
Cryoprotective Response as Part of the Adaptive Strategy of the Red Palm Weevil, Rhynchophorus ferrugineus, against Low Temperatures

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Sampling
2.2. Measurement of Free Glucose, Trehalose and Glycerol in Hemolymph
2.3. Measurement of Free Amino Acids, Potassium and Sodium in Hemolymph
2.4. Statistical Analysis
3. Results
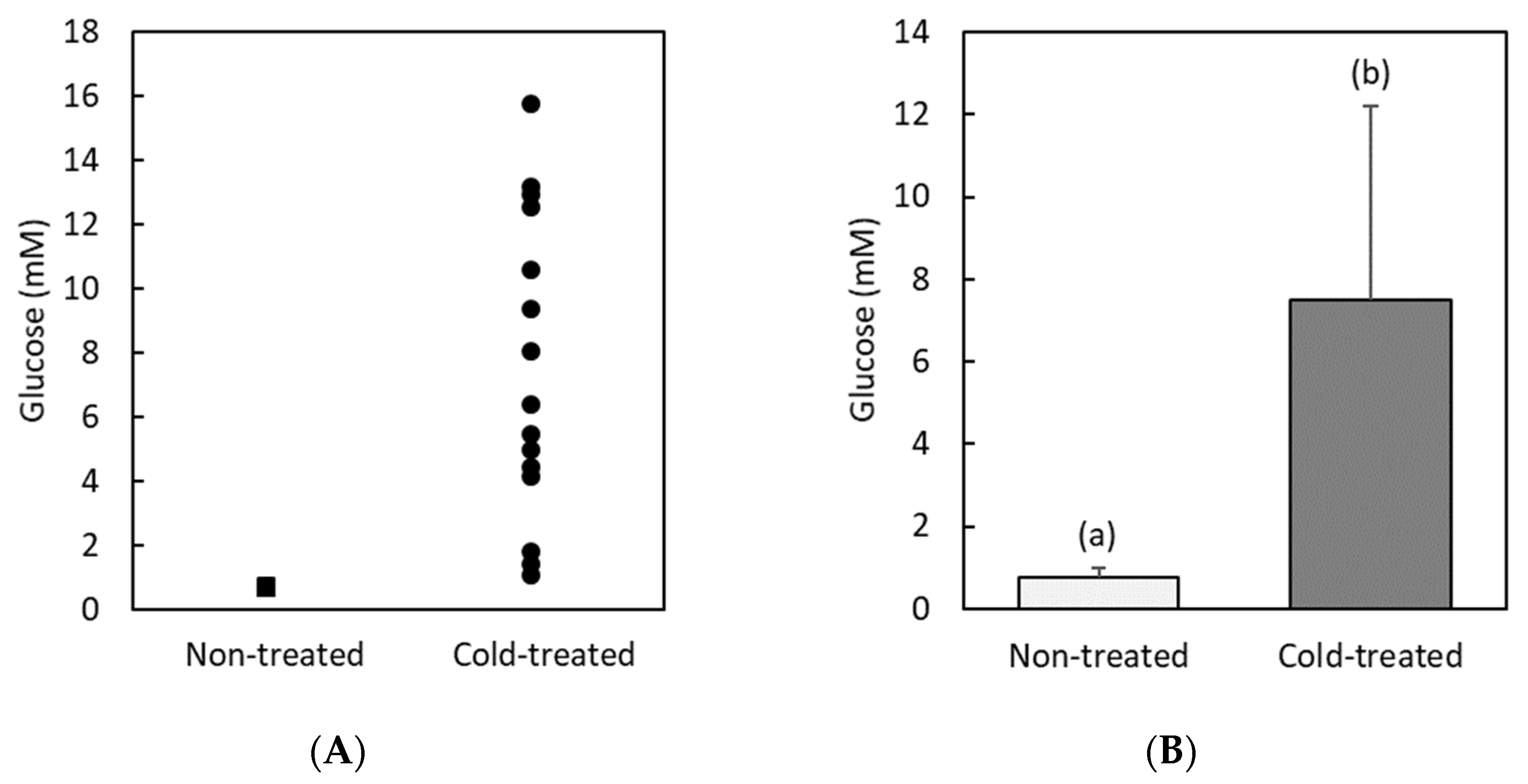
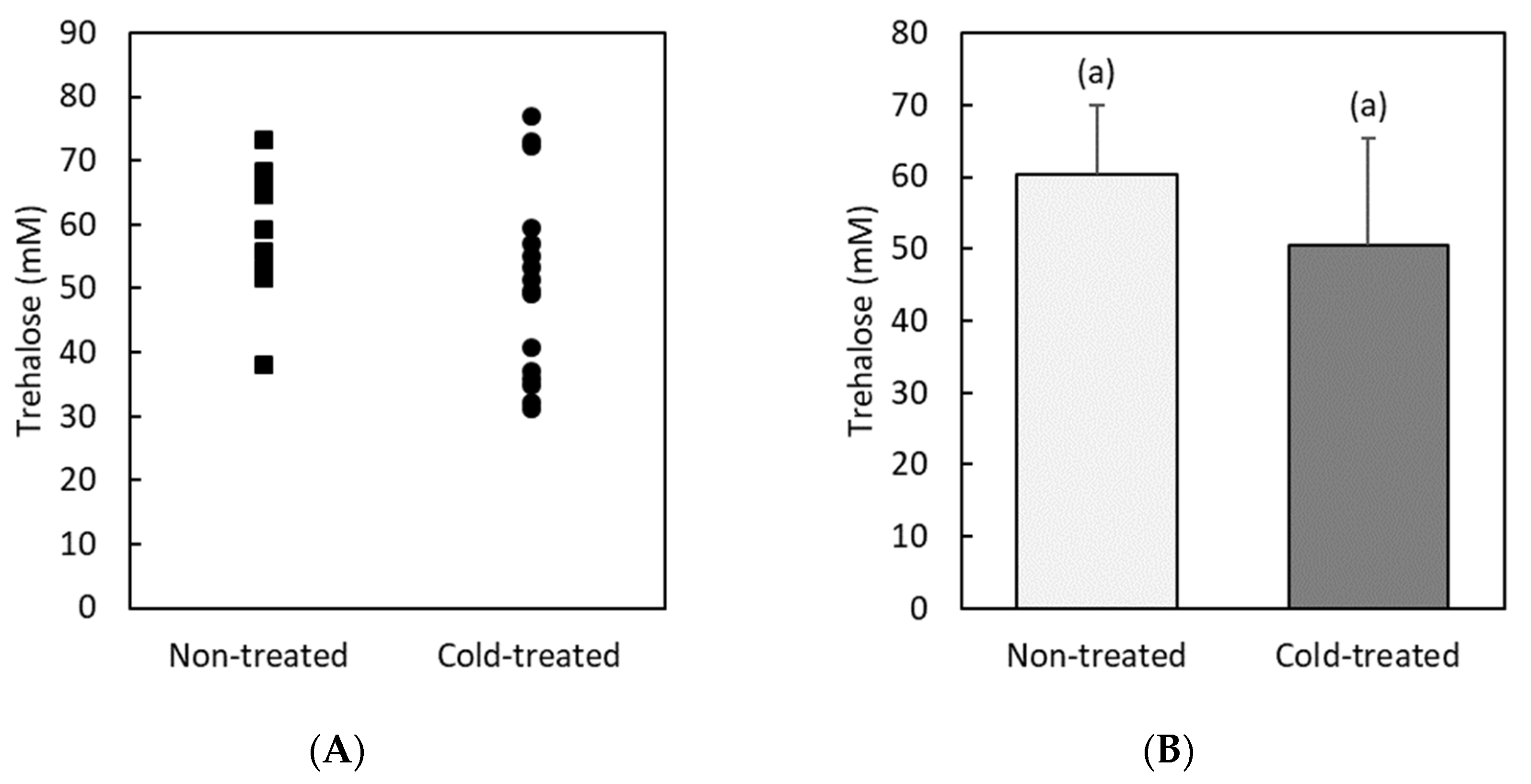
3.1. Glucose and Trehalose
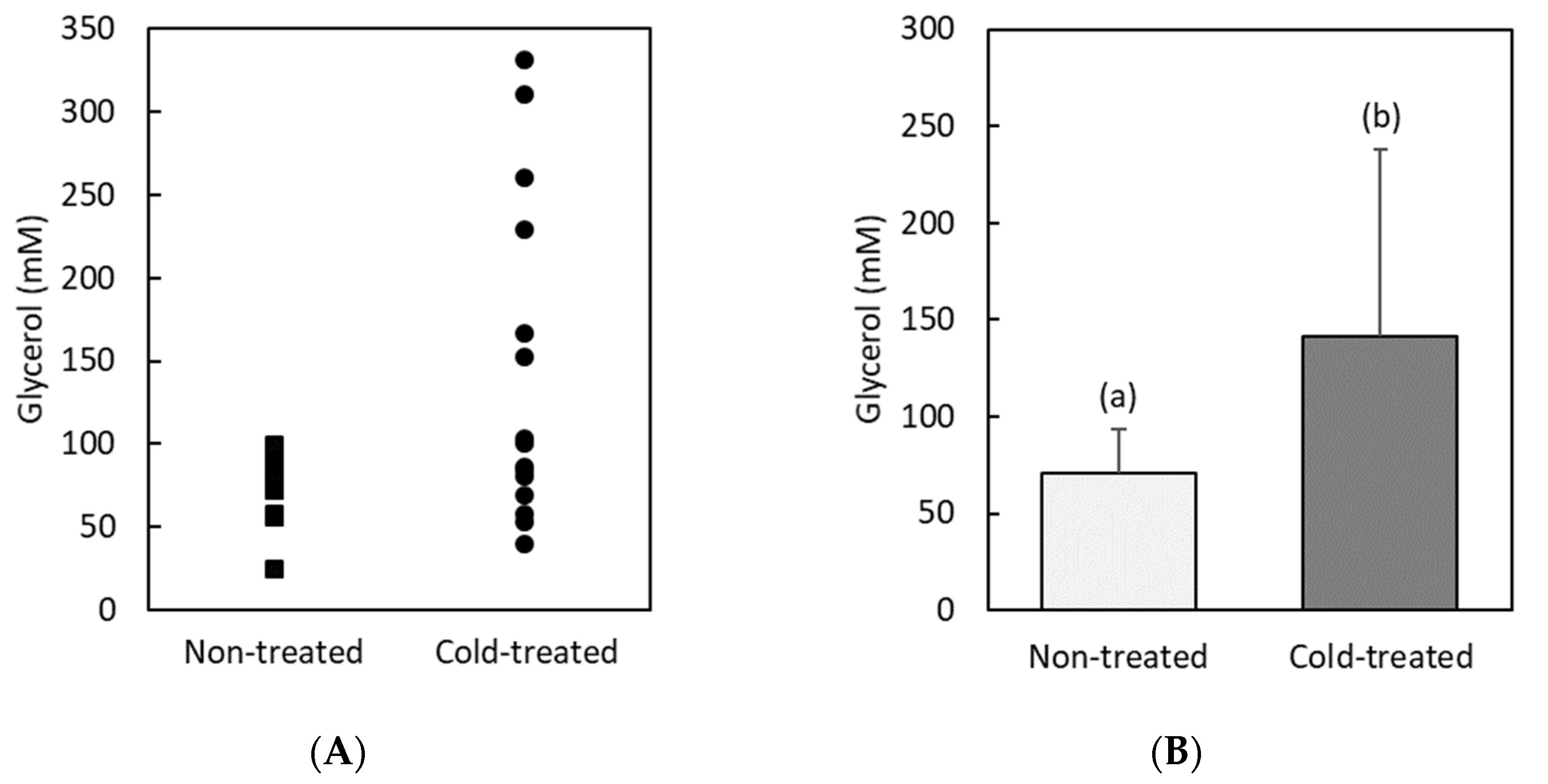
3.2. Glycerol
3.3. Free Amino Acids
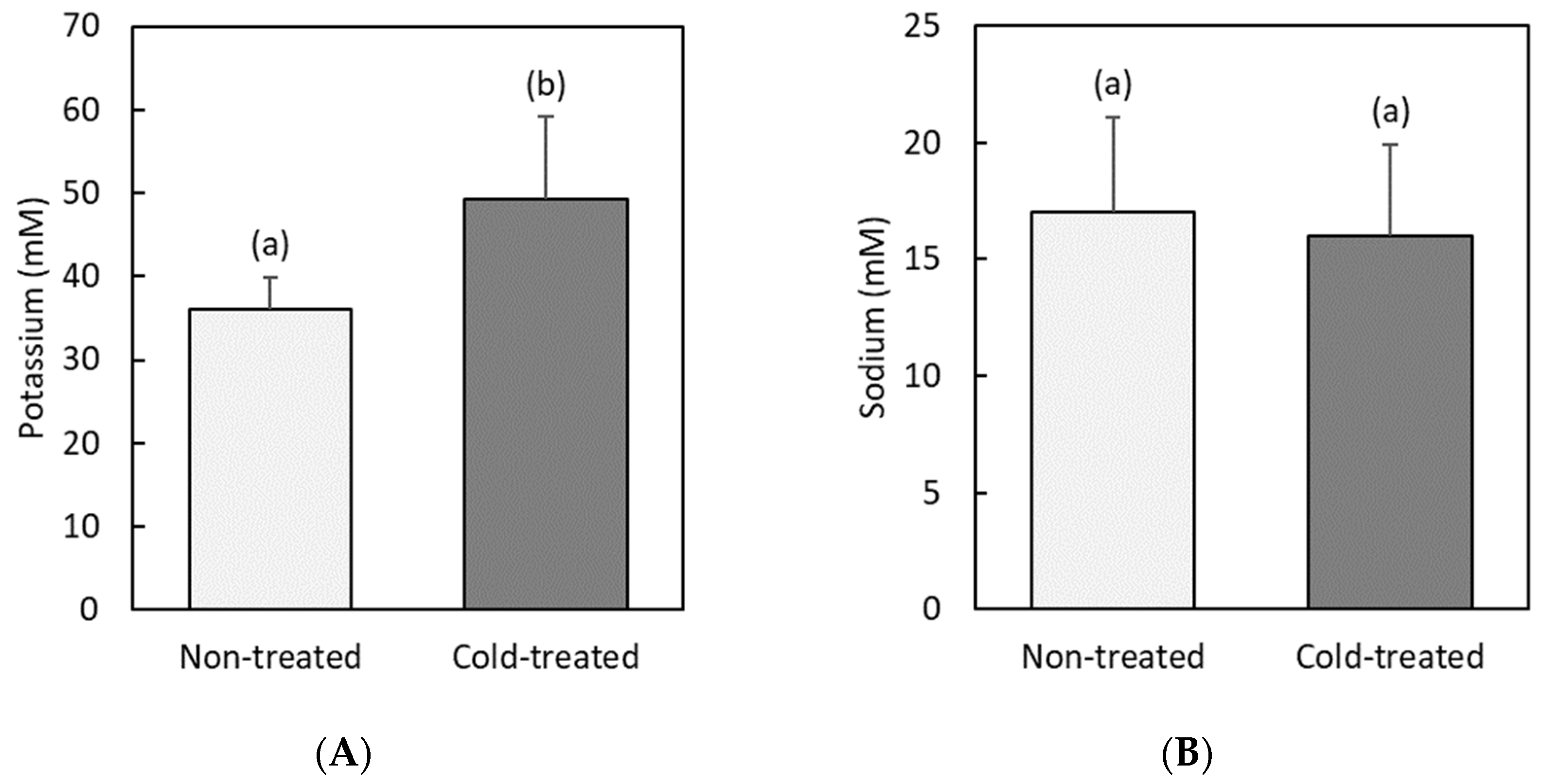
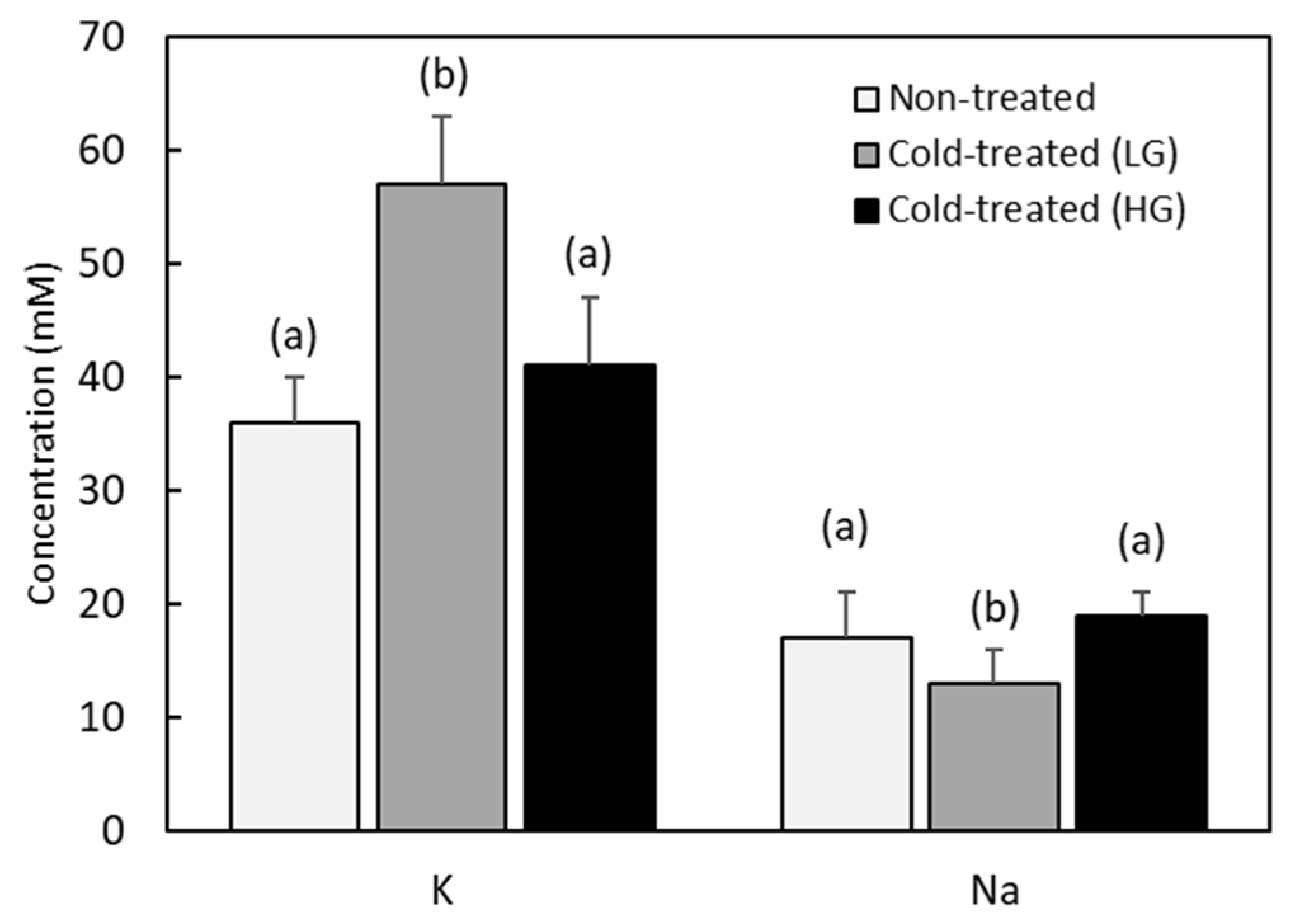
3.4. Potassium and Sodium
4. Discussion
5. Summary and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rugman-Jones, P.F.; Hoddle, C.D.; Hoddle, M.S.; Stouthamer, R. The Lesser of Two Weevils: Molecular-Genetics of Pest Palm Weevil Populations Confirm Rhynchophorus vulneratus (Panzer 1798) as a Valid Species Distinct from R. ferrugineus (Olivier 1790), and Reveal the Global Extent of Both. PLoS ONE 2013, 8, e78379. [Google Scholar] [CrossRef] [PubMed]
- FAO-CIHEAM. Current situation of red palm weevil in the NENA region. In Proceedings of the Scientific Consultation and High-Level Meeting on Red Palm Weevil Management, Rome, Italy, 29–31 March 2017; p. 25. [Google Scholar]
- Dembilio, O.; Jacas, J.A. Basic bio-ecological parameters of the invasive Red Palm Weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae), in Phoenix canariensis under Mediterranean climate. Bull. Entomol. Res. 2011, 101, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Milosavljević, I.; El-Shafie, H.A.F.; Faleiro, J.R.; Hoddle, C.D.; Lewis, M.; Hoddle, M.S. Palmageddon: The wasting of ornamental palms by invasive palm weevils, Rhynchophorus spp. J. Pest Sci. 2019, 92, 143–156. [Google Scholar] [CrossRef]
- Vacas, S.; Primo, J.; Navarro-Llopis, V. Advances in the use of trapping systems for Rhynchophorus ferrugineus (Coleoptera: Curculionidae): Traps and attractants. J. Econ. Entom. 2013, 106, 1739–1746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro-Llopis, V.; Primo, J.; Vacas, S. Improvements in Rhynchophorus ferrugineus (Coleoptera: Dryophthoridae) trapping systems. J. Econ. Entomol. 2018, 111, 1298–1305. [Google Scholar] [CrossRef] [PubMed]
- Dembilio, Ó.; Llácer, E.; Martínez de Altube, M.D.M.; Jacas, J.A. Field efficacy of imidacloprid and Steinernema carpocapsae in a chitosan formulation against the red palm weevil Rhynchophorus ferrugineus (Coleoptera: Curculionidae) in Phoenix canariensis. Pest. Manag. Sci. 2010, 66, 365–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gindin, G.; Levski, S.; Glazer, I.; Soroker, V. Evaluation of the entomopathogenic fungi Metarhizium anisopilae and Beauveria bassiana against the red palm weevil Rhynchophorus ferrugineus. Phytoparasitica 2006, 34, 370–379. [Google Scholar] [CrossRef]
- Solano-Rojas, Y.; Gámez, M.; López, I.; Garay, J.; Varga, Z.; Cabello, T. Conservation Strategy for Palm Groves: Optimal Chemical Control Model for Red Palm Weevil, Rhynchophorus ferrugineus. Agronomy 2021, 11, 920. [Google Scholar] [CrossRef]
- Fiaboe, K.K.M.; Peterson, A.T.; Kairo, M.T.K.; Roda, A.L. Predicting the Potential Worldwide Distribution of the Red Palm Weevil Rhynchophorus ferrugineus (Olivier) (Coleoptera: Curculionidae) using Ecological Niche Modeling. Fla. Entomol. 2012, 95, 659–673. [Google Scholar] [CrossRef]
- Ge, X.; He, S.; Wang, T.; Yan, W.; Zong, S. Potential Distribution Predicted for Rhynchophorus ferrugineus in China under Different Climate Warming Scenarios. PLoS ONE 2015, 10, e0141111. [Google Scholar] [CrossRef]
- Addo-Bediako, A.; Chown, S.L.; Gaston, K.J. Thermal tolerance, climatic variability and latitude. Proc. R. Soc. Lond. B 2000, 267, 739–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrew, N.R.; Hill, S.J.; Binns, M.; Bahar, M.H.; Ridley, E.V.; Jung, M.P.; Fyfe, C.; Yates, M.; Khusro, M. Assessing insect responses to climate change: What are we testing for? Where should we beheading? PeerJ 2013, 1, e11. [Google Scholar] [CrossRef] [PubMed]
- Harrison, J.F.; Woods, H.A.; Roberts, S.P. Ecological and Environmental Physiology of Insects; Oxford University Press: Oxford, UK, 2013; pp. 64–101. [Google Scholar]
- Kellermann, V.; Van Heerwaarden, B. Terrestrial insects and climate change: Adaptive responses in key traits. Phys. Entomol. 2019, 44, 99–115. [Google Scholar] [CrossRef] [Green Version]
- Lenoir, J.; Bertrand, R.; Comte, L.; Bourgeaud, L.; Hattab, T.; Murienne, J.; Grenouillet, G. Species better track climate warming in the oceans than on land. Nat. Ecol. Evol. 2020, 4, 1044–1059. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid Range Shifts of Species Associated with High Levels of Climate Warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.-C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B.; et al. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, eaai9214. [Google Scholar] [CrossRef]
- Rind, D. The dynamics of warm and cold climates. J. Atmos. Sci. 1986, 43, 3–43. [Google Scholar] [CrossRef] [Green Version]
- Lencioni, V. Survival strategies of freshwater insects in cold environments. J. Limnol. 2004, 63, 45–55. [Google Scholar] [CrossRef]
- Denlinger, D.L.; Lee, R.E. Low Temperature Biology of Insects; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Koštál, V.; Korbelová, J.; Štĕtina, T.; Poupardin, R.; Colinet, H.; Zahradníčková, H.; Opekarová, I.; Moos, M.; Šimek, P. Physiological basis for low temperature survival and storage of quiescent larvae of the fruit fly Drosophila melanogaster. Sci. Rep. 2016, 6, 32346. [Google Scholar] [CrossRef]
- Cubillos, C.; Cáceres, J.C.; Villablanca, C.; Villarreal, P.; Baeza, M.; Cabrera, R.; Graether, P.; Veloso, C. Cold tolerance mechanisms of two arthropods from the andean range of Central Chile: Agathemera crassa (insecta: Agathemeridae) and Euathlus condorito (arachnida: Theraphosidae). J. Therm. Biol. 2018, 74, 133–139. [Google Scholar] [CrossRef]
- Toxopeus, J.; Sinclair, B. Mechanisms underlying insect freeze tolerance. Biol. Rev. 2018, 93, 1891–1914. [Google Scholar] [CrossRef] [PubMed]
- Saeidi, M.; Moharramipour, S. Physiology of cold hardiness, seasonal fluctuations, and cryoprotectant contents in overwintering adults of Hypera postica (Coleoptera: Curculionidae). Environ. Entomol. 2017, 46, 960–966. [Google Scholar] [CrossRef] [PubMed]
- Storey, K.B.; Storey, J.M. Freeze tolerance in animals. Physiol. Rev. 1988, 68, 27–84. [Google Scholar] [CrossRef] [PubMed]
- Toxopeus, J.; Koštál, V.; Sinclair, B.J. Evidence for non-colligative function of small cryoprotectants in a freeze-tolerant insect. Proc. R. Soc. B 2019, 286, 20190050. [Google Scholar] [CrossRef] [Green Version]
- Olsson, T.; MacMillan, H.A.; Nyberg, N.; Staerk, D.; Malmendal, A.; Overgaard, J. Hemolymph metabolites and osmolality are tightly linked to cold tolerance of Drosophila species: A comparative study. J. Exp. Biol. 2016, 219, 2504–2513. [Google Scholar] [CrossRef] [Green Version]
- Sømme, L. Effects of glycerol on cold-hardiness in insects. Can. J. Zool. 2011, 42, 87–101. [Google Scholar] [CrossRef]
- Yoder, J.A.; Benoit, J.B.; Denlinger, D.L.; Rivers, D.B. Stress induced accumulation of glycerol in the flesh fly, Sarcophaga bullata: Evidence indicating anti-desiccant and cryoprotectant functions of this polyol and a role for the brain in coordinating the response. J. Insect Physiol. 2006, 52, 202–214. [Google Scholar] [CrossRef]
- Wang, H.; Lei, Z.; Li, X.; Oetting, R.D. Rapid cold hardening and expression of heat shock protein genes in the B-biotype Bemisia tabaci. Environ. Entomol. 2011, 40, 132–139. [Google Scholar] [CrossRef]
- Overgaard, J.; Sorensen, J.G.; Com, E.; Colinet, H. The rapid cold hardening response of Drosophila melanogaster: Complex regulation across different levels of biological organization. J. Insect Physiol. 2014, 62, 46–53. [Google Scholar] [CrossRef]
- Martin, M.M.; Cabello, T. Biología y Ecología del Curculiónido Rojo de la Palmera, Rhynchophorus ferrugineus; Universidad de Almería: Almería, Spain, 2005; pp. 1–202. ISBN 84-689-5292-3. [Google Scholar]
- Martin, M.M.; Cabello, T. Manejo de la cría del picudo rojo de la palmera, Rhynchophorus ferrugineus (Olivier, 1790) (Coleoptera, Dryophthoridae), en dieta artificial y efectos en su biometría y biología. Boletín Sanid. Veg. Plagas 2006, 32, 631–641. [Google Scholar]
- León-Quinto, T.; Fimia, A.; Madrigal, R.; Serna, A. Morphological response of the red palm weevil, Rhynchophorus ferrugineus, to a transient low temperature analyzed by computer tomography and holographic microscopy. J. Therm. Biol. 2020, 94, 102748. [Google Scholar] [CrossRef] [PubMed]
- Koštál, V.; Renaultc, D.; Mehrabianová, A.; Bastl, J. Insect cold tolerance and repair of chill-injury at fluctuating thermal regimes: Role of ion homeostasis. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 147, 231–238. [Google Scholar] [CrossRef] [PubMed]
- MacMillan, H.A.; Baatrup, E.; Overgaard, J. Concurrent effects of cold and hyperkalaemia cause insect chilling injury. Proc. R. Soc. B 2015, 282, 20151483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrington, J.; Andersen, M.A.; Brzezinski, K.; MacMillan, H.A. Hyperkalaemia, not apoptosis, accurately predicts insect chilling injury. Proc. R. Soc. B 2020, 287, 20201663. [Google Scholar] [CrossRef] [PubMed]
- MacMillan, H.A.; Andersen, J.L.; Davies, S.A.; Overgaard, J. The capacity to maintain ion and water homeostasis underlies interspecific variation in Drosophila cold tolerance. Sci. Rep. 2015, 5, 18607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, M.S.; Worland, M.R. How insects survive the cold: Molecular mechanisms—A review. J. Comp. Physiol. B 2008, 178, 917–933. [Google Scholar] [CrossRef] [Green Version]
- Teets, N.M.; Denlinger, D.L. Physiological mechanisms of seasonal and rapid cold-hardening in insects. Phys. Ent. 2013, 38, 105–116. [Google Scholar] [CrossRef]
- Michaud, M.R.; Denlinger, D.L. Shifts in the carbohydrate, polyol, and amino acid pools during rapid cold-hardening and diapause-associated cold-hardening in flesh flies (Sarcophaga crassipalpis): A metabolomic comparison. J. Comp. Physiol. B 2007, 177, 753–763. [Google Scholar] [CrossRef]
- Lalouette, L.; Koštál, V.; Colinet, H.; Gagneul, D.; Renault, D. Cold exposure and associated metabolic changes in adult tropical beetles exposed to fluctuating thermal regimes. FEBS J. 2007, 274, 1759–1767. [Google Scholar] [CrossRef]
- Li, N.G. Cryoprotectant systems and cold tolerance of insects inhabiting central Yakutia (Russian Far East). Eur. J. Èntomol. 2016, 113, 537–541. [Google Scholar] [CrossRef] [Green Version]
- Chowanski, S.; Lubawy, J.; Spochacz, M.; Paluch, E.; Smykalla, G.; Rosinski, G.; Slocinska, M. Cold induced changes in lipid, protein and carbohydrate levels in the tropical insect Gromphadorhina coquereliana. Comp. Bioch. Phys. A 2015, 183, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.H.; Qi, X.L.; Kang, L. Geographic differences on accumulation of sugars and polyols in locust eggs in response to cold acclimation. J. Insect Physiol. 2010, 56, 966–970. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Xu, L.; Li, W.; Xu, Z.; Cao, M.; Wang, J.; Tao, J.; Xong, S. Seasonal changes in supercooling capacity and major cryoprotectants of overwintering Asian longhorned beetle (Anoplophora glabripennis) larvae. Agric. For. Entomol. 2016, 18, 302–312. [Google Scholar] [CrossRef]
- Izumi, Y.; Sonoda, S.; Yoshida, H.; Danks, H.V.; Tsumuki, H. Role of membrane transport of water and glycerol in the freeze tolerance of the rice stem borer, Chilo suppressalis Walker (Lepidoptera: Pyralidae). J. Insect Physiol. 2006, 52, 215–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fields, P.G.; Fleurat-Lessard, F.; Lavenseau, L.; Febvay, G.; Peypelut, L.; Bonnot, G. The effect of cold acclimation and deacclimation on cold tolerance, trehalose and free amino acid levels in Sitophilus granarius and Cryptolestes ferrugineus (Coleoptera). J. Insect Phys. 1998, 44, 955–965. [Google Scholar] [CrossRef]
- Goto, M.; Li, Y.-P.; Kayaba, S.; Outani, S.; Suzuki, K. Cold hardiness in summer and winter diapause and post-diapause pupae of the cabbage armyworm, Mamestra brassicae L. under temperature acclimation. J. Insect Phys. 2001, 47, 709–714. [Google Scholar] [CrossRef]
- Rivers, D.B.; Lee, R.E., Jr.; Denlinger, D.L. Cold hardiness of the fly pupal parasitoid Nasonia vitripennis is enhanced by its host, Sarcophaga crassipalpis. J. Insect Phys. 2000, 46, 99–106. [Google Scholar] [CrossRef]
- Carpenter, J.F.; Crowe, J.H. The mechanism of cryoprotection of proteins by solutes. Cryobiology 1988, 25, 244–255. [Google Scholar] [CrossRef]
- Michaud, M.R.; Benoit, J.B.; Lopez-Martinez, G.; Elnitsky, M.A.; Lee, R.E., Jr.; Denlinger, D.L. Metabolomics reveals unique and shared metabolic changes in response to heat shock, freezing and desiccation in the Antarctic midge, Belgica Antarctica. J. Insect Phys. 2008, 54, 645–655. [Google Scholar] [CrossRef]
- Goto, M.; Sekine, Y.; Outa, H.; Hujikura, M.; Suzuki, K. Relationships between cold hardiness and diapause, and between glycerol and free amino acid contents in overwintering larvae of the oriental corn borer, Ostrinia furnacalis. J. Insect Phys. 2001, 47, 157–165. [Google Scholar] [CrossRef]
- Uzelac, I.; Avramov, M.; Čelić, T.; Vukašinović, E.; Gošić-Dondo, S.; Purać, J.; Kojić, D.; Blagojević, D.; Popović, Z.D. Effect of Cold Acclimation on Selected Metabolic Enzymes During Diapause in The European Corn Borer Ostrinia nubilalis (Hbn.). Sci. Rep. 2020, 10, 9085. [Google Scholar] [CrossRef] [PubMed]
- Coello Alvarado, L.E.C.; MacMillan, H.A.; Sinclair, B.J. Chill-tolerant Gryllus crickets maintain ion balance at low temperatures. J. Insect Physiol. 2015, 77, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Lebenzon, J.E.; Des Marteaux, L.E.; Sinclair, B.J. Reversing sodium differentials between the hemolymph and hindgut speeds chill coma recovery but reduces survival in the fall field cricket, Gryllus pennsylvanicus. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2020, 244, 110699. [Google Scholar] [CrossRef] [PubMed]
- MacMillan, H.A.; Andersen, J.L.; Loeschcke, V.; Overgaard, J. Sodium distribution predicts the chill tolerance of Drosophila melanogaster raised in different thermal conditions. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 308, 823–831. [Google Scholar] [CrossRef] [Green Version]
- Ju, R.T.; Wang, F.; Xiao, Y.Y.; Li, Y.Z.; Du, Y.Z. Cold hardiness of the red palm weevil, Rhyncophorus ferrugineuss (Olivier) (Coleoptera: Curculionidae) in Shanghai. Acta Entomol. Sinica 2010, 53, 226–232. [Google Scholar]
- Abe, F.; Ohkusu, M.; Kubo, T.; Kawamoto, S.; Sone, K.; Hata, K. Isolation of yeasts from palm tissues damaged by the red palm weevil and their possible effect on the weevil overwintering. Mycoscience 2010, 51, 215–223. [Google Scholar] [CrossRef]
- Khalil, S.R.; Awad, A.; Mohammed, H.H.; Nassan, M.A. Imidacloprid insecticide exposure induces stress and disrupts glucose homeostasis in male rats. Environ. Toxicol Pharmacol. 2017, 55, 165–174. [Google Scholar] [CrossRef]
- Sawczyn, T.; Dolezych, B.; Klosok, M.; Augustyniak, M.; Stygar, D.; Buldak, R.J.; Kukla, M.; Michalczyk, K.; Karcz-Socha, I.; Zwirska-Korczala, K. Alteration of carbohydrates metabolism and midgut glucose absorption in Gromphadorhina portentosa after subchronic exposure to imidacloprid and fenitrothion. J. Environ. Sci. Health Part A 2012, 47, 1644–1651. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | Concentration (mg/mL) | ANOVA Values | |||
|---|---|---|---|---|---|
| Non-Treated | Cold-Treated | F | p | ||
| Aspartic acid + Asparagine | ASP | 1.3 ± 0.3 a | 1.8 ± 0.3 b | 8.0 | 0.01 |
| Glutamic acid + Glutamine | GLU | 2.6 ± 0.5 a | 2.3 ± 0.4 a | 1.2 | 0.3 |
| Alanine | ALA | 1.8 ± 0.6 a | 2.7 ± 0.9 b | 4.9 | 0.04 |
| Arginine | ARG | 0.7 ± 0.2 a | 0.8 ± 0.2 a | 1.7 | 0.2 |
| Cysteine | CYS | <0.2 | < 0.2 | ||
| Phenylalanine | PHE | 1.2 ± 0.3 a | 1.5 ± 0.2 a | 3.6 | 0.08 |
| Glycine | GLY | 1.6 ± 0.6 a | 1.6 ± 0.8 a | 0.01 | 0.9 |
| Histidine | HIS | 0.7 ± 0.2 a | 1.2 ± 0.2 b | 21.0 | 0.001 |
| Isoleucine | ISO | 1.1 ± 0.2 a | 1.5 ± 0.2 b | 13.2 | 0.003 |
| Leucine | LEU | 1.6 ± 0.3 a | 2.1 ± 0.4 b | 9.0 | 0.01 |
| Lysine | LYS | 2.1 ± 0.5 a | 2.6 ± 0.5 a | 3.6 | 0.08 |
| Methionine | MET | <0.2 | <0.2 | ||
| Proline | PRO | 7.1 ± 2.2 a | 6.2 ± 1.4 a | 0.9 | 0.4 |
| Serine | SER | 0.8 ± 0.3 a | 1.1 ± 0.2 b | 5.6 | 0.04 |
| Tyrosine | TYR | 0.6 ± 0.2 a | 0.9 ± 0.2 a | 4.5 | 0.05 |
| Threonine | THR | 0.7 ± 0.2 a | 1.0 ± 0.1 b | 19.7 | 0.001 |
| Tryptophan | TRY | <0.2 | <0.2 | ||
| Valine | VAL | 1.4 ± 0.2 a | 2.0 ± 0.2 b | 19.4 | 0.001 |
| TOTAL | TOT | 25 ± 3 a | 29 ± 4 a | 4.3 | 0.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
León-Quinto, T.; Serna, A. Cryoprotective Response as Part of the Adaptive Strategy of the Red Palm Weevil, Rhynchophorus ferrugineus, against Low Temperatures. Insects 2022, 13, 134. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020134
León-Quinto T, Serna A. Cryoprotective Response as Part of the Adaptive Strategy of the Red Palm Weevil, Rhynchophorus ferrugineus, against Low Temperatures. Insects. 2022; 13(2):134. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020134
Chicago/Turabian StyleLeón-Quinto, Trinidad, and Arturo Serna. 2022. "Cryoprotective Response as Part of the Adaptive Strategy of the Red Palm Weevil, Rhynchophorus ferrugineus, against Low Temperatures" Insects 13, no. 2: 134. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020134