
Three-Dimensional Culture of Rhipicephalus (Boophilus) microplus BmVIII-SCC Cells on Multiple Synthetic Scaffold Systems and in Rotating Bioreactors

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tick Cell Line
2.2. Cell Culture Medium
2.3. 2-D Cell Culture
2.4. 2-D Cell Morphometrics
2.5. 2-D Cell Histological Evaluation
2.6. 3-D Tick Cell-Tissue Culture
2.6.1. Nanofiber Disks
2.6.2. Hydrogel
2.6.3. RCCS Bioreactors
Bead Preparation for Bioreactors
Bioreactor Setup
Sampling and Microscopic Observation of Cell Aggregates
2.7. Immunohistochemistry (IHC)
2.8. 3-D Light Microscopy
2.9. Electron Microscopy
2.10. Immunohistochemistry of E-SEM and TEM Prepared Tick Material
3. Results
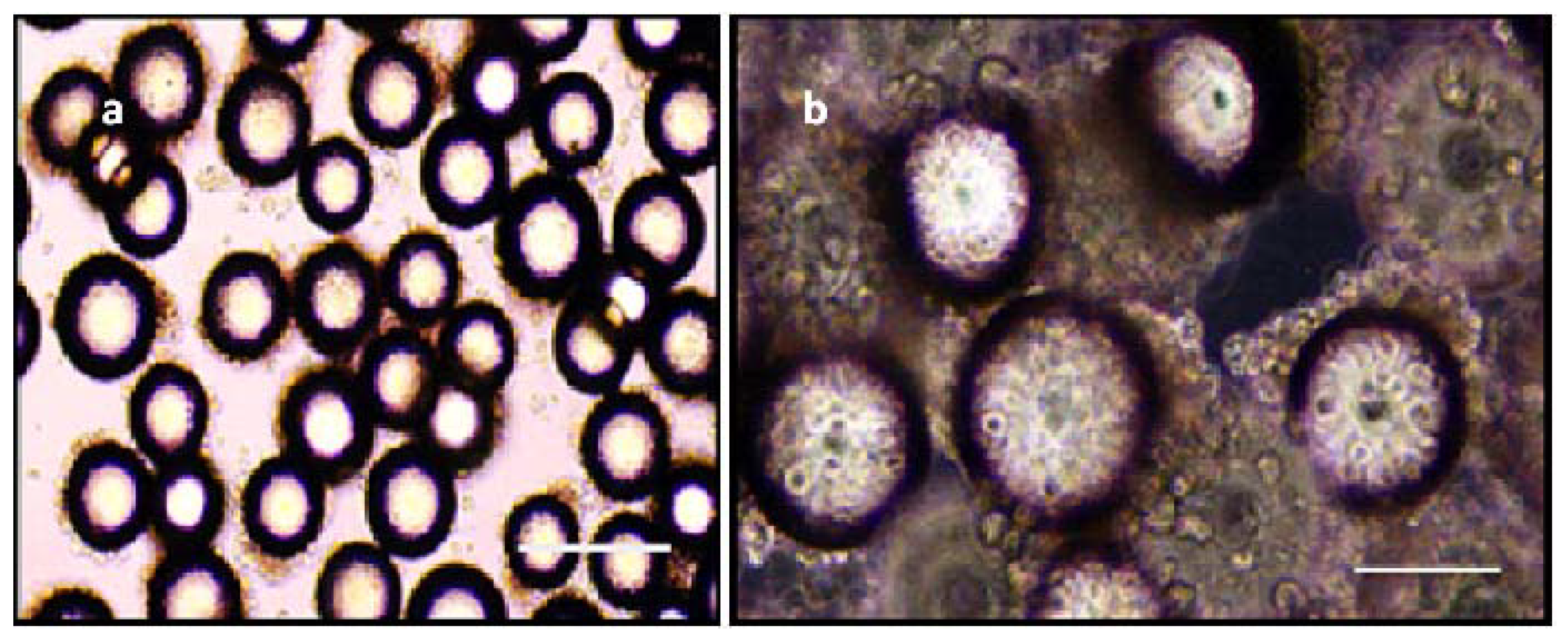
3.1. Cell Source for 3-D Culture Experiments
3.2. Hydrogel and 3-D Culture
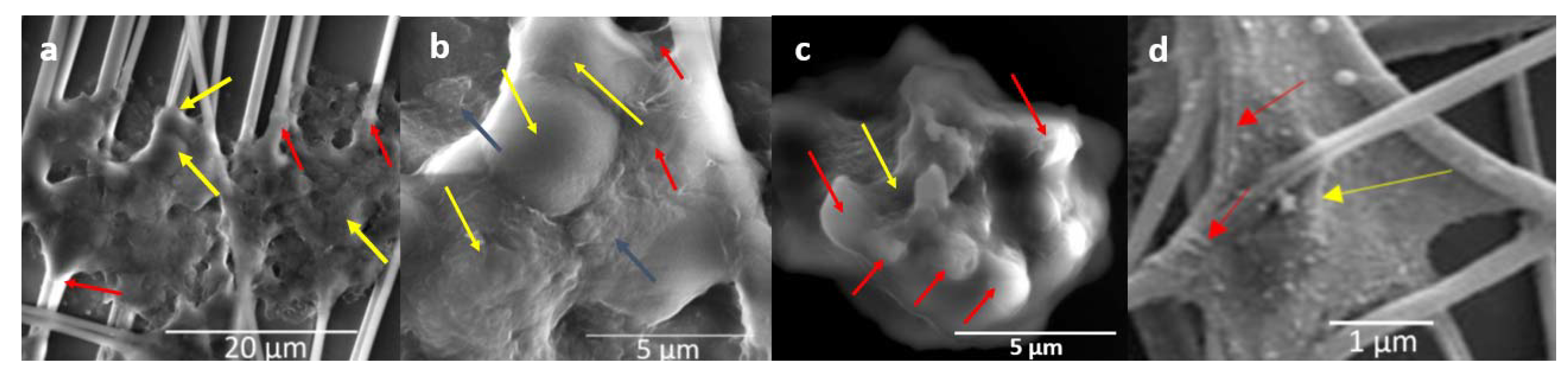
3.3. Nanofiber Disk 3-D Culture
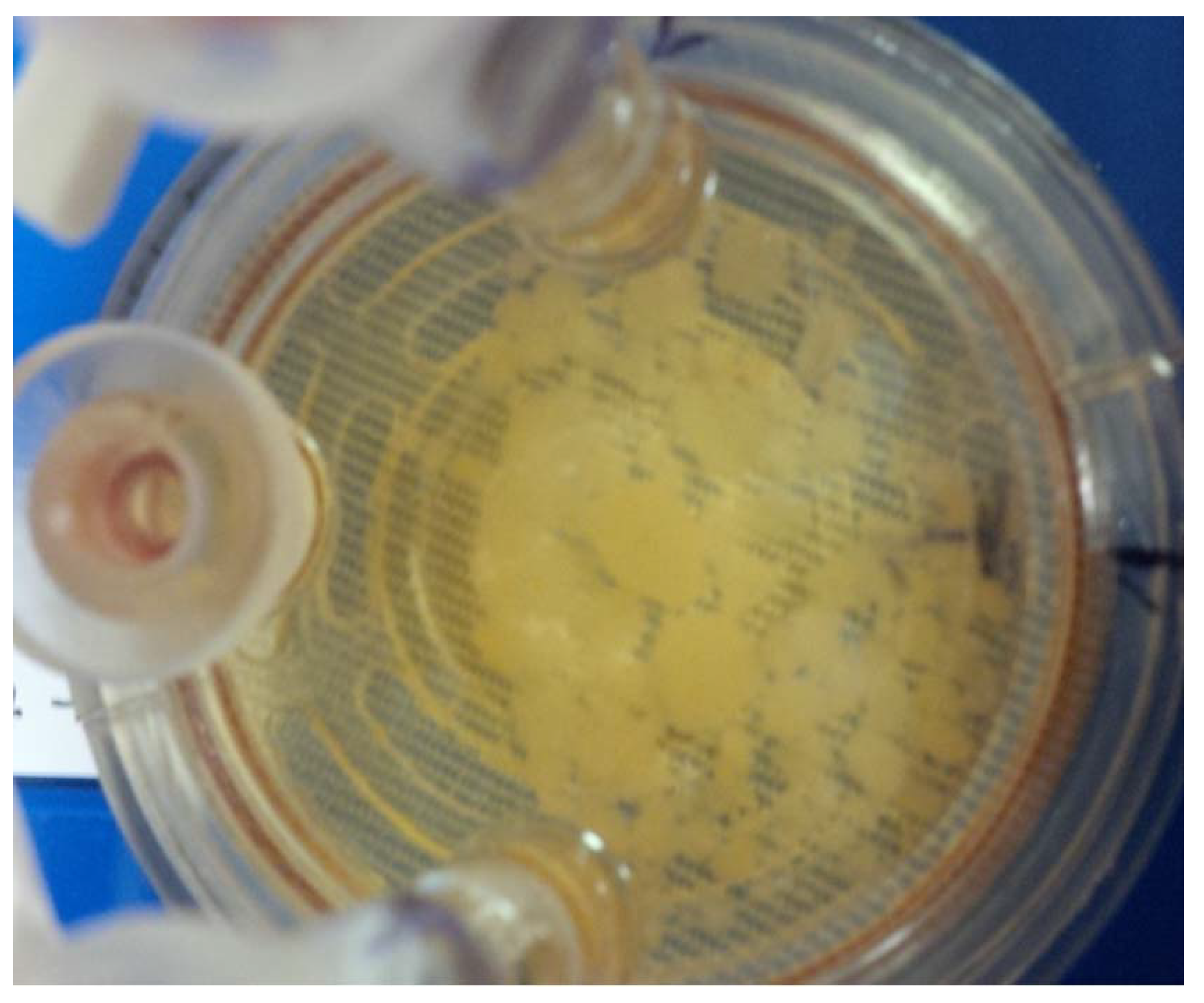
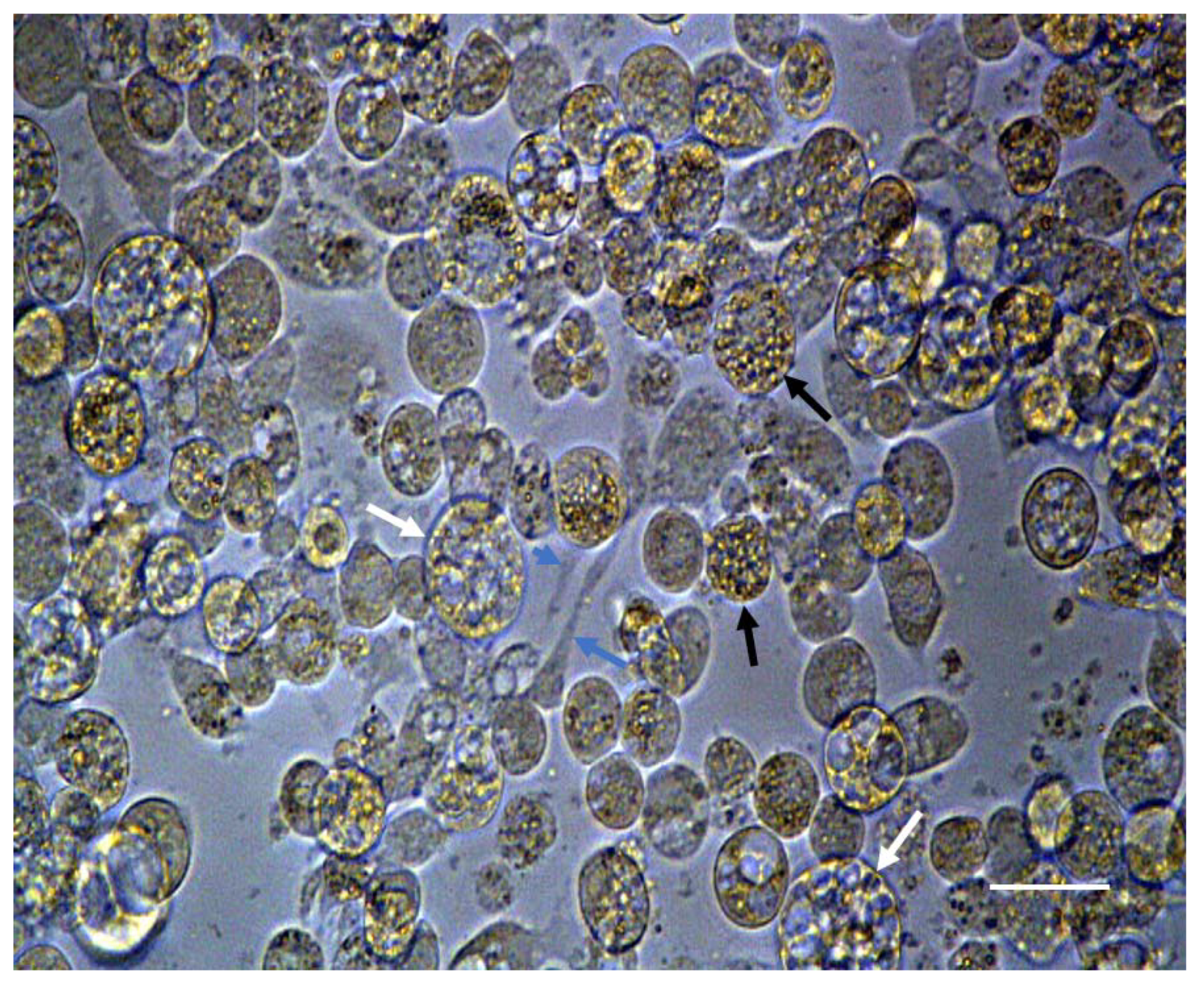
3.4. Beads in RCCS
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boulanger, N.; Boyer, P.; Talagrand-Reboul, E.; Hansmann, Y. Review—Ticks and tick-borne diseases [Tiques et maladies vectorielles á tiques]. Méd. Mal. Infect. 2019, 49, 87–97. [Google Scholar] [CrossRef]
- Eisen, L. Control of ixodid ticks and prevention of tick-borne diseases in the United States: The prospect of a new Lyme disease vaccine and the continuing problem with tick exposure on residential properties. Ticks Tick borne Dis. 2021, 12, 101648. [Google Scholar] [CrossRef]
- Grisi, L.; Leite, R.C.; de Souza Martins, J.R.; Medeiros de Barros, A.T.; Anderotti, R.; Duarte Cancado, P.H.; Pérez de León, A.A.; Barros Pereira, J.; Silva Villela, H. Reassessment of the potential economic impact of cattle parasites in Brasil. Rev. Bras. Parasitol. Vet. 2014, 23, 150–156. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Vivas, R.I.; Grisi, L.; Pérez de León, A.A.; Silva Villela, H.; Torres-Acosta, J.F.J.; Fragoso Sanchez, H.; Romero Salas, D.; Rosario Cruz, R.; Saldierna, F.; Garcia Carrasco, D. Potential economic impact assessment for cattle parasites in Mexico. Rev. Mex. Cienc. Pec. 2017, 8, 61–74. [Google Scholar] [CrossRef]
- Pérez de León, A.A.; Mitchell, R.D., III; Miller, R.J.; Lohmeyer, K.H. Advances in integrated tick management research for area-wide mitigation of tick-borne disease burden. In Area-Wide Integrated Pest Management: Development and Field Application; Pereira, R., Vreysen, M.J.B., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 251–274. [Google Scholar]
- Higa, L.O.S.; Piña, F.T.B.; Rodrigues, V.S.; Garcia, M.V.; Salas, D.R.; Miller, R.J.; Pérez de León, A.; Barros, J.C.; Andreotti, R. Evidence of acaricide resistance in different life stages of Amblyomma mixtum and Rhipicephalus microplus (Acari: Ixodidae) collected from the same farm in the state of Veracruz, Mexico. Prev. Vet. Med. 2020, 174, 1–5. [Google Scholar] [CrossRef]
- Valsoni, L.M.; de Freitas, M.G.; Echeverria, J.T.; Borges, D.G.L.; Tutija, J.; de Almeida Borges, F. Resistance to all chemical groups of acaricides in a single isolate of Rhipicephalus microplus in Mato Grosso do Sul, Brazil. Int. J. Acarol. 2020, 46, 276–280. [Google Scholar] [CrossRef]
- Rodriguez-Vivas, R.I.; Jonsson, N.N.; Bhushan, C. Strategies for the control of Rhipicephalus microplus ticks in a world of conventional acaricide and macrocyclic lactone resistance. Parasitol. Res. 2018, 117, 3–29. [Google Scholar] [CrossRef] [Green Version]
- Vilela, V.L.R.; Feitosa, T.F.; Bezerra, R.A.; Klafke, G.M.; Riet-Correa, F. Multiple acaricide-resistant Rhipicephalus microplus in the semi-arid region of Paraíba State, Brazil. Ticks Tick Borne Dis. 2020, 11, 101413. [Google Scholar] [CrossRef]
- Bell-Sakyi, L.; Darby, A.; Baylis, M.; Makepeace, B.L. The tick cell biobank: A global resource for in vitro research on ticks, other arthropods and the pathogens they transmit. Ticks Tick borne Dis. 2018, 9, 1364–1371. [Google Scholar] [CrossRef]
- Al-Rofaai, A.; Bell-Sakyi, L. Tick cell lines in research on tick control. Front. Physiol. 2020, 11, 152. [Google Scholar] [CrossRef] [Green Version]
- Bell-Sakyi, L.; Zweygarth, E.; Blouin, E.F.; Gould, E.A.; Jongejan, F. Tick cell lines: Tools for tick and tick-borne disease research. Trends Parasitol. 2007, 23, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Bell-Sakyi, L.; Palomar, A.M.; Bradford, E.L.; Shkap, V. Propagation of the Israeli vaccine strain of Anaplasma centrale in tick cell lines. Vet. Microbiol. 2015, 179, 270–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moniuszko, A.; Rückert, C.; Alberdi, M.P.; Barry, G.; Stevenson, B.; Fazakerley, J.K.; Kohl, A.; Bell-Sakyi, L. Coinfection of tick cell lines has variable effects on replication of intracellular bacterial and viral pathogens. Ticks Tick Borne Dis. 2014, 5, 415–422. [Google Scholar] [CrossRef]
- Bell-Sakyi, L.; Attoui, H. Virus discovery using tick cell lines. Evol. Bioinform. 2016, 12 (Suppl. 2), 31–34. [Google Scholar]
- Contreras, M.; Villar, M.; Alberdi, P.; de la Fuente, J. Vaccinomics approach to tick vaccine development. Methods Mol. Biol. 2016, 1404, 275–286. [Google Scholar] [PubMed]
- Mangia, C.; Vismarra, A.; Kramer, L.; Bell-Sakyi, L.; Porretta, D.; Otranto, D.; Epis, S.; Grandi, G. Evaluation of the in vitro expression of ATP binding-cassette (ABC) proteins in an Ixodes ricinus cell line exposed to ivermectin. Parasit. Vectors 2016, 9, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pohl, P.C.; Carvalho, D.D.; Daffre, S.; da Silva Vaz, I., Jr.; Masuda, A. In vitro establishment of ivermectin-resistant Rhipicephalus microplus cell line and the contribution of ABC transporters on the resistance mechanism. Vet. Parasitol. 2014, 204, 316–322. [Google Scholar] [CrossRef] [Green Version]
- Guerrero, F.D.; Miller, R.J.; Pérez de Léon, A.A. Cattle tick vaccines: Many candidate antigens, but will a commercially viable product emerge? Int. J. Parasitol. 2012, 42, 421–427. [Google Scholar] [CrossRef]
- Pudney, M.; Varma, M.G.R.; Leake, C.J. Culture of embryonic cells from the tick Boophilus microplus (Ixodidae). J. Med. Entomol. 1973, 10, 493–496. [Google Scholar] [CrossRef]
- Holman, P.J.; Ronald, N.C. A new tick cell line derived from Boophilus microplus. Res. Vet. Sci. 1980, 29, 383–387. [Google Scholar]
- Holman, P.J. Partial characterization of a unique female diploid cell strain from the tick Boophilus microplus (Acari: Ixodidae). J. Med. Entomol. 1981, 18, 84–88. [Google Scholar] [CrossRef]
- Kurtti, T.J.; Munderloh, U.G.; Ahlstrand, G.G.; Johnson, R.C. Borrelia burgdorferi in tick cell culture: Growth and cellular adherence. J. Med. Entomol. 1988, 25, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Bell-Sakyi, L.; Kohl, A.; Bente, D.A.; Fazakerley, J.K. Tick cell lines for study of Crimean-Congo hemorrhagic fever virus and other arboviruses. Vector Borne Zoonotic Dis. 2012, 12, 769–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esteves, E.; Lara, F.A.; Lorenzini, D.M.; Costa, G.H.; Fukuzawa, A.H.; Pressinotti, L.N.; Silva, J.R.; Ferro, J.A.; Kurtti, T.J.; Munderloh, U.G.; et al. Cellular and molecular characterization of an embryonic cell line (BME26) from the tick Rhipicephalus (Boophilus) microplus. Insect Biochem. Mol. Biol. 2008, 38, 568–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuckow, A.P.; Temeyer, K.B. Discovery, adaptation and transcriptional activity of two tick promoters: Construction of a dual luciferase reporter system for optimization of RNA interference in Rhipicephalus (Boophilus) microplus cell lines. Insect Mol. Biol. 2015, 24, 454–466. [Google Scholar] [CrossRef] [PubMed]
- Cossio-Bayugar, R.; Barhoumi, R.; Burghardt, R.C.; Wagner, G.G.; Holman, P. Basal cellular alterations of esterase, glutathione, glutathione S-transferase, intracellular calcium, and membrane potentials in coumaphos-resistant Boophilus microplus (Acari: Ixodidae) cell lines. Pesticide Biochem. Physiol. 2002, 72, 1–9. [Google Scholar] [CrossRef]
- Cossio-Bayugar, R.; Wagner, G.G.; Holman, P. In vitro generation of organophosphate resistant Boophilus microplus (Acari: Ixodidae) cell lines. J. Med. Entomol. 2002b, 39, 278–284. [Google Scholar] [CrossRef]
- Pohl, P.C.; Klafke, G.M.; Carvalho, D.D.; Martins, J.R.; Daffre, S.; da Silva Vaz, I., Jr.; Masuda, A. ABC transporter efflux pumps: A defense mechanism against ivermectin in Rhipicephalus (Boophilus) microplus. Int. J. Parasitol. 2011, 41, 1323–1333. [Google Scholar] [CrossRef]
- LeGall, V.L.; Klafke, G.M.; Torres, T.T. Detoxification mechanisms involved in ivermectin resistance in the cattle tick, Rhipicephalus (Boophilus) microplus. Sci. Rep. 2018, 8, 12401. [Google Scholar] [CrossRef] [Green Version]
- Koh-Tan, H.H.; Strachan, E.; Cooper, K.; Bell-Sakyi, L.; Jonsson, N.N. Identification of a novel β-adrenergic octopamine receptor-like gene (βAOR-like) and increased ATP-binding cassette B10 (ABCB10) expression in a Rhipicephalus microplus cell line derived from acaricide-resistant ticks. Parasit. Vectors 2016, 9, 425. [Google Scholar] [CrossRef] [Green Version]
- Droleskey, R.E.; Holman, P.J.; Craig, T.M.; Wagner, G.G.; Mollenhauer, H.H. Ultrastructure of Babesia bovis sexual stages as observed in Boophilus microplus cell cultures. Res. Vet. J. 1983, 34, 249–251. [Google Scholar] [CrossRef]
- Esteves, E.; Fogaça, A.C.; Maldonado, R.; Silva, F.D.; Manso, P.P.; Pelajo-Machado, M.; Valle, D.; Daffre, S. Antimicrobial activity in the tick Rhipicephalus (Boophilus) microplus eggs: Cellular localization and temporal expression of microplusin during oogenesis and embryogenesis. Dev. Comp. Immunol. 2009, 33, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Alberdi, M.P.; Dalby, M.J.; Rodriguez-Andres, J.; Fazakerley, J.K.; Kohl, A.; Bell-Sakyi, L. Detection and identification of putative bacterial endosymbionts and endogenous viruses in tick cell lines. Ticks Tick Borne Dis. 2012, 3, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, R.C.; Baêta, B.A.; Ferreira, J.S.; Medeiros, R.C.; Maya-Monteiro, C.M.; Lara, L.A.; Bell-Sakyi, L.; Fonseca, A.H. Fluorescent membrane markers elucidate the association of Borrelia burgdorferi with tick cell lines. Braz J. Med. Biol. Res. 2016, 49, e5211. [Google Scholar] [CrossRef]
- Saramago, L.; Franceschi, M.; Logullo, C.; Masuda, A.; da SilvaVaz, I.; Farias, S.E.; Moraes, J. Inhibition of enzyme activity of Rhipicephalus (Boophilus) microplus triosephosphate isomerase and BME26 cell growth by monoclonal antibodies. Int. J. Mol. Sci. 2012, 13, 13118–13133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Abreu, L.A.; Calixto, C.; Waltero, C.F.; Della Noce, B.P.; Githaka, N.W.; Seixas, A.; Parizi, L.F.; Konnai, S.; da Silva Junior Vaz, I.; Ohashi, K.; et al. The conserved role of the AKT/GSK3 axis in cell survival and glycogen metabolism in Rhipicephalus (Boophilus) microplus embryo tick cell line BME26. Biochim. Biophys. Acta 2013, 1830, 2574–2582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, H.; Romeiro, N.C.; Braz, G.R.; de Oliveira, E.A.; Rodrigues, C.; da Fonseca, C.R.N.; Githaka, N.; Isezaki, M.; Konnai, S.; Ohashi, K.; et al. Identification and structural -functional analysis of cyclin-dependent kinases of the cattle tick Rhipicephalus (Boophilus) microplus. PLoS ONE 2013, 8, e76128. [Google Scholar] [CrossRef] [Green Version]
- da Silva, R.M.; Noce, B.D.; Waltero, C.F.; Costa, E.P.; de Abreu, L.A.; Githaka, N.W.; Moraes, J.; Gomes, H.F.; Konnai, S.; Vaz Ida, S., Jr.; et al. Non-classical gluconeogenesis-dependent glucose metabolism in Rhipicephalus microplus embryonic cell line BME26. Int. J. Mol. Sci. 2015, 16, 1821–1839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estrela, A.B.; Seixas, A.; Teixeira, V.d.O.N.; Pinto, A.F.M.; Termignoni, C. Vitellin- and hemoglobin-digesting enzymes in Rhipicephalus (Boophilus) microplus larvae and females. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2010, 157, 326–335. [Google Scholar] [CrossRef]
- Machado-Ferreira, E.; Balsemão-Pires, E.; Dietrich, G.; Hojgaard, A.; Vizzoni, V.F.; Scoles, G.; Bell-Sakyi, L.; Piesman, J.; Zeidner, N.S.; Soares, C.A. Transgene expression in tick cells using Agrobacterium tumefaciens. Exp. Appl. Acarol. 2015, 67, 269–287. [Google Scholar] [CrossRef]
- Lawrie, C.H.; Uzcátegui, N.Y.; Armesto, M.; Bell-Sakyi, L.; Gould, E.A. Susceptibility of mosquito and tick cell lines to infection with various flaviviruses. Med. Vet. Entomol. 2004, 18, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Langhans, S.A. Three-dimensional in vitro cell culture models in drug discovery and drug repositioning. Front. Pharmacol. 2018, 9, 6. [Google Scholar] [CrossRef]
- Lagies, S.; Schlimpert, M.; Neumann, S.; Wäldin, A.; Kammerer, B.; Borner, C.; Peintner, L. Cells grown in three-dimensional spheroids mirror in vivo metabolic response of epithelial cells. Commun. Biol. 2020, 3, 246. [Google Scholar] [CrossRef]
- Edmondson, R.; Broglie, J.J.; Adcock, A.F.; Yang, L. Three-dimensional cell culture systems and their applications in drug discovery and cell-based biosensors. Assay Drug Dev. Technol. 2014, 12, 207–218. [Google Scholar] [CrossRef] [Green Version]
- El Assal, R.; Gurkan, U.A.; Chen, P.; Juillard, F.; Tocchio, A.; Chinnasamy, T.; Beauchemin, C.; Unluisler, S.; Canikyan, S.; Holman, A.; et al. 3-D microwell array system for culturing virus infected tumor cells. Sci. Rep. 2016, 6, 39144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouchet, B.P.; Achmanova, A. Microtubules in 3D cell motility. J. Cell Sci. 2017, 130, 39–50. [Google Scholar] [CrossRef] [Green Version]
- Eyckmans, J.; Chen, C.S. 3D culture models of tissues under tension. J. Cell Sci. 2017, 130, 63–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakobsson, A.; Ottosson, M.; Zalis, M.C.; O’Carroll, D.; Johansson, U.E.; Johansson, F. Three-dimensional functional human neuronal networks in uncompressed low-density electrospun fiber scaffolds. Nanomedicine 2017, 13, 1563–1573. [Google Scholar] [CrossRef]
- Piccinini, F.; De Santis, I.; Bevilacqua, A. Advances in cancer modeling: Fluidic systems for increasing representativeness of large 3D multicellular spheroids. BioTechniques 2018, 65, 312–314. [Google Scholar] [CrossRef]
- Zhang, K.; Manninen, A. 3D cell culture models of epithelial tissues. Methods Mol. Biol. 2019, 1926, 77–84. [Google Scholar] [PubMed] [Green Version]
- Nickerson, C.A.; Richter, E.G.; Ott, C.M. Studying host-pathogen interactions in 3-D: Organotypic models for infectious disease and drug development. J. Neuroimmune Pharmacol. 2007, 2, 26–31. [Google Scholar] [CrossRef] [Green Version]
- Crabbe, A.; Ledesma, M.A.; Nickerson, C.A. Mimicking the host and its microenvironment in vitro for study in mucosal infections by Pseudomonas aeruginosa. Pathogens Dis. 2014, 71, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardner, J.K.; Herbst-Kralovetz, M.M. Three-dimensional rotating wall vessel-derived cell culture models for studying virus-host interactions. Viruses 2016, 8, 204. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, S.; Clary, J.M.; Seliktar, D.; Lipke, E.A. A three-dimensional spheroidal cancer model based on PEG-fibrinogen hydrogel microspheres. Biomaterials 2017, 115, 141–154. [Google Scholar] [CrossRef]
- Pradhan, S.; Hassani, I.; Seeto, W.J.; Lipke, E.A. PEG-fibrinogen hydrogels for three-dimensional breast cancer cell culture. J. Biomed. Mater. Res. A 2017, 105, 236–252. [Google Scholar] [CrossRef]
- Kim, H.S.; Yoo, H.S. Therapeutic application of electrospun nanofibrous meshes. Nanomedicine 2014, 9, 517–533. [Google Scholar] [CrossRef]
- Prasad, T.; Shabeena, E.A.; Vinod, D.; Kumary, T.V.; Anil Kumar, P.R. Characterization and in vitro evaluation of electrospun chitosan/polycaprolactone blend fibrous mat for skin tissue engineering. J. Mater. Sci. Mater. Med. 2015, 26, 5352–5365. [Google Scholar] [CrossRef]
- Grego, S.; Dougherty, E.R.; Alexander, F.J.; Auerbach, S.S.; Berridge, B.R.; Bittner, M.L.; Casey, W.; Cooley, P.C.; Dash, A.; Ferguson, S.S.; et al. Systems biology for organotypic cell cultures. ALTEX 2017, 34, 301–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalil, S.; El-Badri, N.; El-Mokhtaar, M.; Al-Mofty, S.; Farghaly, M.; Ayman, R.; Habib, D.; Mousa, N. A cost-effective method to assemble biomimetic 3D cell culture platforms. PLoS ONE 2016, 11, e0167116. [Google Scholar]
- Khalil, H.A.; Lei, N.Y.; Brinkley, G.; Scott, A.; Wang, J.; Kar, U.K.; Babaji, Z.B.; Lewis, M.; Martín, M.G.; Dunn, J.C.Y.; et al. A novel culture system for adult porcine intestinal crypts. Cell Tissue Res. 2016, 365, 123–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caballero, D.; Samitier, J. Topological control of extracellular matrix growth: A native-like model for cell morphodynamics studies. ACS Appl. Mater Interfaces 2017, 9, 4159–4170. [Google Scholar] [CrossRef]
- Wang, Y.; DiSalvo, M.; Gunasekara, D.B.; Dutton, J.; Proctor, A.; Lebhar, M.S.; Williamson, I.A.; Speer, J.; Howard, R.L.; Smiddy, N.M.; et al. Self-renewing monolayer of primary colonic or rectal epithelial cells. Cell. Mol. Gastroenterol. Hepatol. 2017, 4, 165–182. [Google Scholar] [CrossRef] [Green Version]
- Dutt, K.; Cao, Y. Attachment to cytodex beads enhances differentiation of human retinal progenitors in 3-D bioreactor culture. Curr. Stem Cell Res. Ther. 2011, 6, 350–361. [Google Scholar] [CrossRef]
- Arranz, A.; Dong, D.; Zhu, S.; Savakis, C.; Tian, J.; Ripolli, J. In-vivo optical tomography of small scattering specimens: Time-lapse 3D imaging of the head eversion process in Drosophilia melanogaster. Sci. Rep. 2014, 4, 7325. [Google Scholar] [CrossRef] [Green Version]
- Järvinen, E.; Muhonen, V.; Haaparanta, A.M.; Kellomäki, M.; Kiviranta, I. Optical projection tomography can be used to investigate spatial distribution of chondrocytes in three-dimensional biomaterial scaffolds for cartilage tissue engineering. Biomed. Mater Eng. 2014, 24, 1549–1553. [Google Scholar] [CrossRef] [PubMed]
- Shamir, E.R.; Ewald, A.J. Three-dimensional organotypic culture: Experimental models of mammalian biology and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 647–663. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Lancaster, M.A.; Castanon, R.; Nery, J.R.; Knoblich, J.A.; Ecker, J.R. Cerebral organoids recapitulate epigenomic signatures of the human fetal brain. Cell Rep. 2016, 17, 3369–3384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bissell, M.J. Goodbye flat biology-time for the 3rd and the 4th dimensions. J. Cell Sci. 2017, 130, 3–5. [Google Scholar] [CrossRef] [Green Version]
- Sakalem, M.E.; Teresa De Sibio, M.; Da Silva da Costa, F.A.; de Oliveira, M. Historical evolution of spheroids and organoids, and possibilities of use in life sciences and medicine. Biotechnol. J. 2021, 16, 2000463. [Google Scholar] [CrossRef]
- Ewald, A.J. 3D cell biology-the expanding frontier. J. Cell Sci. 2017, 130, 1. [Google Scholar] [CrossRef] [Green Version]
- Bhat, U.K.; Yunker, C.E. Establishment and characterization of a diploid cell line from the tick Dermacentor parumapertus Neumann (Acarina: Ixodidae). J. Parasitol. 1977, 63, 1092–1098. [Google Scholar] [CrossRef]
- Roth, J. The silver anniversary of gold: 25 years of the colloidal gold marker system for immunocytochemistry and histochemistry. Histochem. Cell. Biol. 1996, 106, 1–8. [Google Scholar] [CrossRef]
- Zuber, C.; Fan, J.; Guhl, B.; Roth, J. Application of immunogold labelling in ultrastructure pathology. Ultrastruct. Pathol. 2005, 29, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Bairoch, A. The cellosaurus, a cell line knowledge resource. J. Biomol. Tech. 2018, 29, 25–38. [Google Scholar] [CrossRef]
- Suarez, C.E.; Alzan, H.F.; Silva, M.G.; Rathinasamy, V.; Poole, W.A.; Cooke, B.M. Unravelling the cellular and molecular pathogenesis of bovine babesiosis: Is the sky the limit? Int. J. Parasitol. 2019, 49, 183–197. [Google Scholar] [CrossRef]
- Pérez de León, A.A.; Mitchell, R.D., 3rd; Watson, D.W. Ectoparasites of cattle. Vet. Clin. N. Am. Food Anim. Pract. 2020, 36, 173–185. [Google Scholar] [CrossRef]
- Zanoni, M.; Cortesi, M.; Zamagni, A.; Arienti, C.; Pignatta, S.; Tesei, A. Modeling neoplastic disease with spheroids and organoids. J. Hematol. Oncol. 2020, 13, 97. [Google Scholar] [CrossRef] [PubMed]
- Labowska, M.B.; Cierluk, K.; Jankowska, A.M.; Kulbacka, J.; Detyna, J.; Michalak, I. A review on the adaptation of alginate-gelatin hydrogels for 3D cultures and bioprinting. Materials 2021, 14, 858. [Google Scholar] [CrossRef]
- Bierbaumer, L.; Schwarze, U.Y.; Gruber, R.; Neuhaus, W. Cell culture models of oral mucosal barriers: A review with a focus on applications, culture conditions and barrier properties. Tissue Barriers 2018, 6, 1479568. [Google Scholar] [CrossRef] [PubMed]
- Malakpour-Permlid, A.; Buzzi, I.; Hegardt, C.; Johansson, F.; Oredsson, S. Identification of extracellular matrix proteins secreted by human dermal fibroblasts cultured in 3D electrospun scaffolds. Sci. Rep. 2021, 11, 6655. [Google Scholar] [CrossRef] [PubMed]
- Rotstein, B.; Post, Y.; Reinhardt, M.; Lammers, K.; Buhr, A.; Heinisch, J.J.; Meyer, H.; Paulauat, A. Distinct domains in the matricellular protein Lonely heart are crucial for cardiac extracellular matrix formation and heart function in Drosophila. J. Biol Chem. 2018, 293, 7864–7879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnans, C.; Chou, J.; Werb, Z. Remodeling the extracellular matrix in development and disease. Nat. Rev. Mol. Cell. Biol. 2014, 15, 786–801. [Google Scholar] [CrossRef]
- Zhang, T.-T.; Qiu, A.-Q.; Li, Y.; Wang, W.-Y.; Li, M.-M.; Guo, P.; Liu, J.-Z. The mRNA expression and enzymatic activity of three enzymes during embryonic development of the hard tick Haemaphysalis longicornis. Parasit. Vectors 2019, 12, 96. [Google Scholar] [CrossRef]
- Vechtova, P.; Fussy, Z.; Cegan, R.; Sterba, J.; Erhart, J.; Benes, V.; Grubhoffer, L. Catalogue of stage-specific transcripts in Ixodes ricinus and their potential functions during the tick life-cycle. Parasit. Vectors 2020, 13, 311. [Google Scholar] [CrossRef]
- Brown, N.H. Extracellular matrix in development: Insights from mechanisms conserved between invertebrates and vertebrates. Cold Spring Harb Perspect Biol. 2011, 3, a005082. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.; Khan, S.; Ali, I.; Karim, S.; Vaz, I.D.; Termignoni, C. Probing the functional role of tick metalloproteases. Physiol. Entomol. 2015, 40, 177–188. [Google Scholar] [CrossRef] [Green Version]
- Skerrett, I.M.; Williams, J.B. A structural and functional comparison of gap junction channels composed of connexins and innexins. Dev Neurobiol. 2017, 77, 522–547. [Google Scholar] [CrossRef] [PubMed]
- Richards, S.A.; Stutzer, C.; Bosman, A.M.; Maritz-Olivier, C. Transmembrane proteins--Mining the cattle tick transcriptome. Ticks Tick Borne Dis. 2015, 6, 695–710. [Google Scholar] [CrossRef] [Green Version]
- Güiza, J.; Barría, I.; Sáez, J.C.; Vega, J.L. Innexins: Expression, Regulation, and Functions. Front. Physiol. 2018, 9, 1414. [Google Scholar] [CrossRef]
- Baker, B.M.; Chen, C.B. Deconstructing the third dimension-how 3D culture microenvironments alter cellular clues. J. Cell Sci. 2012, 125, 3015–3024. [Google Scholar] [PubMed] [Green Version]
- Wrzesinski, K.; Rogowsk-Wrzesinka, A.; Kanlaya, R.; Borkowski, K.; Schwämmle, V.; Dai, J.; Joensen, K.E.; Wojdyla, K.; Carvalho, V.B.; Fey, S.J. The cultural divide: Exponential growth in classical 2D and metabolic equilibrium in 3D environments. PLoS ONE 2014, 9, e106973, Erratum in: PLoS ONE 2021, 10, e0118050. [Google Scholar] [CrossRef] [Green Version]
- Dance, A. AI spots cell structures that humans can’t. Nature 2021, 592, 154–155. [Google Scholar] [CrossRef]
- Yu, J.H.; Liu, X.L.; Liu, Y.J.; He, X.E.; Hui, Y.; Zhang, B.; Zhao, W. Three-dimensional morphology of C6/36 cells infected by dengue virus: A study based on digital holographic microscopy. J. Southern Med. Univ. 2017, 37, 301–307. [Google Scholar]
- Yardimci, S.; Burnham, D.R.; Terry, S.Y.A.; Yardimci, H. Three-dimensional super-resolution fluorescence imaging of DNA. Sci. Rep. 2020, 10, 12504. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, J.; Ahlgren, U.; Perry, P.; Hill, B.; Ross, A.; Hecksher-Sørensen, J.; Baldock, R.; Davidson, D. Optical projection tomography as a tool for 3D microscopy and gene expression studies. Science 2002, 296, 541–545. [Google Scholar] [CrossRef] [Green Version]
- Kuś, A.; Dudek, M.; Kemper, B.; Kujawinska, M.; Vollmer, A. Tomographic phase microscopy of living three-dimensional cell cultures. J. Biomed. Opt. 2014, 19, 046009. [Google Scholar] [CrossRef]
- Figueiras, E.; Soto, A.M.; Jesus, D.; Lehti, M.; Koivist, J.; Parraga, J.E.; Silva-Correia, J.; Oliveira, J.M.; Reis, R.I.; Kellomäki, M.; et al. Optical projection tomography as a tool for 3D imaging of hydrogels. Biomed. Opt. Express. 2014, 5, 3443–3449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabadi, P.K.; Vantangoli, M.M.; Rodd, A.L.; Leary, E.; Madnick, S.J.; Morgan, J.R.; Kane, A.; Boekelheide, K. Into the depths: Techniques for in vitro three-dimensional visualization. BioTechniques 2015, 59, 279–286. [Google Scholar] [CrossRef]
- de Abreu, L.A.; Fabres, A.; Esteves, E.; Masuda, A.; Vaz, I.S.; Dffre, S.; Logullo, C. Exogenous insulin stimulates glycogen accumulation in IRhipicephaalus (Boophilus) microplus embryo cell line BME26 vi P13/AKT pathway. IComp. Biochem. Physiol. Part B 2009, 153, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Voronin, D.; Tricoche, N.; Jawahar, S.; Shlossman, M.; Bulman, C.A.; Fischer, C.; Suderman, M.T.; Sakanari, J.A.; Lustigman, S. Development of a preliminary in vitro drug screening assay based on a newly established culturing system for pre-adult fifth stage Onchocerca volvulus worms. PLoS Negl. Trop. Dis. 2019, 13, e0007108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Killmaster, L.F.; Levin, M.L. Isolation and short-term persistence of Ehrlichia ewingii in cell culture. Vector Borne Zoonotic Dis. 2016, 16, 445–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunz-Schughart, L.A.; Wenninger, S.; Neumeier, T.; Seidl, P.; Knuechel, R. Three-dimensional tissue structure affects sensitivity of fibroblasts to TGF-β1. Am. J. Physiol. Cell Physiol. 2003, 284, C209–C219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chawla, S.; Chameettachal, S.; Ghosh, S. Probing the role of scaffold dimensionality and media composition on matrix production and phenotype of fibroblasts. Mater. Sci. Eng. C Mater. Biol. Appl. 2015, 49, 588–596. [Google Scholar] [CrossRef]
- Ku, S.H.; Park, C.B. Human endothelial cell growth on mussel-inspired nanofiber scaffold for vascular tissue engineering. Biomaterials 2010, 31, 9431–9437. [Google Scholar] [CrossRef]
- Fang, Y.; Eglen, R.M. Three-Dimensional Cell Cultures in Drug Discovery and Development. SLAS Discov. 2017, 22, 456–472. [Google Scholar]
- Biswas, N.; Kumar, K.; Bose, S.; Bera, R.; Chakrabarti, S. Analysis of Pan-omics Data in Human Interactome Network (APODHIN). Front. Genet. 2020, 11, 589231. [Google Scholar] [CrossRef]
- Gross, A.D.; Temeyer, K.B.; Day, T.A.; Pérez de León, A.A.; Kimber, M.J.; Coats, J.R. Interaction of plant essential oil terpenoids with the southern cattle tick tyramine receptor: A potential biopesticide target. Chem. Biol. Interact. 2017, 263, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Bhowmick, B.; Han, Q. Understanding Tick Biology and Its Implications in Anti-tick and Transmission Blocking Vaccines Against Tick-Borne Pathogens. Front. Vet. Sci. 2020, 7, 319, Erratum in: Front. Vet. Sci. 2020, 7, 575. [Google Scholar] [CrossRef] [PubMed]
- Yi, H.G.; Kim, H.; Kwon, J.; Choi, Y.J.; Jang, J.; Cho, D.W. Application of 3D bioprinting in the prevention and the therapy for human diseases. Signal Transduct Target Ther. 2021, 6, 177. [Google Scholar] [CrossRef]
- Almazan, C.; Tipacamu, G.A.; Rodriguez, S.; Mosqueda, J.; Pérez de León, A. Immunological control of ticks and tick-borne diseases that impact cattle health and production. Front. Biosci. 2018, 23, 1535–1551. [Google Scholar] [CrossRef] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
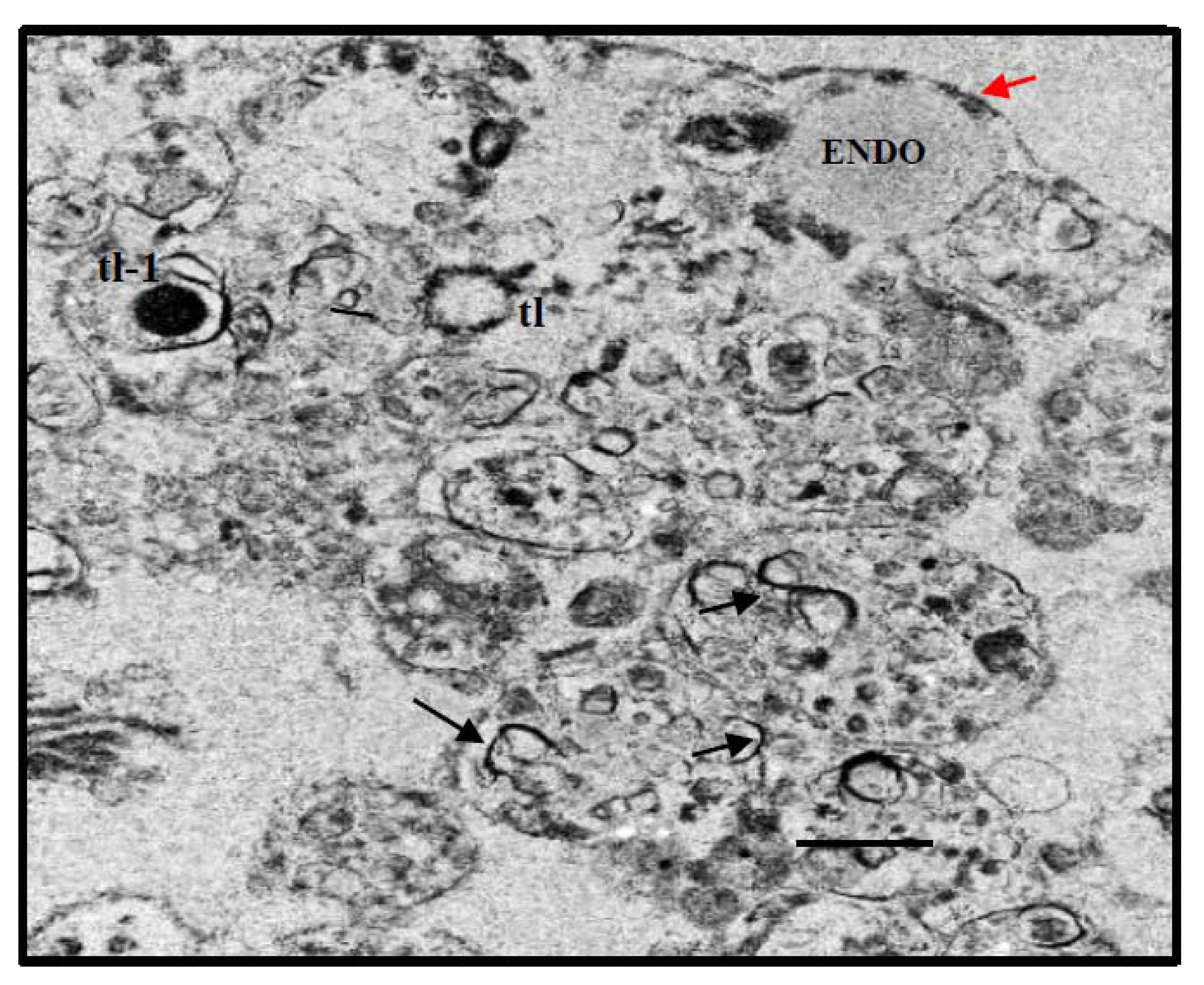{kind=link}
{kind=link}
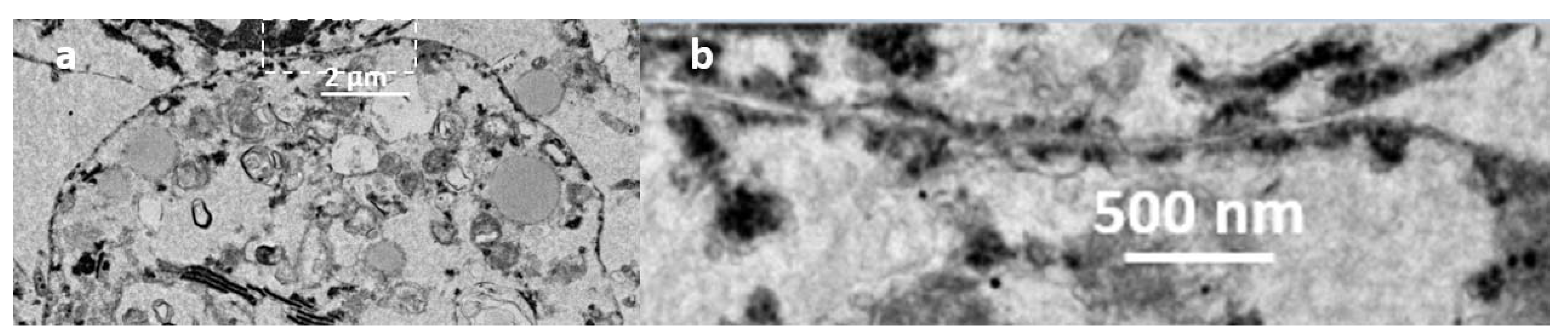{kind=link}
| Day | 0 | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|---|
| Cells/Ml * | 5.0 × 105 | 5.9 × 105 | 6.4 × 105 | 7.3 × 105 | 8.1 × 105 | 9.1 × 105 | 1.3 × 106 |
| Cell Morphology | Day 30 (2-D) | Day 10 Hydrogel | Day 25 Hydrogel | Day 30 Hydrogel | Day 40 Hydrogel |
|---|---|---|---|---|---|
| Fibrocyte-like | 39 µm ± 9.25 µm * (27–55 µm) | N.D. | N.D. | N.D. | N.D. |
| Small Granular Endothelial-like | 16 µm ± 3.25 µm (12–25 µm) | N.D. | 18.3 µm ± 1.09 µm (17–20 µm) | 18 µm ± 2.07 µm (16–21 µm) | 15 µm ± 0.8 µm (14–16 µm) |
| Large with cytoplasmic vacuoles | 33 µm ± 6.65 µm (12–40 µm) | N.D. | 31.8 µm ± 5.90 µm (21–34 µm) | 24.5 µm ± 2.36 µm (23–33 µm) | 36.4 µm ± 2.0 µm (34–39 µm) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suderman, M.T.; Temeyer, K.B.; Schlechte, K.G.; Pérez de León, A.A. Three-Dimensional Culture of Rhipicephalus (Boophilus) microplus BmVIII-SCC Cells on Multiple Synthetic Scaffold Systems and in Rotating Bioreactors. Insects 2021, 12, 747. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12080747
Suderman MT, Temeyer KB, Schlechte KG, Pérez de León AA. Three-Dimensional Culture of Rhipicephalus (Boophilus) microplus BmVIII-SCC Cells on Multiple Synthetic Scaffold Systems and in Rotating Bioreactors. Insects. 2021; 12(8):747. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12080747
Chicago/Turabian StyleSuderman, Michael T., Kevin B. Temeyer, Kristie G. Schlechte, and Adalberto A. Pérez de León. 2021. "Three-Dimensional Culture of Rhipicephalus (Boophilus) microplus BmVIII-SCC Cells on Multiple Synthetic Scaffold Systems and in Rotating Bioreactors" Insects 12, no. 8: 747. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12080747