Isolation of Beauveria Strains and Their Potential as Control Agents for Lema bilineata Germar (Coleoptera: Chrysomelidae)

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of Entomopathogenic Fungi
2.1.1. Molecular Identification
2.1.2. Phylogenetic Analysis
2.2. Rearing of Lema bilineata (Coleoptera: Chrysomelidae)
2.3. Pathogenicity Assays
2.3.1. Inoculum Preparation
2.3.2. Bioassay: Effects of Fungal Isolates on Adults of Lema bilineata (Coleoptera: Chrysomelidae)
2.3.3. Bioassay: Effects of Fungal Isolates on the Viability of Lema bilineata (Coleoptera: Chrysomelidae) Eggs
2.4. Statistical Analysis
3. Results and Discussion
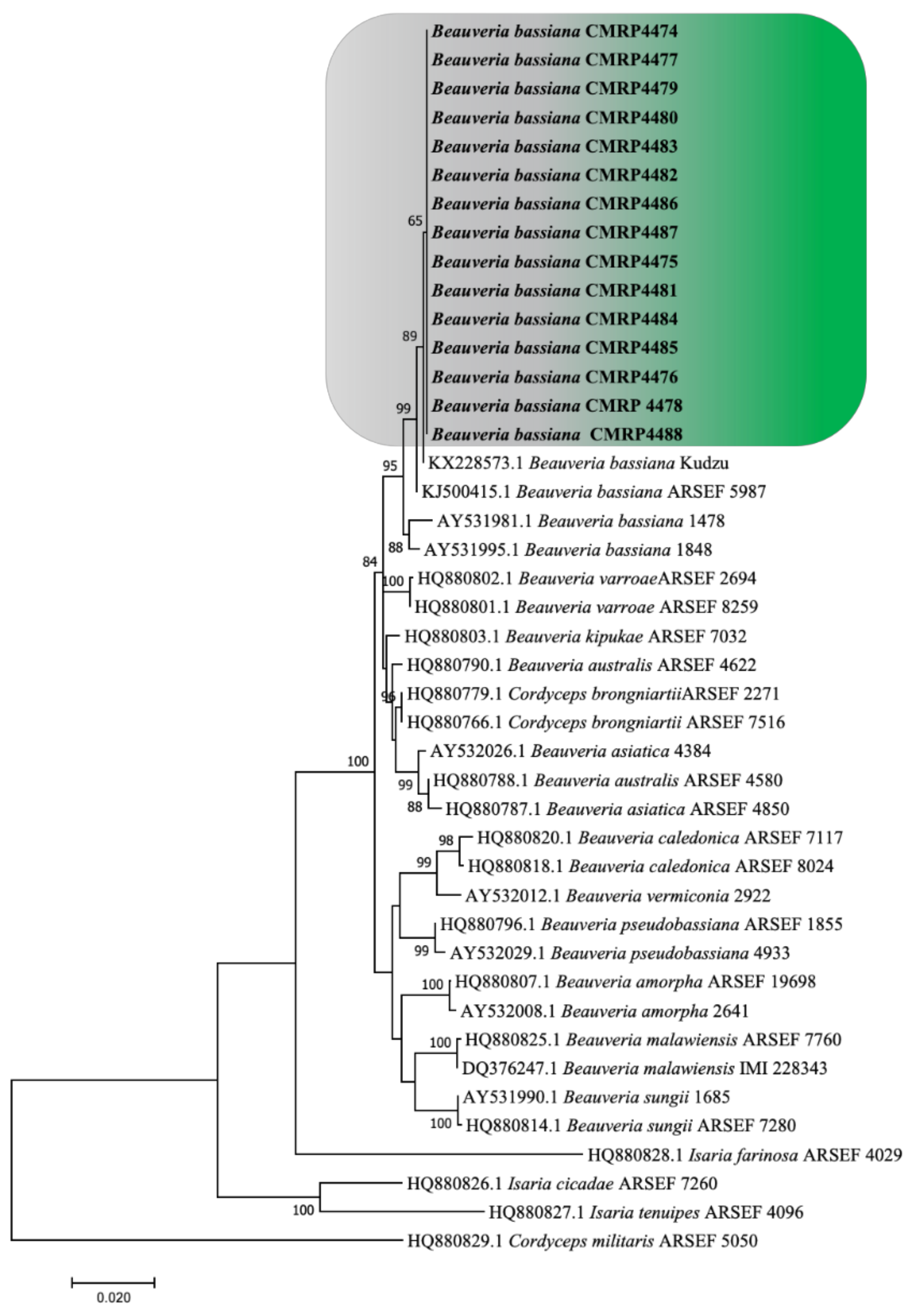
3.1. Isolation and Identification of Entomopathogenic Fungi
3.2. Pathogenicity Tests against Lema bilineata
3.2.1. Pathogenicity Test in Lema bilineata Adults
3.2.2. Pathogenicity Test in Lema bilineata Eggs
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Monti, M.M.; Ruocco, M.; Grobbelaar, E.; Pedata, A. Morphological and molecular characterization of Lema bilineata (Germar), a new alien invasive leaf beetle for Europe, with notes on the related species Lema daturaphila Kogan & Goeden. Insects 2020, 11, 295. [Google Scholar] [CrossRef]
- Bado, S.G.; Cerri, A.M.; Vilella, F.; Molina, A. Aspectos morfológicos, biológicos y de ingesta de Lema bilineata (Germ) (Coleoptera: Chrysomelidae) sobre Physalis peruviana L. Bol. San. Veg. Plagas 2000, 26, 5–10. [Google Scholar]
- Servizio Fitosanitario Regionale. Lema bilineata (Germar)—Chrysomelidae Sudamericano del Tabaco. Assesorato Agricoltura: Regione Campania, Italy. 2017. Available online: http://agricoltura.regione.campania.it/difesa/lema-bilineata.html (accessed on 21 May 2020).
- Bischoff, A.M.; Furuie, J.L.; Benatto, A.; Zimmermann, R.C.; Araujo, E.S.; Baena, R.; De Oliveira, M.C.H.; Zawadneak, M.A.C. First report of Lema bilineata Germar (Coleoptera: Chrysomelidae) damaging Physalis peruviana (L.) in Brazil. Ciência Rural 2021, 51, e20200735. [Google Scholar] [CrossRef]
- Stevens, M.M.; Stanton, R.A.; Wu, H.; Sampson, B.; Weir, T.A.; Reid, C.A.M.; Mo, J. Detection of Lema bilineata Germar (Coleoptera: Chrysomelidae) in Australia. Gen. Appl. Entomol. 2010, 39, 1–3. [Google Scholar]
- Ormeño, J.; Araya, J.E.; Díaz, C.A. Supervivencia y alimentación comparativa de larvas de Lema bilineata Germar en Datura spp., tabaco y otras solanáceas. Bol. San. Veg. Plagas 2002, 28, 21–31. [Google Scholar]
- Bennett, A.; Du Toit, C.L.N.; Bennett, A.L. A new record of Lema trilinea White (Coleoptera: Chrysomelidae) on tobacco in South Africa, with reference to the common pest species, Lema bilineata (Germar) (Coleoptera: Chrysomelidae, Criocerinae): Short Communication. Afr. Entomol. 1999, 7, 316–319. [Google Scholar]
- Brasil. Ministério da Agricultura, Pecuária e Abastecimento. Agrofit—Sistema de Agrotóxicos Fitossanitários. 2020. Available online: http://agrofit.agricultura.gov.br/agrofit_cons/principal_agrofit_cons (accessed on 21 February 2020).
- Baja, F.; Poitevin, C.G.; Araujo, E.S.; Mirás-Avalos, J.M.; Zawadneak, M.A.C.; Pimentel, I.C. Infection of Beauveria bassiana and Cordyceps javanica on different immature stages of Duponchelia fovealis Zeller (Lepidoptera: Crambidae). Crop Prot. 2020, 138, 105347. [Google Scholar] [CrossRef]
- Araujo, E.S.; Poltronieri, A.S.; Poitevin, C.G.; Mirás-Avalos, J.M.; Zawadneak, M.A.C.; Pimentel, I.C. Compatibility between entomopathogenic fungi and egg parasitoids (Trichogrammatidae): A laboratory study of their combined use to control Duponchelia fovealis. Insects 2020, 11, 630. [Google Scholar] [CrossRef] [PubMed]
- Araujo, E.S.; Benatto, A.; Rizzato, F.B.; Poltronieri, A.S.; Poitevin, C.G.; Zawadneak, M.A.C.; Pimentel, I.C. Combining biocontrol agents with different mechanisms of action to control Duponchelia fovealis, an invasive pest in South America. Crop Prot. 2020, 134, 105184. [Google Scholar] [CrossRef]
- Amatuzzi, R.F.; Poitevin, C.G.; Poltronieri, A.S.; Zawadneak, M.A.C.; Pimentel, I.C. Susceptibility of Duponchelia fovealis Zeller (Lepidoptera: Crambidae) to soil-borne entomopathogenic fungi. Insects 2018, 9, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poitevin, C.G.; Porsani, M.V.; Poltronieri, A.S.; Zawadneak, M.A.C.; Pimentel, I.C. Fungi isolated from insects in strawberry crops act as potential biological control agents of Duponchelia fovealis (Lepidoptera: Crambidae). Appl. Entomol. Zool. 2018, 53, 323–331. [Google Scholar] [CrossRef]
- Thomas, S.R.; Elkinton, J.S. Pathogenicity and virulence. J. Invertebr. Pathol. 2004, 85, 146–151. [Google Scholar] [CrossRef]
- De Hoog, G.S.; Guarro, J.; Gené, J.; Ahmed, S.; Al-Hatmi, A.M.S.; Figueras, M.J.; Vitale, R.G. Atlas of Clinical Fungi, 4th ed.; Utrecht, Universitat Rovira i Virgilli: Hilversum, The Netherlands, 2020; Available online: https://clinicalfungi.org/ (accessed on 21 January 2021).
- Vicente, V.A.; Attili-Agelis, D.; Pie, M.R.; Queiroz-Telles, F.; Cruz, M.; Najafzadeh, M.J.; De Hoog, G.S.; Zhao, J.; Pizzirani-Kleiner, A. Environmental isolation of black yeast-like fungi involved in human infection. Stud. Mycol. 2008, 61, 137–144. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Toh, H. Recent developments in the MAFFT multiple sequence alignment program. Brief Bioinform. 2008, 9, 286–298. [Google Scholar] [CrossRef] [Green Version]
- Rehner, S.A.; Minnis, A.M.; Sung, G.; Luangsa-ard, J.; Devotto, S.; Humber, R.A. Phylogeny and systematics of the anamorphic, entomopathogenic genus Beauveria. Mycologia 2011, 103, 1055–1073. [Google Scholar] [CrossRef] [PubMed]
- Alves, S.B.; Pereira, R.M. Produção de fungos entomopatogênicos. In Controle Microbiano de Insetos, 2nd ed.; Alves, S.B., Ed.; FEALQ: Piracicaba, Brazil, 1998; pp. 845–869. [Google Scholar]
- Abbott, W.S.A. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 58, 265–267. [Google Scholar] [CrossRef]
- Pell, J.K.; Hannam, J.J.; Steinkraus, D.C. Conservation biological control using fungal entomopathogens. BioControl 2010, 55, 187–198. [Google Scholar] [CrossRef]
- Altinok, H.H.; Altinok, M.A.; Koca, A.S. Modes of action of entomopathogenic fungi. Curr. Trends Nat. Sci. 2019, 8, 117–124. [Google Scholar]



{kind=link}
{kind=link}
| Isolate | Identification | GENBANK Accesion Number | ||
|---|---|---|---|---|
| Morphological | Molecular | ITS | ef-1α | |
| CMRP4474/L1 | Beauveria sp. | B. bassiana | MZ567032 | |
| CMRP4475/L2 | MZ567033 | |||
| CMRP4476/L3 | MZ567034 | |||
| CMRP4477/L4 | MZ567035 | |||
| CMRP4478/L5 | MZ567036 | |||
| CMRP4479/L6 | MZ567037 | |||
| CMRP4480/L7 | MZ567038 | MZ574443 | ||
| CMRP4481/L8 | MZ567039 | |||
| CMRP4482/L9 | MZ567040 | |||
| CMRP4483/L10 | MZ567041 | |||
| CMRP4484/L11 | MZ567042 | |||
| CMRP4485/L12 | MZ567043 | |||
| CMRP4486/L13 | MZ567044 | |||
| CMRP4487/L14 | MZ567045 | MZ574442 | ||
| CMRP4488/L15 | MZ567046 | |||
| Isolate | Mortality | |
|---|---|---|
| Total Number of Dead Lema bilineata Adults | Rate (%) | |
| Control | 3 | 0 a |
| CMRP4474 | 4 | 20 a |
| CMRP4475 | 8 | 60 a |
| CMRP4476 | 4 | 40 a |
| CMRP4477 | 6 | 40 a |
| CMRP4478 | 6 | 20 a |
| CMRP4479 | 6 | 60 a |
| CMRP4480 | 10 | 80 b |
| CMRP4481 | 8 | 60 a |
| CMRP4482 | 6 | 40 a |
| CMRP4483 | 8 | 60 a |
| CMRP4484 | 4 | 20 a |
| CMRP4485 | 4 | 40 a |
| CMRP4486 | 6 | 60 a |
| CMRP4487 | 10 | 80 b |
| CMRP4488 | 10 | 80 b |
| Isolate | Rate (%) |
|---|---|
| Control | 6.7 ± 1.8 a |
| CMRP4480 | 66.7 ± 1.6 c |
| CMRP4487 | 60.0 ± 2.0 b |
| CMRP4488 | 58.7 ± 2.4 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Furuie, J.L.; Stuart, A.K.d.C.; Voidaleski, M.F.; Zawadneak, M.A.C.; Pimentel, I.C. Isolation of Beauveria Strains and Their Potential as Control Agents for Lema bilineata Germar (Coleoptera: Chrysomelidae). Insects 2022, 13, 93. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010093
Furuie JL, Stuart AKdC, Voidaleski MF, Zawadneak MAC, Pimentel IC. Isolation of Beauveria Strains and Their Potential as Control Agents for Lema bilineata Germar (Coleoptera: Chrysomelidae). Insects. 2022; 13(1):93. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010093
Chicago/Turabian StyleFuruie, Jason Lee, Andressa Katiski da Costa Stuart, Morgana Ferreira Voidaleski, Maria Aparecida Cassilha Zawadneak, and Ida Chapaval Pimentel. 2022. "Isolation of Beauveria Strains and Their Potential as Control Agents for Lema bilineata Germar (Coleoptera: Chrysomelidae)" Insects 13, no. 1: 93. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010093