1. Introduction
The acquisition of a primordial free-living prokaryote ancestor of mitochondria by early eukaryotes is probably the most important step in the evolution of complex life [
1]. The mitochondrial genome codes for only a few genes, but they are immensely important for the metabolism and the high energy efficiency of the eukaryotic cells. Products of the genes encoded by the mitochondrial genome act in conjunction with products of the nuclear genome. These complex interactions include cellular respiration as well as mtDNA replication, transcription and translation, all remarkably important biological processes [
2].
The mitochondrial electron transport chain (METC), which is the site of the oxidative phosphorylation pathway (OXPHOS), is tightly orchestrated by the epistasis of (the genes encoded over) the two genomes. Adenosine triphosphate (ATP) is a cell fuel that is produced in the OXPHOS, by five multi-subunit enzyme complexes, four of which are comprised by the subunits encoded by both genomes [
3]. METC function is dependent on the synchronised interaction between mtDNA-encoded proteins (and RNAs) and nuclear-encoded proteins that are imported in the mitochondria. These protein subunits originating from two different genomes require high compatibility analogous to a ‘lock and key’ principle to preserve their configuration and enzymatic activity. Any incompatibilities can compromise their structural and biochemical properties, which in turn can cause electron leakage and consequently oxidative stress.
Apart from the aforementioned direct influence on the OXPHOS, mito-nuclear interactions have an indirect impact on it by being enrolled in the processes of transcription and translation of mitochondrial subunits involved in METC, as well as replication of the mtDNA. Transcription of these mtDNA genes involved in the OXPHOS pathway is completely regulated by the nucleus. All polypeptides involved in the process are nuclear-coded and imported to the mitochondria. This process usually consists of two transcription factors and a single subunit mitochondrial RNA polymerase (POLRMT) which is liable for promoter-binding specificity and strength. This is the basis for transcriptional mito-nuclear interactions because the nuDNA-coded protein needs to be structurally complementary to the control region of the mitochondrial DNA, or regulation of mtDNA genes transcription is compromised [
4]. Experiments have shown that when combining factors of the mitochondrial transcription machinery from different taxa, the more distant the taxa, the more deficient the transcription [
5,
6,
7]. The authors suggest that this is due to coevolution between binding motifs in the POLRMT or splicing peptides and the mtDNA recognition sites in each species [
5]. Replication of the mtDNA is also reliant on the activity of the POLRMT, as it is responsible for RNA primer creation after which replication can begin using the mtDNA polymerase complex. This way, mito-nuclear co-adaptions are accountable for mtDNA replication as well [
8,
9]. Apart from coding for subunits involved in OXPHOS, mtDNA also encodes for transfer RNAs (tRNAs), which cooperate with nuclear-encoded mitochondrial aminoacyl-tRNA synthetases (mt-aaRSs) in the process of translating mtDNA proteins [
2,
10], giving rise to another level on which mito-nuclear interactions are recognized by selection.
Since mtDNA products play such a pivotal role in the eukaryotic metabolism and normal cell functioning, scientists have long thought that any variation in the mitochondrial genome that impacts fitness would quickly be either purged or fixed by natural selection [
11,
12], resulting in erosion of genetic variation. This view was encouraged by the attributes of the mitochondrial genome, specifically being haploid and its exclusively maternal inheritance. A further consequence of haploidy in the absence of recombinations and heterozygosis, in which recessive alleles would be masked by dominant ones, is that all alleles are exposed to selection. Scientists thought that the high mutation rates of mtDNA are the reason for the relatively high levels of functional mtDNA variation, but theoretical models have proven that the observed levels of standing genetic variation are much higher than expected under the mutation–selection balance [
13].
In the last few decades, this traditional paradigm of rejecting mitochondrial DNAs functional and evolutional relevance has shifted, with the accumulating evidence linking specific mtDNA sequence polymorphisms to substantial phenotypic effects [
14]. First, signs of positive selection acting on the mtDNA in a broad spectrum of phyla have been reported by comparing the distribution of synonymous and nonsynonymous substitutions within and between species [
11,
15,
16,
17,
18,
19]. Second, a correlation between environmental factors and the distribution of mtDNA haplotypes has been observed [
18,
20,
21]. Not long after, experimental evidence started to amass, pointing to fitness consequences ensued by mtDNA sequence variation [
22,
23,
24,
25].
The presence of adaptive variation in mtDNA between populations is quite easy to explain because distinct populations usually mean independent genetic pools from which the stochastic forces sieve different alleles, but also different selection pressures favouring distinct haplotypes. Even detrimental alleles can become specific for a particular population if they are compatible with arising compensatory mutations in the nuclear genome [
26,
27,
28,
29,
30,
31,
32]. Therefore, the joint mito-nuclear interactions mentioned above are important in that respect [
33].
However, the existence of stable adaptive sympatric variation is much more difficult to explain, given its haploid nature and the importance of mitochondrial genes. Over the years, many ambiguous results have been published on the subject of the role of mito-nuclear interactions in sustaining adaptive sympatric variation. While some authors [
25,
34] could not prove sympatric variance being maintained with mito-nuclear interactions, others proved it while using the same model species and similar experimental set-ups [
26,
35]. Current theoretical understanding suggests that conditions required for maintenance of adaptive within-population variance are restrictive [
36,
37,
38]. Experimental papers on the subject have been scarce throughout the years [
1,
26]. A recent growing body of theoretical and experimental work has started to shed light on the subject, especially in light of cytonuclear interactions.
Negative frequency-dependent selection (NFDS) was one of the first mechanisms of balancing selection proposed to be maintaining sympatric mitochondrial variation. When it comes to this type of balancing selection, an allele’s relative fitness is inversely proportional to its frequency in the population. In a nutshell, NFDS constitutes that rarer alleles are favoured by natural selection over more common ones. The role of negative-frequency dependent selection in maintaining genetic variation in the nuclear genome is well described in the example of chromosomal inversions in
Drosophila ananassae [
39] and maintaining self-incompatibility in plants [
40]. The first proposition of the NFDS’s role in maintaining mtDNA variation by acting on cytonuclear gene interactions was brought by Gregorius and Ross [
38], but it was not until recently that it was hypothesised and experimentally proven that NFDS is responsible for sustaining mtDNA polymorphism in laboratory populations of seed beetle [
41,
42]. Furthermore, experiments on
Drosophila subobscura are starting to give weight to NFDS effects on preserving mtDNA variation [
43,
44].
Sex-specific selection (SSS) is an additional balancing selection mechanism that was proposed to be maintaining standing sympatric mtDNA variation [
45,
46]. Theory predicts that some mito-nuclear haplotype combinations result in a higher relative fitness in one sex, while different combinations provide better fitness in the other sex. Since mitochondria are maternally transmitted, the selection can act on them only through females. Because of the different reproduction strategies and behavioural patterns and sex-specific life histories in general, males and females have different fitness, as selection only recognizes females, favouring mutations that have a positive impact on females’ fitness even though these mutations can have deleterious effects in males. This hypothesis is known as the mother’s curse [
47,
48]. Different mutations in mitochondria importing nuclear-encoded proteins that are favourable to males may arise, pulling corresponding mitochondria-coded proteins to compensatory adapt and adjust to the new biochemical and structural stability and to preserve the high compatibility and efficacy [
49]. The idea of SSS maintaining intra-population variation was reinforced in the next couple of years by many experiments on
Drosophila species [
50,
51,
52], confirming that SSS acting on these mito-nuclear interactions can maintain intra-population variation.
Rand [
35] developed a model of joint transmission of X chromosome and cytoplasm in
Drosophila melanogaster and showed that maintenance of sympatric mitochondrial variation can be supported if the nuclear component of the interaction is located on the X chromosome.
Another mechanism proposed is the idea that the mitochondrial or mito-nuclear variation is shaped by differences in environmental factors that vary temporally or within habitat [
22,
50]. In that respect, genotype-by-environment interactions are crucial in upholding sympatric mtDNA variation. Dowling et al. [
50] showed that multiple mito-nuclear haplotypes can be maintained by epistatic interactions between mitochondrial and nuclear genes in a random mating population. Their findings show that the adaptive value of specific mito-nuclear combinations are determined by the environment in which they are expressed. Furthermore, Willet et al. [
53] have shown the substantial influence of temperature and light regime on selection on interpopulation mito-nuclear crosses of intertidal copepod
Tigriopus californicus. In this study, it is advocated that differences in protein interactions at varying temperatures are the reason why specific mito-nuclear combinations are favoured at a given temperature range. Oscillating temperature settings over space and time could act to maintain not only mitochondrial but mito-nuclear variation within populations, when this mtDNA × nuDNA × environment epistasis occurs [
54].
Thus, different mitochondrial polymorphisms can be auspicious in different extrinsic terms, for example, temperature, or different intrinsic conditions, be it a different nuclear genetic background (if possible, X linked) or different sex. In addition, the mere frequency of the polymorphism can make it more or less favourable.
Insects have long served as suitable models to measure the adaptive significance of mitochondrial or mito-nuclear variation. Generally, their generation time is short, making it possible to fully replace the existing nuclear background with the desired one by multi-generational backcrossing in a relatively short time [
41,
52,
54,
55]. The experimental model used in this study,
Drosophila obscura, possesses a high level of sympatric mtDNA variation across the species range [
56]. Therefore, it gives an excellent possibility to test the adaptive significance of sympatric mtDNA variation, and consequently forces that maintain it. Our preliminary work on desiccation resistance, in this model species [
57], identified the importance of temperature-specific effects on mito-nuclear variation, and to a lesser extent, the effects of SSS. In the present study, we use four Experimental Blocks (EBs), each representing variation collected within a specific natural site. Each block has three distinct mtDNA haplotypes and three distinct nuclear genetic backgrounds that are combined in nine possible experimental mito-nuclear introgression lines (MNILs). These MNILs were subjected to measurements of a set of life-history traits (both larval and adult) at two different temperatures. This way, we disentangle the specific effect of mtDNA on fitness and its dependence on nuclear genetic background, experimental temperature and sex of the individual. Ultimately, we discuss the importance of mito-nuclear interactions, genotype-to-environment interactions, and sex-to-genotype interactions, and we identify balancing selection mechanisms that maintain sympatric variation in mtDNA in natural populations of this model species.
2. Materials and Methods
Starting material for this study consisted of isofemale lines (IFL) of
Drosophila obscura that were constructed from females collected in the wild and that were previously genotyped for the
Cyt b gene [
56]. These IFLs were maintained in the laboratory on a standard corn-meal medium for multiple generations. All IFLs used were previously tested and were negative on
Wolbachia. In addition, IFLs used in this experiment were jointly analysed for microbiota composition, and no maternally transmitted bacteria were found [
56]. Four experimental blocks (EB) were formed, each from a specific locality. Three distinct sympatric haplotypes per block, representing IFLs from the same population were chosen in order to construct 9 MNILs.
The first EB consisted of three haplotypes O1, O2 and O8, from the ST population collection site 1 [
56]. Experimental block II was formed by backcrossing haplotypes O2, O9 and O10, which originated from the ST population collection site 2. Combinations of haplotypes O2, O4 and O3 belonging to the population SS, were used to make MNILs for the EB III. The MNILs of the fourth EB were constructed using haplotypes O2, O3 and O18 which come from the SG population. Specific IFLs were renamed to avoid confusion between EBs, as given in
Table 1.
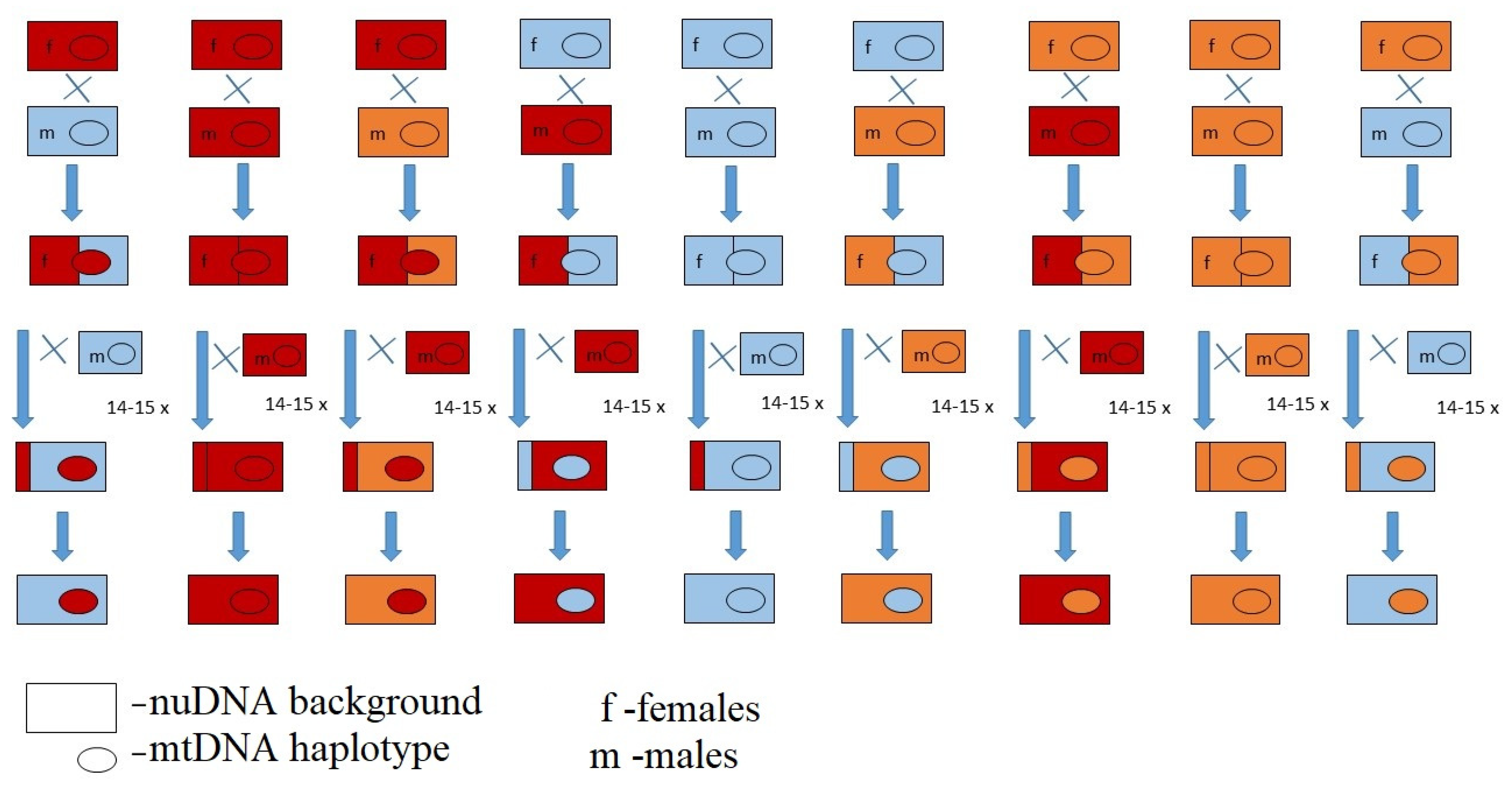
For each population, three selected IFLs were backcrossed for 14 generations, creating nine MNILs with all combinations of mitochondrial haplotypes (mtDNA) and nuclear genetic backgrounds (nuDNA). For the backcrossing procedure, we kept the vials with fly pupae in the dark such that we can collect virgin flies every morning. There is scarce information on the reproductive behaviour of
D. obscura; thus, we tested the methodology for collecting virgin individuals used for
D. subobscura [
25,
52]. From the pilot experiment, we concluded that
D. obscura flies will not mate in the first 24 h after eclosion when being in total dark, since no flies oviposited upon being collected from a dark box every 24 h. Each MNIL was created by mating 10 virgin females of a specific haplotype with 20 virgin males with the desired nuclear genetic background. The full backcrossing design is shown in
Figure 1. After 14 generations, flies were sequenced for the
Cyt b gene again to verify that all of the final 36 MNILs possess the appropriate haplotype.
In each block, we wanted to compare different mitochondrial haplotype triplets; in some, we compared specific single nucleotide polymorphisms that distinguish between most prevalent haplogroups, while some pairs differ in 6 or more mutations [
56]. The scheme of differences between haplotypes in each of the four EBs is given in
Figure 2.
In order to compare the fitness of the different combinations of mito-nuclear haplotypes, we conducted two grand experiments with several fitness components tested, in different generations due to a large number of individual flies of both sexes needed for each experiment. All experiments were conducted at two different temperatures. In all experiments for every block, we modelled three comparisons of haplotype pairs as well as the whole block with three different haplotype pairs compared together.
2.1. Desiccation Resistance
Desiccation resistance was the first conducted experiment after the 14th generation of backcrossing, two experimental temperatures were 16 and 19 °C, while the air humidity was set at 30%. For every specific MNIL, we had up to 40 flies per sex, for each of the two experimental temperatures. In some MNILs, we could not collect 40 flies per sex of the desired age. We had 38.8 females and 37.6 males on average for each MNIL per temperature. Individual virgin flies 5 to 7 days old were placed in small plastic modular containers with small holes for air circulation, where each module is being capped with the next making a total of two columns with 20 connected plastic containers per sex for each MNIL, for easier inspection. Containers were placed in two different rooms with regulated temperature and air-humidity levels. After the experiment was set, the flies were inspected every hour until effectively all flies had died. Mortality was determined by the inability of the Drosophila to keep an upright position or stand up after the plastic container had been shaken. In 4 EBs, each containing 9 MNILs, two temperatures, two sexes and up to 40 individuals had around 5500 flies tested for desiccation resistance.
2.2. Egg-to-Pupa-to-Adult Viability, Developmental Time and Sex Ratio Experiment
This experiment was conducted after the 15th generation of backcrossing, on two developmental temperatures (15 °C and 20 °C). For each MNIL, 40 virgin females and 60 virgin males 6–7 days old were placed in a bottle with a standard corn-meal medium for 5 days to mate. Another bottle with the same number of flies was kept as a backup. After 5 days flies were relocated to the new empty bottle which had a Petri dish instead of the cap. New bottles were placed upside down, standing on the Petri dish with corn-meal medium and yeast paste for flies to oviposit eggs. Each morning during the set-up phase, we replaced the old Petri dishes with the new ones. From the Petri dishes, we collected the oviposited eggs under binoculars and placed 20 eggs per vial for the viability experiment. For every MNIL, we made 30 replicas containing 20 eggs for each of the two experimental temperatures. Vials containing eggs were kept at two thermo-regulated chambers under constant temperature, light and dark cycles (15 °C and 12 h light/12 h dark regime and 20 °C and 16 h light/8 h dark regime) to imitate different seasons. Every two days we shuffled the vials inside the chambers to make sure that the temperature is constant for each vial. To sum up, with 4 EBs, each containing 9 MNILs, two temperatures, and 30 replicated vials with 20 transferred eggs we had over 43,000 eggs (4 × 9 × 2 × 30 × 20 = 43,200).
To measure developmental time 10 vials out of the 30 replicas for each temperature for each of the 36 MNIL, were randomly chosen, to be checked every day for hatched flies. Emerging adult flies were counted and sexed every day (in an air-conditioned room of the desired temperature) until the end of the experiment, with the number of pupae inside each vial counted as well. For the rest of the replicas, the number of adult flies, males, females and pupa was counted at the end of the eclosion after day 36.
2.3. Statistical Analysis
All statistical analyses were conducted using R v.4.0.3 [
58]. All Figures were made in R using the ggplot2 package [
59].
Desiccation data were analysed using a Cox proportional hazards model [
60] using survival package v.3.2-13 [
61] for each EB individually as well as for each of the three pairwise comparisons per EB, with no censoring since all flies died in the experiment. Mitochondrial haplotype (mtDNA), nuclear background (nuDNA), sex and temperature were used as fixed effects with all interactions of the four factors used as well (mtDNA × nuDNA × sex × temp). The proportional hazards assumption was checked with the cox.zph function, and where violated, the corresponding factors were stratified. Cox proportional models for EBs I and III were already presented at the 1
st international electronic conference on Entomology [
57]. Here, we present extended models with all factors and interactions included, in order to make the results of all four blocks comparable.
Developmental time data were analysed with a general linear model using the lmer function from the lme4 package in R [
62]. Full model with REML estimation and type III sums-of-squares was fitted with all interactions of fixed effects factors (mtDNA, nuDNA, sex and temperature) and replica number as random effect factor for each EB as well as for each pairwise comparison inside the EBs. LmerTest package [
63] was used to obtain
p values in ANOVA model fits.
Viability per vial was analysed with generalised linear models using the glm function in R. Egg to pupa (EtP), pupa to adult (PtA) and egg to adult (EtA) viability was scored for each pair of haplotypes compared respectively and for all four blocks apiece. Models had mitochondrial haplotype, nuclear background and temperature as fixed effects and all interactions of the three factors. All three component models, (EtA, EtP and PtA) had binomial error distribution and used the number of eggs/pupae per vial as the denominator.
Sex ratio data calculated as a proportion of males was analysed using the same general linear model with binomial error with the total number of eclosed adults per vial as the denominator. All pairwise comparisons were modelled individually, but also pulled together within each EB.
4. Discussion
We chose haplotypes for this experiment based on the differences in the Cyt b gene. One of our goals was to see if there are principal differences between the number and type (synonymous vs. nonsynonymous) of polymorphisms and the degree of fitness differences achieved by bearers of different mito-nuclear combinations.
First, we wanted to examine whether crossing lines with different
Cyt b 828A > G variants, a nonsynonymous substitution that divides two large groups of haplotypes in
D. obscura [
56], will result in a lower relative fitness. This was completely confirmed in EB I where we compared MNIL A which has G on bp 828 to MNILs B and C which have A, additionally, B and C have six synonymous mutations between them. Practically all fitness components in this EB (except pct. of males that had no factor significant in EB I) showed that the greater the difference in mtDNA sequence the greater the significance and impact on MNILs relative fitness. Comparison with the most different haplotypes (A and C) had both the mtDNA and mtDNA × nuDNA interactions as significant in the largest number of components assayed and the most similar haplotype comparison (B and C) had those factors as significant in the fewest components tested.
Contrary to these results, in other EBs fitness differences between haplotypes with aforementioned 828A > G, and other substitutions were not as correlated with the magnitude and type of sequence variation. Although we obtained predictable results in some components tested both for mtDNA and cytonuclear interaction effects, it was nowhere near the uniformity seen in the first EB, where it was almost as a rule. For some components, synonymous mutations proved more influential on fitness than nonsynonymous ones. Moreover, in some pairwise comparisons, combinations of more distant haplotypes showed greater relative fitness than combinations of close haplotypes.
It has been known for some time that even synonymous mutations have fitness consequences, which may sometimes be greater than nonsynonymous ones [
64]. It is also generally assumed that the greater the number of mutations the bigger the phenotypic differences between the MNILs, but that may not always be the case. The discrepancy in our work between different MNILs could be because the lines we used were sequenced only for the
Cyt b gene, and all other differences between mitochondrial DNA were not known. Thus, what looks more similar or more divergent when we look only in
Cyt b haplotypes may not be the case for the whole mitochondrial genome. In our experiment, we had the same haplotype pairwise comparison within the third and fourth EB, as LN and OP are the same combinations of
Cyt b gene haplotypes but coming from different populations. In almost all the components tested, we had contrasting results when comparing these two sets of identical haplotype comparisons. Apart from all the differences outside the
Cyt b gene that were not screened in our work, these MNILs, with the same
Cyt b haplotype originate from two different populations, and in turn should have completely different nuclear genetic backgrounds. This makes their comparisons difficult and further explains the discrepancy in our results.
During the backcrossing procedure that preceded the experiments, in IFL M we noted an unusually high percentage of females, while in O IFL, the observed portion of females was slightly elevated. The effect was apparent enough that we had difficulties collecting half as many M male flies, for the crossing procedure, as only about 5% hatched individuals were male. This effect is associated with the nuclear genome since the sex ratio experiment showed that this distortion is present only in MNILs that had M nuclear background. Maternally transmitted microorganisms can be excluded as a factor that causes this distortion by male killing or feminization, since it is not associated with a particular mtDNA haplotype which is expected to be transmitted jointly with maternally transmitted microorganisms. In addition, known maternally transmitted organisms were excluded with molecular genetic techniques including microbiome sequencing [
56]. Compared to other MNILs from the third EB, MNILs with M nuclear background did not show lower viability that could be caused by embryonic lethality in males. Rather, at 15 °C, they showed much higher EtA viability indicating the exclusion of Y chromosomes before fertilization.
This sex ratio distortion (SRD), in particular MNILs, which is associated with nuclear genetic background is probably caused by a meiotic drive mechanism. This sex ratio distortion is frequently found in
Drosophila [
65]. The first-ever case of the meiotic drive has been documented in
D. obscura [
66]. Gershenson found sex distortion in two out of nineteen IFLs formed by females collected in the wild. Studious experiments on deviations in the percentage of males that hatched from crossing different
D. obscura lines led him to a conclusion that a gene localized on the X chromosome prevents the genesis of functional spermatozoa without X chromosome. The meiotic drive has been confirmed in a broad range of phyla, and papers on different
Drosophila species showed that there are more than a few molecular mechanisms for it to cause SRD [
67,
68].
Although the majority of MNIL with M nuclear background had around 5% of males eclosed in viability experiment, MM MNIL, scored a twofold increase (12%) at a lower temperature. This observation is in line with the findings that sex ratio distortion is extremely temperature-sensitive, as spermatogenesis in higher temperatures results in a higher percentage of X chromosome bearing sperm [
69,
70]. Although the sex ratio was initially skewed in some MNILs, probably due to meiotic drive, our experimental design enabled us to capture and quantify the effects that mtDNA and nuDNA had on the survival of hatched individuals of a specific sex even in these lines with intrinsic skewed sex ratio.
This study supports a growing body of evidence of non-neutrality of mitochondrial DNA variation [
26,
71,
72], and more importantly, our results give weight to the adaptive significance of intra-population variation in mtDNA [
51,
73,
74]. As we hypothesized, almost all of our models for different fitness components showed that mito-nuclear interactions are more important as units for selection to act on than mitochondrial haplotypes on their own, as our results suggest. This should come as no surprise, considering that the
Cyt b gene is part of the respiratory complex III, which includes subunits coded by both genomes. In addition, as noted previously, haplotypes probably have differences in genes that comprise the other three of four complexes that have subunits coded by both genomes. If an mtDNA haplotype is coupled with a non-matching nuDNA background, a decline in adaptive value is expected, as the subunits from two genomes have to be co-adapted for the optimal energy production in the mitochondria.
As expected, our experimental model identified sex-specific differences in the fitness of bearers of different mtDNA haplotypes. While this effect was significant in seven out of twelve pairwise comparisons in the desiccation experiment, this male-specific mutational load was noticeable in only two comparisons for the developmental time component. This effect was indirectly measured as an effect of mtDNA in the sex ratio component where it was significant in three out of twelve pairwise comparisons. This observation is in line with the mother’s curse hypothesis, a phenomenon frequently found while measuring the effects of mtDNA on life history [
47,
48,
50,
55]. Due to maternal inheritance of mtDNA, mutations that are disadvantageous only in males, and have no effect or are advantageous to females, cannot be purged by natural selection.
One of our goals was to test whether different combinations of mitochondrial and nuclear genomes show different fitness ranks depending on the sex of the individual. This finding would support the theoretical presumptions that SSS is responsible for maintaining stable sympatric mitochondrial and mito-nuclear variation [
45,
46]. Experimental support for this type of balancing selection comes from several previous experiments. When analysing cytonuclear interactions between the X chromosome and mitochondrial DNA, Rand [
35] observed the action of SSS in
D. melanogaster. In a viability experiment with 25 mtDNA haplotypes scored on three nuclear genetic backgrounds, Dowling et al. [
50] found the interaction of mtDNA × nuDNA × sex significant, but only in the first out of three repeated EBs. In the second EB, they could not test this interaction due to missing data, but in the third EB and in overall analyses (across EBs), they did not find evidence of sex specificity of mito-nuclear effects on viability. Jelic et al. [
52] analysed a series of key life-history traits of
Drosophila subobscura MNILs made from three sympatric mtDNA haplotypes. They unequivocally found sex-specific effects (mtDNA × nuDNA × sex) in two experimental modules for adult longevity and indirectly in one module (mtDNA × nuDNA) for egg-to-adult viability when analysing the proportion of males hatching [
52]. Conversely, they found no evidence of SSS in any of the experimental modules for desiccation resistance. Similarly, using seed beetles
Acanthoscelides obtectus, Đorđević et al. performed mortality assays and tested mito-nuclear effects on survival [
55]. Although they did not obtain significant mtDNA × nuDNA interaction in their models, mtDNA × nuDNA × sex interaction was significant in model for variance in lifespan, but not in two survival models. In their work, they also analysed the effects of mtDNA and nuDNA as well as sex on the activity of METC complexes. Variation in ETC activity was significantly influenced by sex-specific mito-nuclear interactions in METC complexes I and II, contrastingly they did not find this interaction significant in complexes III and IV. In our experimental design, the role of SSS on mito-nuclear variation was scored directly in desiccation resistance and developmental time as a significant effect of interaction between mtDNA × nuDNA × sex. Conversely, a significant interaction between mtDNA × nuDNA for a sex ratio of hatched adults is also an indirect measure of SSS. In our model species, we found the interaction between mito-nuclear genotype and sex to be statistically significant only in two out of twelve pairwise comparisons for developmental time and desiccation resistance experiments each (if desiccation resistance is scored jointly with previously analysed blocks [
57]). Signature of SSS was observed in five out of twelve comparisons in the sex ratio experiment. Our findings support theoretical presumptions that SSS is involved in the maintenance of sympatric mtDNA variation.
The aim of this study was also to test whether different mitochondrial haplotypes or combinations of mitochondrial and nuclear genomes show different fitness ranks depending on the experimental temperature. This finding would support the idea that temperature variation may promote stable sympatric genetic variation. Temperature is of key importance in metabolic processes. Numerous papers on the subject point to the particular sensitivity of OXYPHOS enzyme complexes to temperature [
54,
75,
76,
77]. The connection between mtDNA variation and the temperature has been observed as a clinal shift of haplotype frequencies along latitude [
78,
79,
80] and altitude [
81,
82]. Additionally, the optimal function of subunits coded by two different genomes may depend on the thermal environment that the reactions are taking place. While some combinations of mtDNA and nuDNA may be supreme in one thermal environment that may not be the case in others.
Using seed beetles Callosobruchus maculatus, Immonen et al. [
54] had different mtDNA haplotypes compete on two different experimental temperatures. Their results claim that temperature is influencing mtDNA evolution to some extent, most likely through mito-nuclear interactions. Similarly, another study [
22] measuring EtA development time on the same model organism and two temperatures showed the significance of G × G × E interactions. This effect of temperature-specific fitness of MNILs was found once more, [
41] using the same experimental lines in another experiment, measuring metabolic rate. Similarly, Rand [
83] using Drosophila as a model found that altered dietary or oxygen environments modify the fitness of mito-nuclear haplotypes.
Research that analyses the fitness of sympatric mitochondrial variation in regard to the extrinsic environment including temperature is scarce [
14]. For example, Dowling et al. [
50] analysed a single panmictic laboratory population and showed that multiple mitochondrial haplotypes can be preserved within it. Their experiment consisted of three repeated measures (three blocks) of the same experiment, and as they suggest relative fitness of the cytonuclear combination is dependent on the environment that they exist in, as they find the effect of the block to be substantial. They attributed this to the unforeseeable heterogeneity of environmental factors across blocks [
50]. The effects of temperature or other environmental factors on fitness ranks of mito-nuclear genotypes between populations suggest [
14,
22,
50] that this mechanism could potentially support sympatric variation as well. Although the haplotypes used by Immonen et al. [
54] originate from geographically distant populations, their haplogroups have all been found to segregate sympatrically in the same West African population, which gives more support to the above claims of temperature-specific epistasis.
Data for our model species included temperature as an environmental factor in the experiments, and we showed that the genotype × temperature interaction has a significant effect on
Drosophila fitness in all components assayed in this work. This effect was especially important in desiccation resistance (if analysed jointly with [
57]), and viability experiments, with both mtDNA × nuDNA × temp and mtDNA × temp interactions being highly significant in both pairwise and whole block models. The abundance of this type of interaction in our data supports the presumption based on interpopulation research [
22,
41,
54] that genotype-by-environment interactions are also important for maintaining stable intra-population mtDNA variation in nature [
22,
41] and compels for further research to be performed on this phenomenon.
Taken together, the fitness assays performed on D. obscura show the complexity of maintenance of sympatric mtDNA variation. Different balancing selection mechanisms may operate simultaneously in upholding joint genomic polymorphism in the same model. Our results give more weight to environment-mediated selection compared to SSS. However, the question stands as to what extent these results could be extrapolated to variation in natural habitats. Our experiment was performed on arbitrarily chosen temperatures, compared to the continual and unpredictable variation in nature. While sex is a discrete variable, the temperature is continuous, and different results could have been observed if fitness was compared at other experimental temperatures or other environmental conditions.


 ,
, 




{kind=link}
{kind=link}