. 2010, 27, 501–505. [Google Scholar] [CrossRef]
- Masui, S.; Kamoda, S.; Sasaki, T.; Ishikawa, H. Distribution and evolution of bacteriophage WO in Wolbachia, the endosymbiont causing sexual alterations in arthropods. J. Mol. Evol. 2000, 51, 491–497. [Google Scholar] [CrossRef]
- Klasson, L.; Westberg, J.; Sapountzis, P.; Näslund, K.; Lutnaes, Y.; Darby, A.C.; Veneti, Z.; Chen, L.; Braig, H.R.; Garrett, R.; et al. The mosaic genome structure of the Wolbachia wRi strain infecting Drosophila simulans. Proc. Natl. Acad. Sci. USA 2009, 106, 5725–5730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Partensky, F.; Garczarek, L. Microbiology:arms race in a drop of sea water. Nature 2011, 474, 582–583. [Google Scholar] [CrossRef]
- Zhou, W.; Rousset, F.; Neill, S.O. Phylogeny and PCR-based classification of Wolbachia strains using wsp gene sequences. Proc. R. Soc. Lond. B Biol. Sci. 1998, 265, 509. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES science gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Dyer, K.A.; Burke, C.; Jaenike, J. Wolbachia-mediated persistence of mtDNA from a potentially extinct species. Mol. Ecol. 2011, 20, 2805–2817. [Google Scholar] [CrossRef] [PubMed]
- Moran, N.A.; Plague, G.R. Genomic changes following host restriction in bacteria. Curr. Opin. Genet. Dev. 2004, 14, 627–633. [Google Scholar] [CrossRef]
- Bordenstein, S.R.; Reznikoff, W.S. Mobile DNA in obligate intracellular bacteria. Nat. Rev. Microbiol. 2005, 3, 688–699. [Google Scholar] [CrossRef]
- Kent, B.N.; Bordenstein, S.R. Phage WO of Wolbachia: Lambda of the endosymbiont world. Trends Microbiol. 2010, 18, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.H.; Sun, B.F.; Xiong, T.L.; Wang, Y.K.; Murfin, K.E.; Xiao, J.H.; Huang, D.W. Bacteriophage WO can mediate horizontal gene transfer in endosymbiotic Wolbachia genomes. Front. Microbiol. 2016, 7, 1867. [Google Scholar] [CrossRef]
- Kaushik, S.; Sharma, K.K.; Ramani, R.; Lakhanpaul, S. Detection of Wolbachia Phage (WO) in Indian Lac Insect [Kerria lacca (Kerr.)] and Its Implications. Indian. J. Microbiol. 2019, 59, 237–240. [Google Scholar] [CrossRef]
- Chauvatcharin, N.; Ahantarig, A.; Baimai, V.; Kittayapong, P. Bacteriophage WO-B and Wolbachia in natural mosquito hosts: Infection incidence, transmission mode and relative density. Mol. Ecol. 2006, 15, 2451–2461. [Google Scholar] [CrossRef]
Figure 1.
Maximum likelihood tree for phage WO types of A. hakonensis based on full length of orf7 sequences. The numbers in parentheses indicate the number of sequences obtained. WOS-xT refers to the serial number. Phage WO types are shown on the right. Numbers above and below branches are bootstrap values computed from 1000 replications.
Figure 1.
Maximum likelihood tree for phage WO types of A. hakonensis based on full length of orf7 sequences. The numbers in parentheses indicate the number of sequences obtained. WOS-xT refers to the serial number. Phage WO types are shown on the right. Numbers above and below branches are bootstrap values computed from 1000 replications.

Figure 2.
Maximum likelihood tree for phage WO types of A. hakonensis based on orf7 sequences. WOAha-X refers to the phage WO types, “T” indicate the full length sequence of orf7. Numbers above and below branches are bootstrap values computed from 1000 replications.
Figure 2.
Maximum likelihood tree for phage WO types of A. hakonensis based on orf7 sequences. WOAha-X refers to the phage WO types, “T” indicate the full length sequence of orf7. Numbers above and below branches are bootstrap values computed from 1000 replications.

Figure 3.
Maximum likelihood tree for phage WO types of A. hakonensis based on orf7 sequences obtain with new detection primers. The numbers in parentheses indicate the number of sequences obtained.
Figure 3.
Maximum likelihood tree for phage WO types of A. hakonensis based on orf7 sequences obtain with new detection primers. The numbers in parentheses indicate the number of sequences obtained.

Figure 4.
Bases deletions of the orf7 gene (A) WOAha-33 deletion “TAAAAG” resulting in WOAha-7 and WOAha-8, and (B) Bases deletions among WOAha-26, WOAha-11, WOAha-14, WOAha-15, and WOAha-17.
Figure 4.
Bases deletions of the orf7 gene (A) WOAha-33 deletion “TAAAAG” resulting in WOAha-7 and WOAha-8, and (B) Bases deletions among WOAha-26, WOAha-11, WOAha-14, WOAha-15, and WOAha-17.

Figure 5.
Maximum likelihood tree actively transcribed phage WO types of A. hakonensis based on orf7 sequences using WO-F; WO-R primers. WOS-Rx refers to the serial number. The numbers in parentheses indicate the number of sequences obtained. Phage WO types are shown on the right. Numbers above and below branches are bootstrap values computed from 1000 replications.
Figure 5.
Maximum likelihood tree actively transcribed phage WO types of A. hakonensis based on orf7 sequences using WO-F; WO-R primers. WOS-Rx refers to the serial number. The numbers in parentheses indicate the number of sequences obtained. Phage WO types are shown on the right. Numbers above and below branches are bootstrap values computed from 1000 replications.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primers used in this study.
| Gene | Primer | Primer Sequence | Annealing Temperature | References | |
|---|---|---|---|---|---|
| Phage WO | orf7 | WO-F WO-R | 5′-CCC ACA TGA GCC AAT GAC GTC TG-3′ 5′-CGT TCG CTC TGC AAG TAA CTC CAT TAA AAC-3′ | 57 °C | Masui et al., 2000 |
| WO-TF WO-TR | 5′-CTTGGCTACYGTTATYTGRTCTCTTG-3′ 5′-TCYGASGTTACTRCHAATCAAGAGG-3′ | 59 °C | This study | ||
| WO-SUF: WO-SUR: | 5′-ARGCAGGKCTWTATTTTGG-3′ 5′-ATTCTTCTATYTTYYCTGGCA-3′ | 50 °C | This study | ||
| Wolbachia | wsp | 81F 691R | 5′-TGG TCC AAT AAG TGA TGA AGA AAC-3′ 5′-AAA AAT TAA ACG CTA CTC CA-3′ | 52 °C | Zhou et al.,1998 |
| Insect | ITS2 | ITS2F ITS2R | 5′-TGT GAA CTG CAG GAC ACA TG-3′ 5′-AAT GCT TAA ATT TAG GGG GTA-3′ | 54 °C | Partensky and Garczarek, 2011 |
| cox1 | LCO-1490 HCO-2198 | 5′-GGTCAACAAATCATAAAGATATTGG-3′ 5′-TAAACTTCAGGGTGACCAAAAATCA-3′ | 55 °C | Dyer et al., 2011 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Su, C.-Y.; Zhu, D.-H.; Yang, X.-H. Design and Testing of Effective Primers for Amplification of the orf7 Gene of Phage WO Associated with Andricus hakonensis. Insects 2021, 12, 713. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12080713
AMA Style
Su C-Y, Zhu D-H, Yang X-H. Design and Testing of Effective Primers for Amplification of the orf7 Gene of Phage WO Associated with Andricus hakonensis. Insects. 2021; 12(8):713. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12080713
Chicago/Turabian StyleSu, Cheng-Yuan, Dao-Hong Zhu, and Xiao-Hui Yang. 2021. "Design and Testing of Effective Primers for Amplification of the orf7 Gene of Phage WO Associated with Andricus hakonensis" Insects 12, no. 8: 713. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12080713
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.