
Persistency of Indigenous and Exotic Entomopathogenic Fungi Isolates under Ultraviolet B (UV-B) Irradiation to Enhance Field Application Efficacy and Obtain Sustainable Control of the Red Palm Weevil



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Entomopathogenic Fungi Source
2.2. Isolation and Culture of Fungal Isolates on Potato Dextrose Agar Medium
2.3. Sterilization and Disinfection Procedure
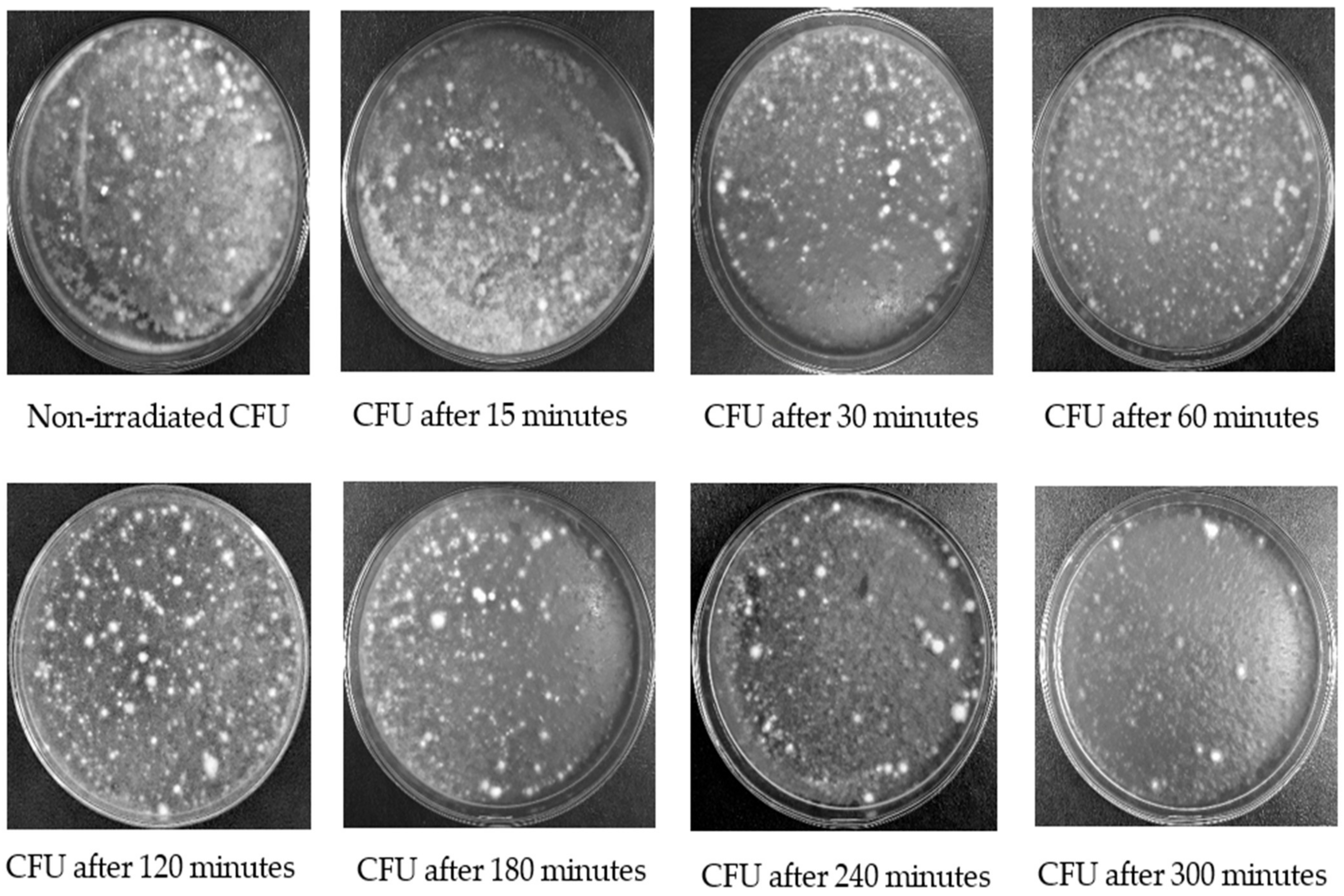
2.4. Evaluation of the Persistence of Fungi Isolates under UV-B Exposures
2.5. Colony-Forming Unit
2.6. Statistical Analysis
3. Results
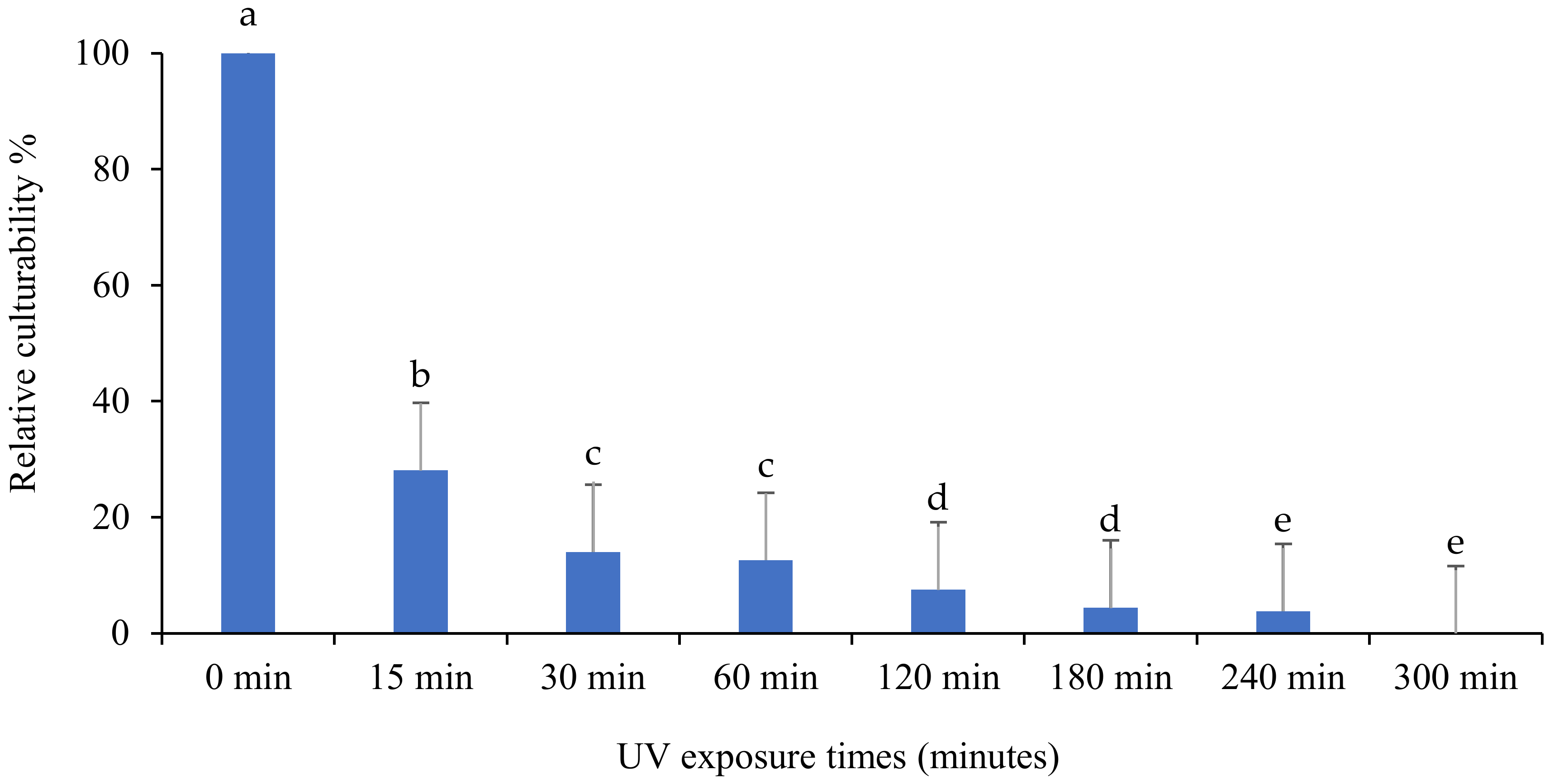
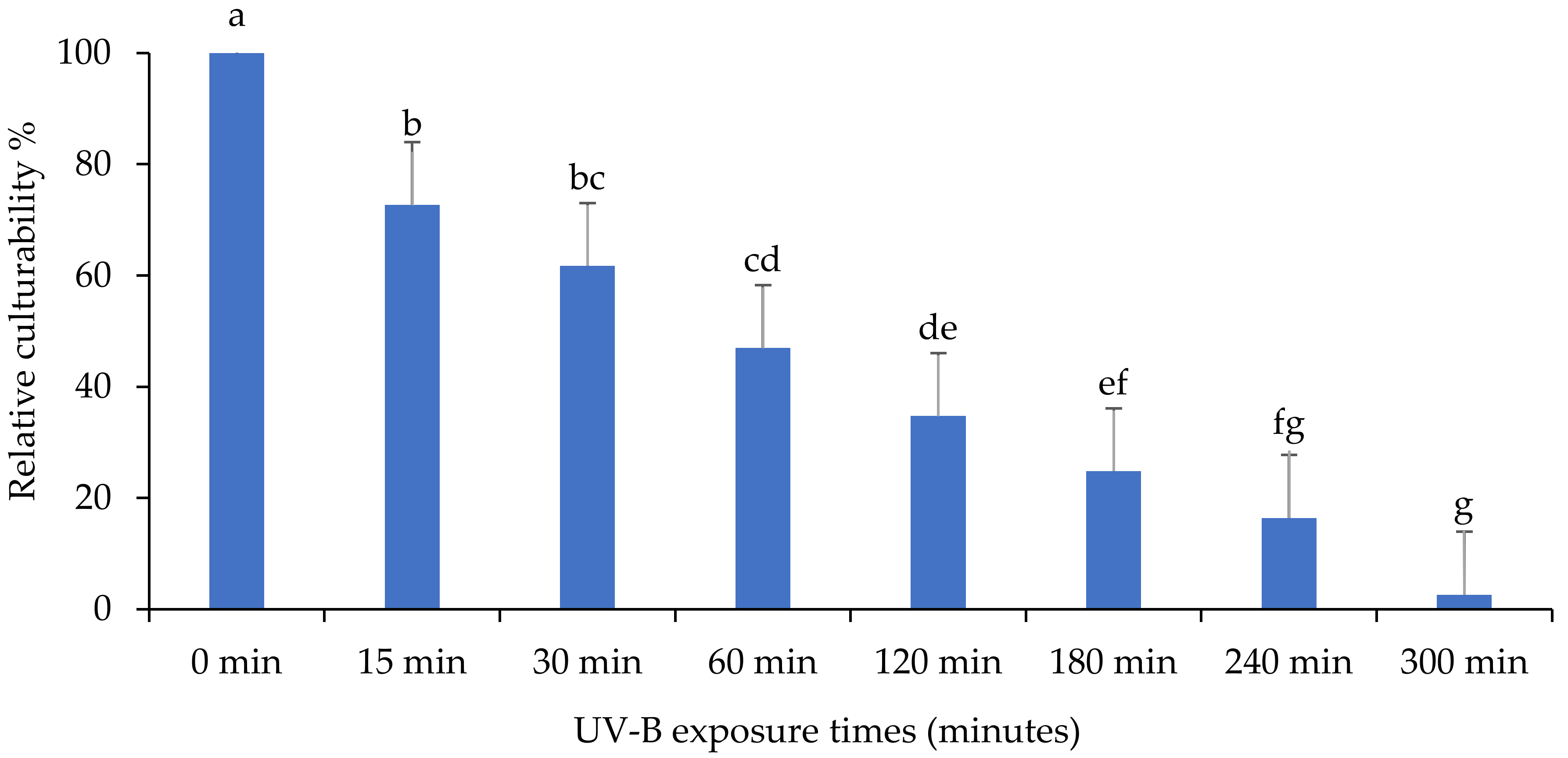
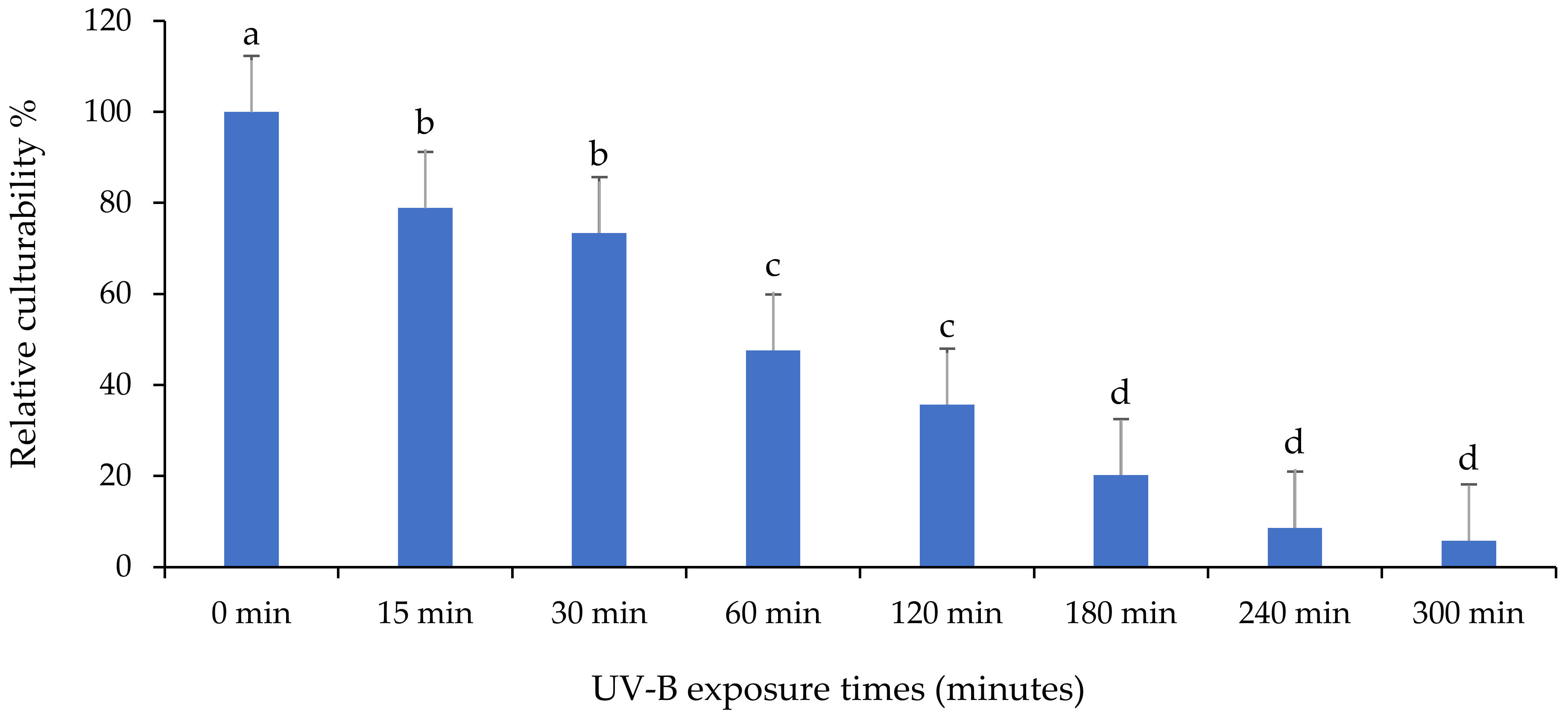
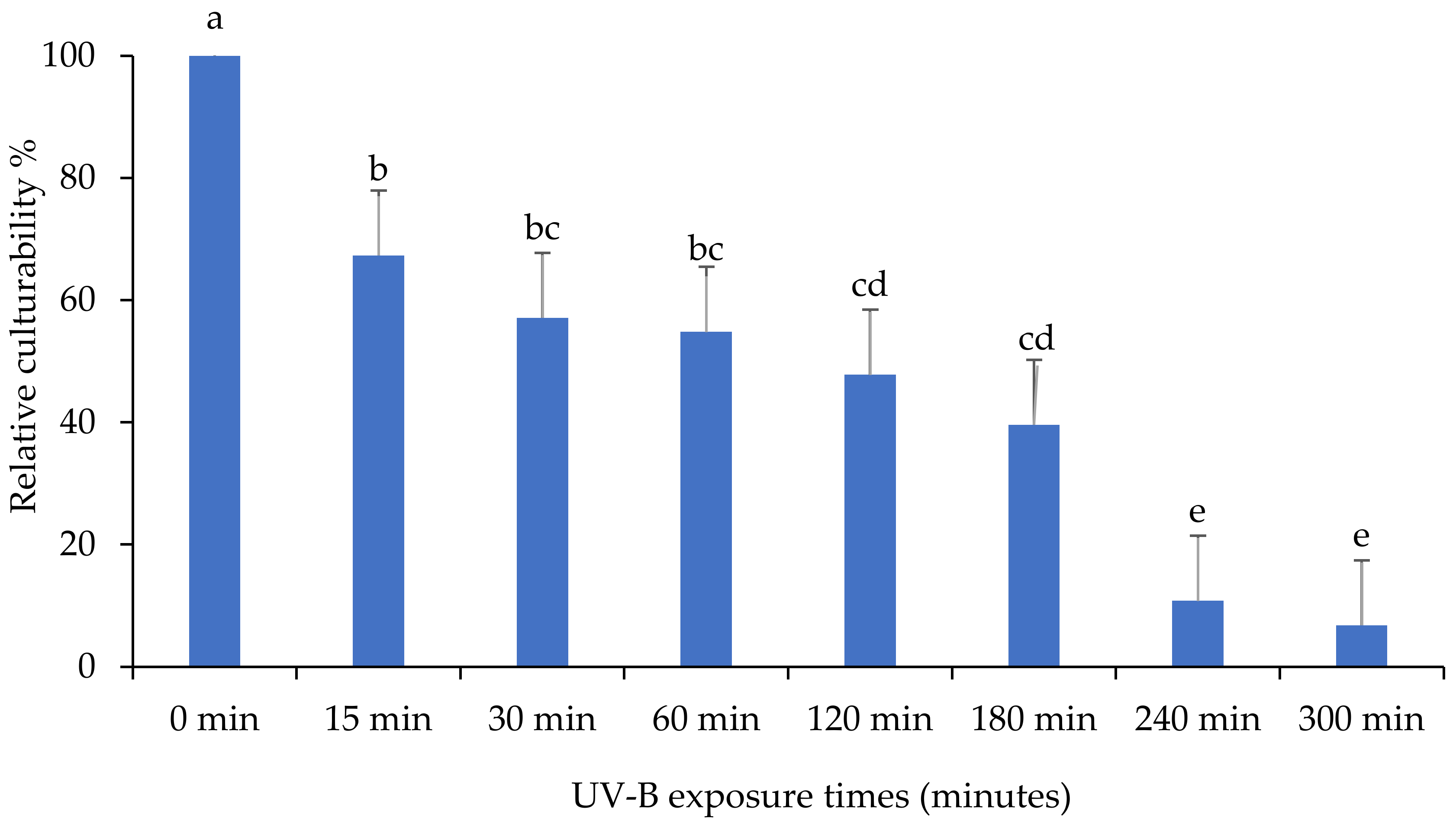
3.1. Evaluation of the Persistence of Fungi Isolates under UV-B Exposures
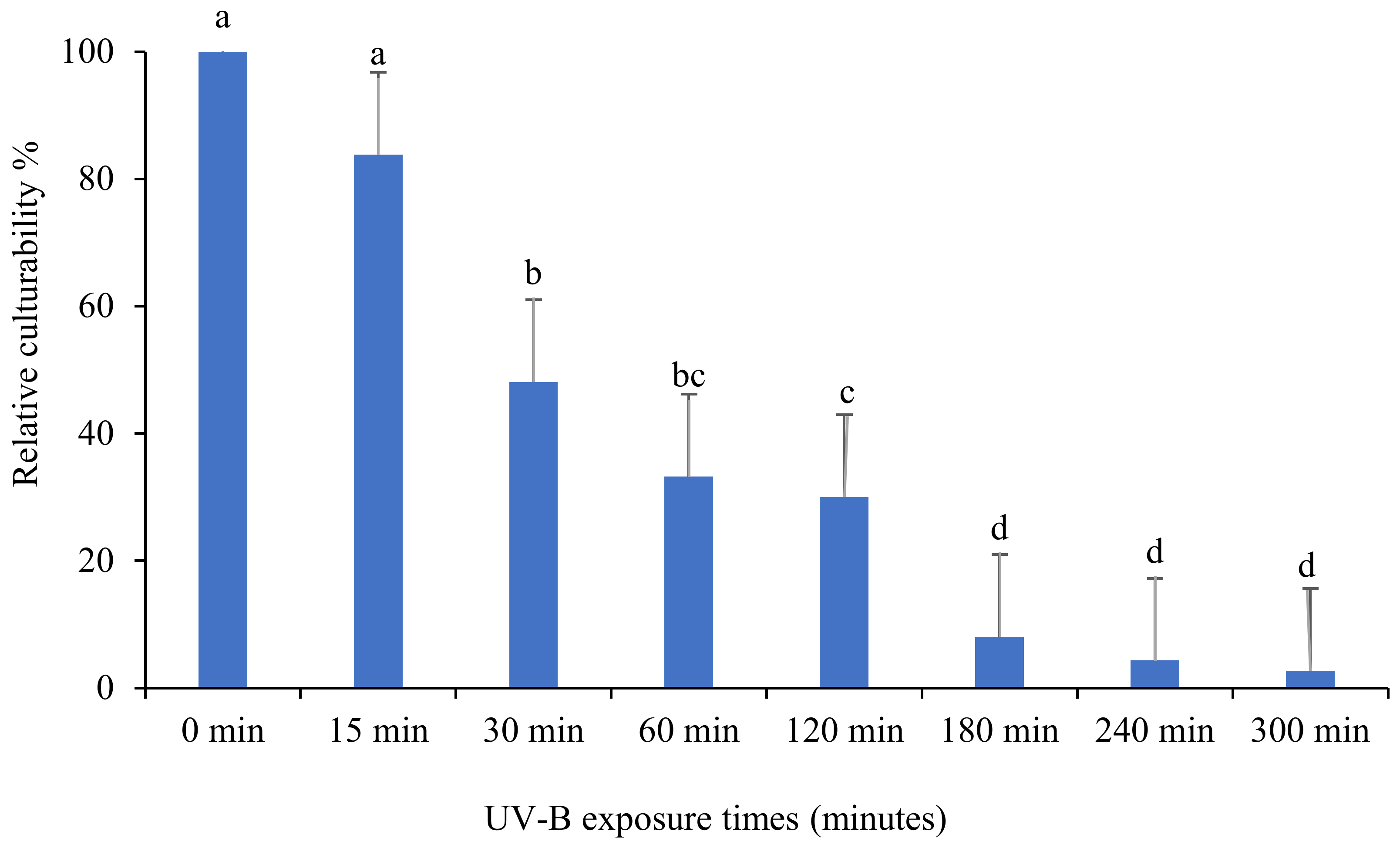
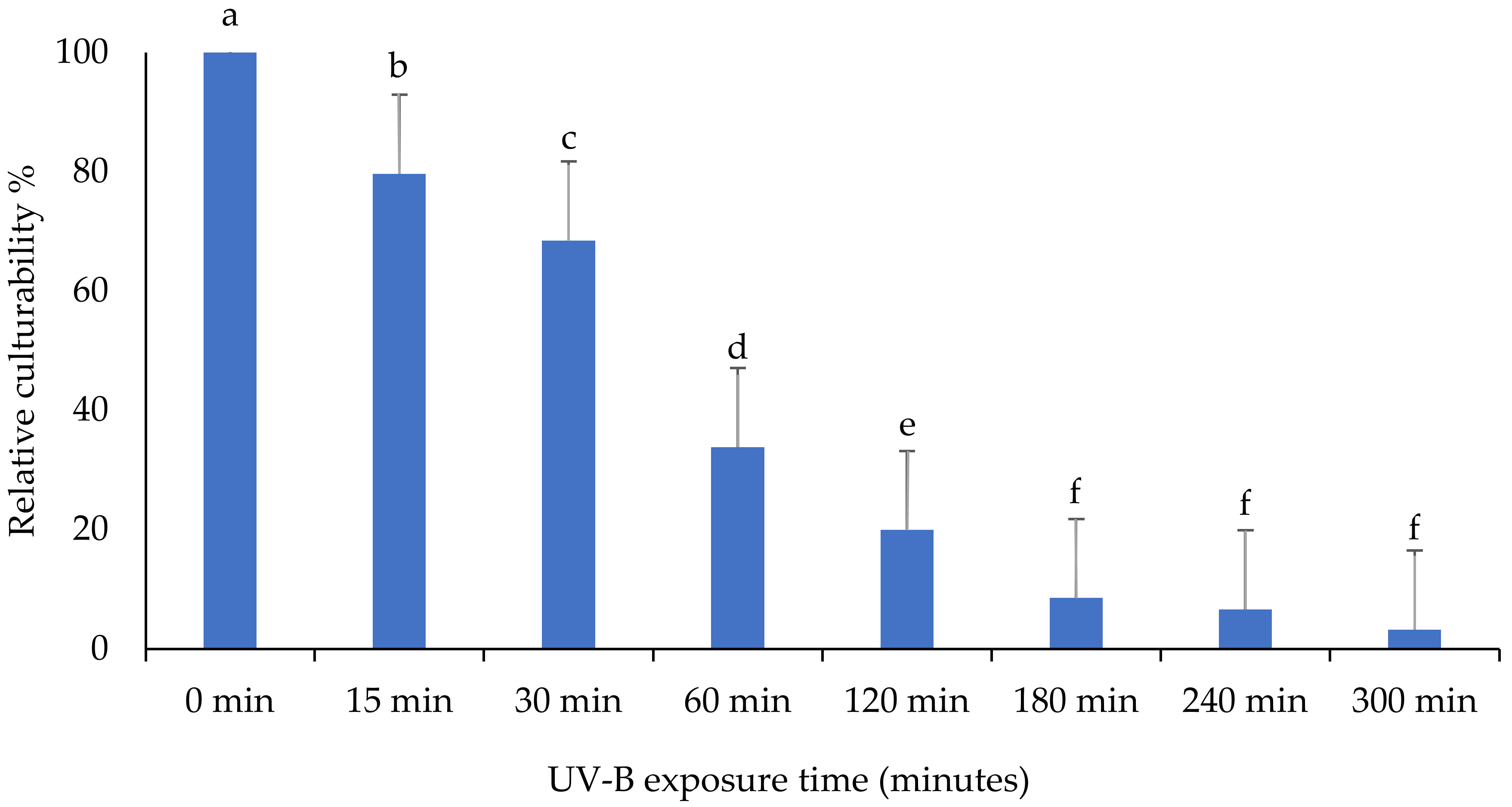
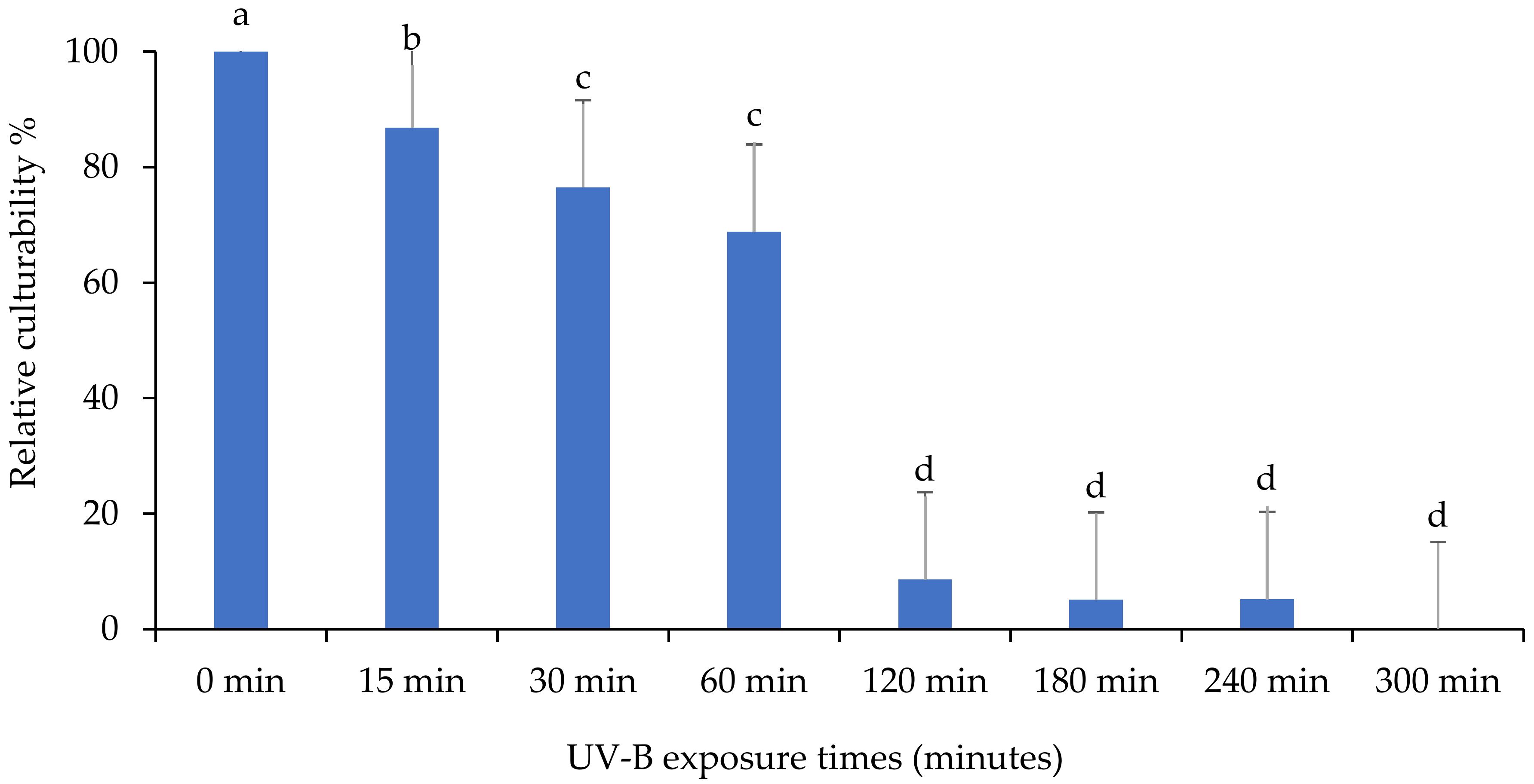
3.2. Relative Culturability of Fungi Isolates after Ultraviolet B (UV-B) Irradiation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vidyasagar, P.S.P.V.; Al Saihati, A.A.; Al Mohanna, O.E.; Subbei, A.I.; Abdul Mohsin, A.M. Management of red palm weevil Rhynchophorus ferrugineus Oliv., A serious pest of date palm in Al Qatif, Kingdom of Saudi Arabia. J. Plant Crops. 2000, 28, 35–43. [Google Scholar]
- Cutler, T.D.; Zimmerman, J.J. Ultraviolet irradiation and the mechanisms underlying its inactivation of infectious agents. Anim. Health Res. Rev. 2011, 12, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, U.; Das, D.; Banerjee, R.K. Reactive oxygen species: Oxidative damage and pathogenesis. Review Article. Curr. Sci. 1999, 77, 658–666. [Google Scholar]
- Schuch, A.P.; Morenob, N.C.; Schuchc, N.J.; Menckb, C.F.M.; Garciad, C.C.M. Sunlight damage to cellular DNA: Focus on oxidatively generated lesions. Free Radic. Biol. Med. 2017, 107, 110–124. [Google Scholar] [CrossRef]
- Chelico, L.; Haughian, J.L.; Woytowich, A.E.; Khachatourians, G.G. Quantification of Ultraviolet-C irradiation induced cyclobutane pyrimidine dimers and their removal in Beauveria bassiana conidiophores DNA. Mycologia 2005, 97, 621–627. [Google Scholar] [CrossRef]
- Braga, G.U.L.; Rangel, D.E.N.; Flint, S.D.; Miller, C.D.; Anderson, A.J.; Roberts, D.W. Damage and recovery from UV-B exposure in conidia of the enthomopathogens Verticilium lecanii and Aphanocladium album. Mycologia 2002, 94, 912–920. [Google Scholar] [CrossRef]
- Huarte-Bonnet, C.; Pereira-Junior, R.A.; Paixao, F.R.A.; Braga, G.U.L.; Roberts, D.W.; Luz, C.; Pedrini, N.; Fernandes, E.K.K. Metarhizium robertsii and M. acridum conidia produced on riboflavin-supplemented medium have increased UV-A tolerance and upregulated photoprotection and photoreactivation genes. BioControl 2020, 65, 211–222. [Google Scholar] [CrossRef]
- Kaiser, D.; Bacher, S.; Mène-Saffrané, L.; Grabenwegera, G. Efficiency of natural substances to protect Beauveria bassiana conidia from UV radiation. Pest Manag. Sci. 2018, 75, 556–563. [Google Scholar] [CrossRef]
- Herlinda, S.; Rizkie, L.; Suwandi; Susilawati; Lakitan, B.; Verawaty, M.; Hasbi. Effects of High Temperature and Ultraviolet-C Irradiance on Conidial Viability and Density of Beauveria Bassiana and Metarhizium Anisopliae Isolated from Soils of Lowland Ecosystems in Indonesia. Eurasian J. Anal Chem. 2018, 13, 209–216. [Google Scholar]
- Couceiro, J.D.C.; Fatoretto, M.B.; Demétrio, C.G.B.; Meyling, N.V.; Delalibera, Í.J. UV-B Radiation Tolerance and Temperature-Dependent Activity within the Entomopathogenic Fungal Genus Metarhizium in Brazil. Front. Fungal Biol. 2021, 2, 6. [Google Scholar] [CrossRef]
- Fernandes, É.K.K.; Rangel, D.E.N.; Braga, G.U.; Roberts, D.W. Tolerance of fungi to ultraviolet radiation: A review on screening of strains and their formulation. Curr. Genet. 2015, 61, 427–440. [Google Scholar] [CrossRef]
- Francardi, V.; Benvenuti, C.; Paolobarzanti, G.; Roversi, P.F. Auto-contamination trap with fungi: A possible strategy in the control of Rhynchophorus ferrugineus (Oliver) (Coleoptera: Curculionidae). Redia 2013, 96, 57–67. [Google Scholar]
- Le Grand, M.; Cliquet, S. Impact of culture age on conidial germination, desiccation and UV tolerance of entomopathogenic fungi. Biocontrol Sci. Technol. 2013, 23, 847–859. [Google Scholar] [CrossRef]
- Cagáň, Ľ.; Švercel, M. The influence of ultraviolet light on pathogenicity of entomopathogenic fungi Beauveria bassiana (balsamo) vuillemin to the european corn borer, Ostrinia nubilalis hbn. (Lepidoptera: Crambidae). J. Cent. Eur. Agric. 2021, 2, 228–234. [Google Scholar] [CrossRef]
- Tobar, S.P.; Vélez-Arango, P.E.; Montoya-Restrepo, E.C. Evaluación en campo de un aislamiento de Beauveria bassiana seleccionado por resistencia a la luz ultravioleta. Cenicafé 1999, 50, 195–204. [Google Scholar]
- Humber, R.A. Fungi: Identification Manual of Techniques in Insect Pathology; Academic Press: New York, NY, USA, 2012; pp. 151–187. [Google Scholar]
- Sutanto, K.D.; Husain, M.; Rasool, K.G.; Al-Qahtani, W.H.; Aldawood, A.S. Pathogenicity of local and exotic entomopathogenic fungi isolates against different life stages of red palm weevil (Rhynchophorus ferrugineus). PLoS ONE 2021, 16, e0255029. [Google Scholar] [CrossRef]
- Sahayaraj, K.; Namasivayam, S.K.R. Mass production of fungi using agricultural products and by products. Afr. J. Biotechnol. 2008, 7, 1907–1910. [Google Scholar] [CrossRef]
- Gutierrez-Coarite, R.; Javier, M.; Lisa, K.; Wade, P.H.; Lionel, S.; Mark, G.W.; Stacey, C. Entomopathogenic Fungi as Mortality Factors of Macadamia Felted Coccid, Eriococcus ironsidei (Hemiptera: Eriococcidae) in Hawaii. Proc. Hawaii. Entomol. Soc. 2018, 50, 9–16. [Google Scholar]
- El Husseini, M.M. Efficacy of the fungus Beauveria bassiana (Balsamo) Vuillemin on the red palm weevil Rhynchophorus ferrugineus Olivier (Coleoptera: Curculionidae) larvae and adults under laboratory conditions. Egypt J. Biol. Pest Control 2019, 29, 58. [Google Scholar] [CrossRef]
- Oliveira, D.G.P.; Pauli, G.; Mascarin, G.M.; Delalibera, I. A protocol for determination of conidial viability of the fungal entomopathogens Beauveria bassiana and Metarhizium anisopliae from commercial products. J. Microbiol. Methods 2015, 119, 44–52. [Google Scholar] [CrossRef] [Green Version]
- SAS. SAS/STAT® 9.1. Users Guide; SAS Institute: Cary, NC, USA, 2004; pp. 1–5136. [Google Scholar]
- Braga, G.U.L.; Flint, S.D.; Messias, C.L.; Anderson, A.J.; Roberts, D.W. Effects of UVB Irradiance on Conidia and Germinants of the Entomopathogenic Hyphomycete Metarhizium anisopliae: A Study of Reciprocity and Recovery. Photochem. Photobiol. 2001, 73, 140–146. [Google Scholar] [CrossRef]
- Diffey, B.L. Solar ultraviolet radiation effects on biological system. Phys. Med. Biol. 1991, 36, 299. [Google Scholar] [CrossRef]
- Alves, R.T.; Bateman, R.P.; Prior, C.; Leather, S.R. Effects of simulated solar radiation on conidial germination of Metarhizium anisopliae in different formulations. Crop Prot. 1998, 17, 675–679. [Google Scholar] [CrossRef]
- Braga, G.U.L.; Rangel, D.E.N.; Fernandes, E.K.K.; Roberts, D.W. Molecular and physiological effects of environmental UV radiation on fungal conidia. Curr. Genet. 2015, 61, 405–425. [Google Scholar] [CrossRef]
- Ottati-De-Lima, E.L.; Filho, A.B.; Almeida, J.E.M.D.; Gassen, M.H.; Wenzel, I.M.; Almeida, A.M.B.D.; Zapellini, L.O. Liquid production of entomopathogenic fungi and ultraviolet radiation and temperature effects on produced propagules. Arq. Inst. Biol. 2014, 81, 342–350. [Google Scholar] [CrossRef]
- Morley-Davies, J.; Moore, D.; Prior, C. Screening of Metarhizium and Beauveria spp. conidia with exposure to simulated sunlight and a range of temperatures. Mycol. Res. 1995, 100, 31–38. [Google Scholar] [CrossRef]
- Fang, W.; St Leger, R.J. Enhanced UV Resistance and Improved Killing of Malaria Mosquitoes by Photolyase Transgenic Entomopathogenic Fungi. PLoS ONE 2012, 7, e0043069. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, N.; Walker, G.C. Mechanisms of DNA Damage, Repair, and Mutagenesis. Environ. Mol. Mutagen. 2017, 58, 235–263. [Google Scholar] [CrossRef] [Green Version]
- Braga, G.U.L.; Rangel, D.E.N.; Flint, S.D.; Anderson, A.J.; Roberts, D.W. Conidial Pigmentation Is Important to Tolerance Against Solar-simulated Radiation in the Entomopathogenic fungi Metarhizium anisopliae. Photochem. Photobiol. 2006, 82, 418–422. [Google Scholar] [CrossRef]
- Hu, W.; Hou, R.F.; Talekar, N.S. Pathogenicity of Beauveria bassiana to Riptortus linearis (Hemiptera: Coreidae), a pest of soybean. Appl. Entomol. Zool. 1996, 31, 187–194. [Google Scholar] [CrossRef] [Green Version]
- Loong, C.Y.; Sajap, A.S.; Noor, H.M.; Omar, D.; Abood, F. Effects of UV-B and Solar Radiation on the Efficacy of Isaria fumosorosea and Metarhizium anisopliae (Deuteromycetes: Hyphomycetes) for Controlling Bagworm, Pteroma pendula (Lepidoptera: Psychidae). J. Entomol. 2013, 10, 53–65. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, I.; Forim, M.; Silva, M.; Fernandes, J.; Filho, A. Effect of Ultraviolet Radiation on Fungi Beauveria bassiana and Metarhizium anisopliae, Pure and Encapsulated, and Bio-Insecticide Action on Diatraea saccharalis. Adv. Entomol. 2016, 4, 151–162. [Google Scholar] [CrossRef] [Green Version]
- Acheampong, M.A.; Hill, M.P.; Moore, S.D.; Coombes, C.A. UV sensitivity of Beauveria bassiana and Metarhizium anisopliae isolates under investigation as potential biological control agents in South African citrus orchards. Fungal Biol. 2020, 124, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.A.; Burges, H.G. Introduction. In Formulation of Microbial Biopesticides, Beneficial Microorganisms, Nematodes and Seed Treatments; Burges, H.D., Ed.; Kluwer Academic: London, UK, 1998; pp. 1–4. [Google Scholar]
- Brancini, G.T.P.; Rangel, D.E.N.; Braga, G.Ú.L. Exposure of Metarhizium acridum mycelium to light induce tolerance to UV-B radiation. Microbiol. Lett. 2016, 363, fnw036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rangel, D.E.N.; Roberts, D.W. Inducing UV-B tolerance of Metarhizium anisopliae var. anisopliae conidia results in a tradeoff between conidial production and conidial stress tolerance. J. Anhui Agric. Univ. 2007, 34, 195–202. [Google Scholar]
- Mustafa, U.; Kaur, G. UV-B radiation and temperature stress causes variable growth response in Metarhizium anisopliae and Beauveria bassiana isolates. Internet J. Microbiol. 2008, 7, 1–8. [Google Scholar] [CrossRef]
- Fargues, J.; Goettel, M.S.; Smits, N.; Ouedraogo, A.; Vidal, C.; Lacey, L.L.; Lomer, C.J.; Rougier, M. Variability in susceptibility to simulated sunlight of conidia among isolates of entomopathogenic Hyphomycetes. Mycopathologia 1996, 135, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Rangel, D.E.N.; Braga, G.U.L.; Anderson, A.J.; Roberts, D.W. Variability in conidial thermotolerance of Metarhizium anisopliae isolates from different geographic origins. J. Invertbr. Pathol. 2005, 88, 116–125. [Google Scholar] [CrossRef]
- Santoro, P.H.; Zorzetti, J.; Constanski, K.; Neves, P.M.O.J. Quality of Beauveria bassiana conidia after successive passages through Alphitobius diaperinus (Coleoptera: Tenebrionidae). Rev. Colomb. Entomol. 2015, 41, 87–94. [Google Scholar]
- Mazza, G.; Francardi, V.; Simoni, S.; Benvenuti, C.; Cervo, R.; Faleiro, J.R.; Llacer, E.; Longo, S.; Nannelli, R.; Tarasco, E.; et al. An overview on the natural enemies of Rhynchophorus palm weevils, with focus on R. ferrugineus. Biol. Control 2014, 77, 83–92. [Google Scholar] [CrossRef]
- Tarasco, E.; Porcelli, F.; Poliseno, M.; Quesada, M.E.; Santiago, Á.C.; Triggiani, O. Natural occurrence of entomopathogenic fungi infecting the Red Palm Weevil Rhynchophorus ferrugineus (Olivier, 1790) (Coleoptera: Curculionidae) in southern Italy. IOBC/WPRS Bull. 2008, 31, 195–197. [Google Scholar]
- Triggiani, O.; Tarasco, E. Evaluation of the effects of autochthonous and commercial isolates of Steinernematidae and Heterorhabditidae on Rhynchophorus ferrugineus. Bull. Insectology 2011, 64, 175–180. [Google Scholar]
- Dyah, R.I.; Priyantini, W.; Haryuni, M.S.; Yoris, A.M. Effectiveness of M. anisopliae and entomopathogenic nematodes to control Oryctes rhinoceros larvae in the rainy season. Pak. J. Biol. Sci. 2017, 20, 320–327. [Google Scholar]
- Fernandes, É.K.K.; Rangel, D.E.N.; Moraes, A.M.L.; Bittencourt, V.R.E.P.; Roberts, D.W. Variability in tolerance to UV-B radiation among Beauveria spp. isolates. J. Invertebr. Pathol. 2007, 96, 237–243. [Google Scholar] [CrossRef]
- Rangel, D.E.N.; Butler, M.J.; Torabinejad, J.; Anderson, A.J.; Braga, G.U.L.; Day, A.W.; Roberts, D.W. Mutants and isolates of Metarhizium anisopliae are diverse in their relationships between conidial pigmentation and stress tolerance. J. Invertbr. Pathol. 2006, 93, 170–182. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungi | Isolates’ Code | Insect Species/ Order | Source of Isolation | Original Location (s) of Isolates | Coordinates |
|---|---|---|---|---|---|
| B. bassiana | BbSA-1 | Red palm weevil R. ferrugineus/Coleoptera | adult | Al Qatif, Saudi Arabia | N: 26.34437°; E: 43.69217° |
| B. bassiana | BbSA-2 | Cotton leafworm Spodoptera litura /Lepidoptera | larva | Al Qatif, Saudi Arabia | N: 24.41867°; E: 46.65408° |
| B. bassiana | BbSA-3 | Red palm weevil R. ferrugineus/Coleoptera | adult | Al Qatif, Saudi Arabia | N: 26.35231°; E: 43.71789° |
| B. bassiana | BbIDN-1 | Corn earworm Helicoverpa zea /Lepidoptera | larva | Kalimantan, Indonesia | N: −0.02633°; E: 109.3425° |
| M. anisopliae | MaSA-1 | Red palm weevil R. ferrugineus /Coleoptera | adult | Riyadh, Saudi Arabia | N: 24.41867°; E: 46.65408° |
| M. anisopliae | MaIDN-1 | Coconut Rhinoceros beetle Oryctes rhinoceros/Coleoptera | adult | Kalimantan, Indonesia | N: −0.02633°; E: 109.3425° |
| M. anisopliae | MaIDN-2 | Coconut leaf beetle Brontispa longissima/Coleoptera | adult | Kalimantan, Indonesia | N: −0.03962°; E: 109.3128° |
| Isolate Name | Isolate Code | Colony-Forming Unit (CFU)/Ml of Fungus Isolate | |||||||
|---|---|---|---|---|---|---|---|---|---|
| UV-B Exposure Times | |||||||||
| 0 Min | 15 Min | 30 Min | 60 Min | 120 Min | 180 Min | 240 Min | 300 Min | ||
| B. bassiana | BbSA-1 | 433 ± 30 a | 355 ± 32 b | 203 ± 26 c | 143 ± 7 d | 127 ± 10 d | 33 ± 5 e | 18 ± 4 e | 11.6 ± 3 e |
| BbSA-2 | 311 ± 5 a | 247 ± 16 b | 212 ± 17 c | 105 ± 14 d | 62 ± 3 e | 26 ± 1 f | 20 ± 3 f | 10 ± 1 f | |
| BbSA-3 | 368 ± 30 a | 323 ± 45 ab | 275 ± 23 bc | 225 ± 28 c | 31 ± 4 d | 19 ± 3 d | 20 ± 2d | 0 ± 0 d | |
| BbIDN-1 | 340 ± 13 a | 95 ± 10 b | 48 ± 3 c | 43 ± 7 cd | 25 ± 2 de | 15 ± 2 ef | 13 ± 0.1 ef | 0 ± 0 f | |
| M. anisopliae | MaIDN-1 | 420 ± 30 a | 305 ± 20 b | 259 ± 10 bc | 197 ± 36 cd | 146 ± 2 de | 104 ± 25 ef | 69 ± 9 fg | 11 ± 0.5 g |
| MaIDN-2 | 453 ± 29 a | 355 ± 31 b | 328 ± 17 b | 213 ± 8 c | 162 ± 26 c | 89 ± 7 d | 39 ± 8 de | 25 ± 3 e | |
| MaSA-1 | 439 ± 24 a | 293 ± 9 b | 250 ± 10 bc | 238 ± 13 c | 207 ± 14 cd | 171 ± 23 d | 47 ± 6 e | 29 ± 4 e | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sutanto, K.D.; Husain, M.; Rasool, K.G.; Malik, A.F.; Al-Qahtani, W.H.; Aldawood, A.S. Persistency of Indigenous and Exotic Entomopathogenic Fungi Isolates under Ultraviolet B (UV-B) Irradiation to Enhance Field Application Efficacy and Obtain Sustainable Control of the Red Palm Weevil. Insects 2022, 13, 103. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010103
Sutanto KD, Husain M, Rasool KG, Malik AF, Al-Qahtani WH, Aldawood AS. Persistency of Indigenous and Exotic Entomopathogenic Fungi Isolates under Ultraviolet B (UV-B) Irradiation to Enhance Field Application Efficacy and Obtain Sustainable Control of the Red Palm Weevil. Insects. 2022; 13(1):103. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010103
Chicago/Turabian StyleSutanto, Koko Dwi, Mureed Husain, Khawaja Ghulam Rasool, Akhmad Faisal Malik, Wahidah Hazza Al-Qahtani, and Abdulrahman Saad Aldawood. 2022. "Persistency of Indigenous and Exotic Entomopathogenic Fungi Isolates under Ultraviolet B (UV-B) Irradiation to Enhance Field Application Efficacy and Obtain Sustainable Control of the Red Palm Weevil" Insects 13, no. 1: 103. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010103