
Phenology and Potential Fecundity of Neoleucopis kartliana in Greece

 ,
, 
 and
and 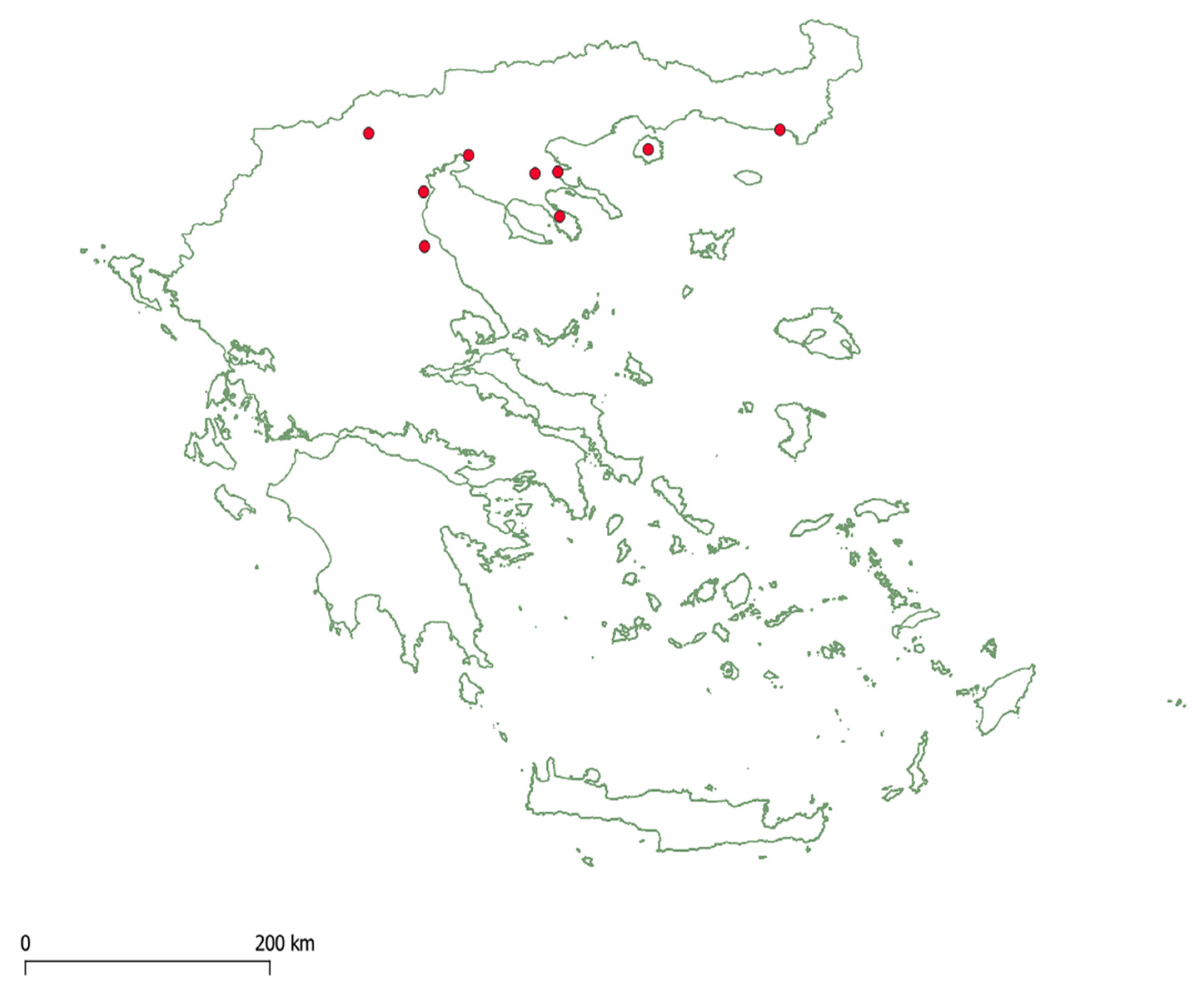{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
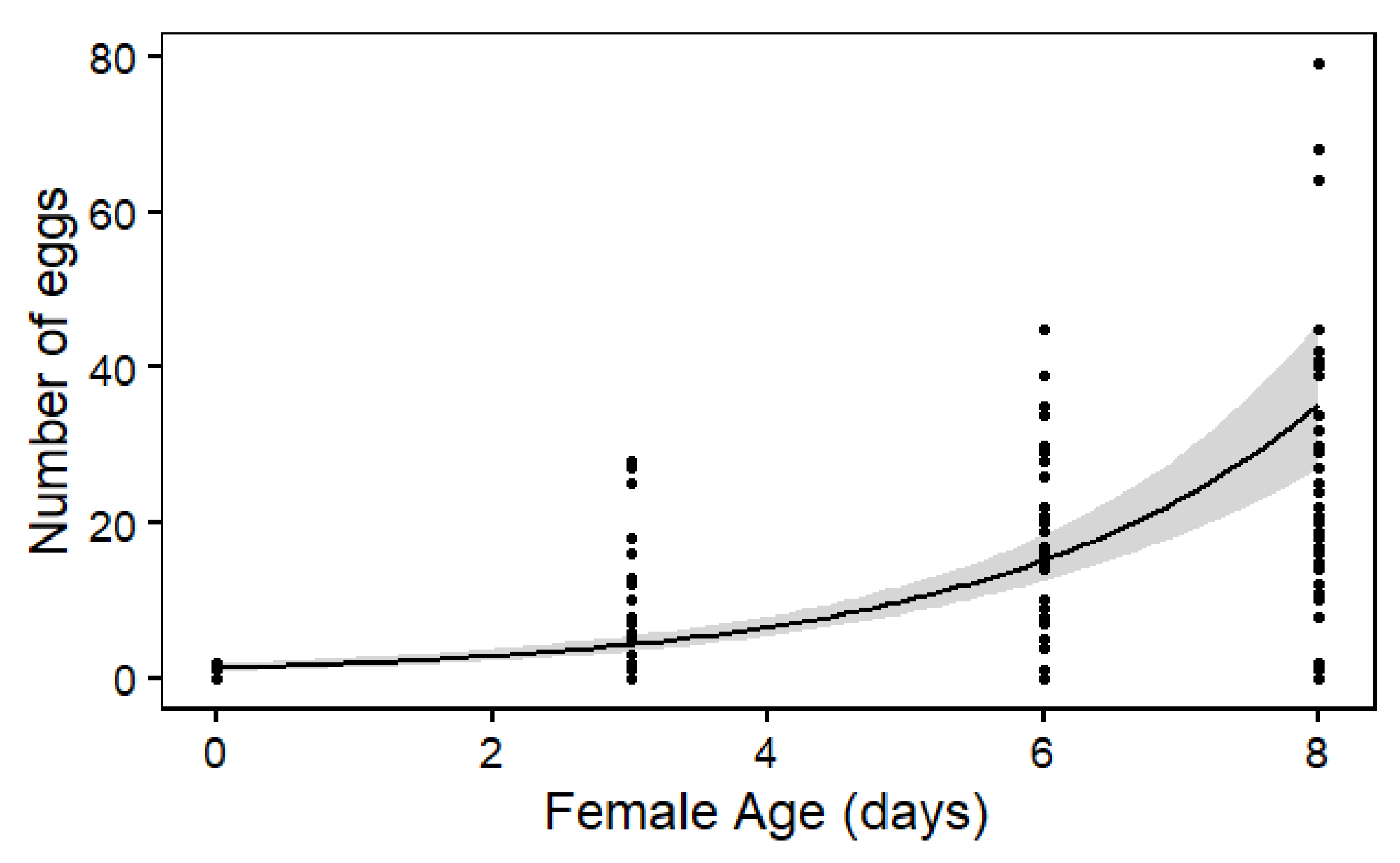
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gaimari, S.D.; Turner, W.J. Methods for rearing aphidophagous Leucopis spp. (Diptera: Chamaemyiidae). J. Kans. Entomol. Soc. 1996, 69, 363–369. [Google Scholar]
- Gaimari, S.D. Two new genera of Nearctic Chamaemyiidae (Diptera: Lauxanioidea) associated with Cinara aphids (hemiptera: Aphididae) on Pinus. Zootaxa 2020, 4852, 61–82. [Google Scholar] [CrossRef] [PubMed]
- McLean, I.F.G. Leucopis psylliphaga sp. n., a new species of silverfly (Diptera, Chamaemyiidae) from Britain. Dipter. Dig. 1998, 5, 49–54. [Google Scholar]
- Ross, D.W.; Gaimari, S.D.; Kohler, G.R.; Wallin, K.F.; Grubin, S.M. Chamaemyiid predators of the hemlock woolly adelgid from the Pacific Northwest. In ImplementatIon and Status of BIologIcal Control of the Hemlock Woolly AdelgId; Onken, B., Reardon, R., Eds.; US Department of Agriculture, Forest Service, Forest Health Technology Enterprise Team: Morgantown, WV, USA, 2011; pp. 97–106. [Google Scholar]
- Krsteska, V. Chamaemyiidae (Diptera)-Predators of aphids on tobacco. Tutun/Tobacc 2015, 65, 30–38. [Google Scholar]
- Balci, Ş.; Ülgentürk, S. New records of Chamaemyiidae and Cryptochaetidae (Diptera) on scale insects (Hemiptera: Coccomorpha) in Turkey. Turk. Biyol. Mucadele Derg. 2019, 10, 127–132. [Google Scholar]
- Grubin, S.M.; Ross, D.W.; Wallin, K.F. Prey Suitability and Phenology of Leucopis spp. (Diptera: Chamaemyiidae) Associated with Hemlock Woolly Adelgid (Hemiptera: Adelgidae) in the Pacific Northwest. Environ. Entomol. 2011, 40, 1410–1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McAlpine, J.F.; Tanasijtshuk, V.N. Identity of Leucopis argenticollis and description of a new species (Diptera: Chamaemyiidae). Can. Entomol. 1972, 104, 1865–1875. [Google Scholar] [CrossRef]
- Ohnisni, S. Effects of population density and temperature condition on fitness in Drosophila melanogaster II. fecundity and mortality. Jpn. J. Genet. 1976, 51, 305–314. [Google Scholar] [CrossRef] [Green Version]
- Akoh, J.I.; Aigbodion, F.I.; Kumbak, D. Studies on the effect of larval diet, adult body weight, size of blood-meal and age on the fecundity of Culex Quinquefasciatus (Diptera: Culicidae). Int. J. Trop. Insect Sci. 1992, 13, 177–181. [Google Scholar] [CrossRef]
- Adams, T.S.; Nelson, D.R. The influence of diet on ovarian maturation, mating, and pheromone production in the housefly, Musca domestica. Invertebr. Reprod. Dev. 1990, 17, 193–201. [Google Scholar] [CrossRef]
- Pastor, B.; Čičková, H.; Kozánek, M.; Anabel, M.S.; Takáč, P.; Rojo, S. Effect of the size of the pupae, adult diet, oviposition substrate and adult population density on egg production in Musca domestica (Diptera: Muscidae). Eur. J. Entomol. 2011, 108, 587–596. [Google Scholar] [CrossRef] [Green Version]
- Aluja, M.; Díaz-Fleischer, F.; Papaj, D.R.; Lagunes, G.; Sivinski, J. Effects of age, diet, female density, and the host resource on egg load in Anastrepha ludens and Anastrepha obliqua (Diptera: Tephritidae). J. Insect Physiol. 2001, 47, 975–988. [Google Scholar] [CrossRef]
- Papanastasiou, S.A.; Nakas, C.T.; Carey, J.R.; Papadopoulos, N.T. Condition-dependent effects of mating on longevity and fecundity of female medflies: The interplay between nutrition and age of mating. PLoS ONE 2013, 8, e70181. [Google Scholar] [CrossRef] [PubMed]
- McCann, S.; Day, J.F.; Allan, S.; Lord, C.C. Age modifies the effect of body size on fecundity in Culex quinquefasciatus Say (Diptera: Culicidae). J. Vector Ecol. 2009, 34, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Lavagnino, N.J.; Fanara, J.J.; Mensch, J. Comparison of overwintering survival and fertility of Zaprionus indianus (Diptera: Drosophilidae) flies from native and invaded ranges. J. Therm. Biol. 2020, 87, 102470. [Google Scholar] [CrossRef]
- Broufas, G.D.; Pappas, M.L.; Koveos, D.S. Effect of relative humidity on longevity, Ovarian maturation, and egg production in the olive fruit fly (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 2009, 102, 70–75. [Google Scholar] [CrossRef] [Green Version]
- Culliney, T.W.; Beardsley, J.W.; Drea, J.J. Population regulation of the eurasian pine adelgid (Homoptera: Adelgidae) in Hawaii. J. Econ. Entomol. 1988, 81, 142–147. [Google Scholar] [CrossRef]
- Zondag, R.; Nuttall, M.J. Pineus laevis (Maskell), pine twig chermes or pine woolly aphid (Homoptera: Adelgidae). In Review of Biological Control of Invertebrate Pests and Weeds in New Zealand 1874 to 1987; Cameron, P.J., Hill, R.L., Bain, J., Thomas, W.P., Eds.; CAB International Institute of Biological Control: Wallingford, UK, 1989; pp. 295–297. [Google Scholar]
- Mills, N.J. Biological control of forest aphid pests in Africa. Bull. Entomol. Res. 1990, 80, 31–36. [Google Scholar] [CrossRef]
- Zúñiga, S. Eighty years of biological control in Chile. Historical review and evaluation of the projects undertaken (1903–1983). Agric. Tec. 1985, 45, 175–184. [Google Scholar]
- Garonna, A.P.; Viggiani, G. The establishment in Italy of Neoleucopis kartliana (Tanasjtshuk) (Diptera: Chamaemyiidae), predator of Marchalina hellenica (Gennadius) (Hemiptera: Margarodidae). In Proceedings of the XXIII Italian National Congress of Entomology, Genoa, Italy, 13–16 June 2011; p. 346. [Google Scholar]
- Gounari, S. Studies on the phenology of Marchalina Hellenica (gen.) (Hemiptera: Coccoidea, Margarodidae) in relation to honeydew flow. J. Apic. Res. 2006, 45, 8–12. [Google Scholar] [CrossRef]
- Yeşil, A.; Gürkan, B.; Saraçoǧlu, Ö.; Zengin, H. Effect of the pest Marchalina hellenica Gennadius (Homoptera, Margarodidae) on the growth parameters of Pinus brutia Ten. in Mugla region (Turkey). Pol. J. Ecol. 2005, 53, 451–458. [Google Scholar]
- Petrakis, P.V.; Spanos, K.; Feest, A. Insect biodiversity reduction of pinewoods in southern Greece caused by the pine scale (Marchalina hellenica). For. Syst. 2011, 20, 27–41. [Google Scholar] [CrossRef] [Green Version]
- Thrasyvoulou, A.; Manikis, J. Some physicochemical and microscopic characteristics of Greek unifloral honeys. Apidologie 1995, 26, 441–452. [Google Scholar] [CrossRef] [Green Version]
- Mendel, Z.; Branco, M.; Battisti, A. Invasive sap-sucker insects in the mediterranean basin. In Insects and Diseases of Mediterranean Forest Systems; Paine, T.D., Lieutier, F., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 261–291. [Google Scholar]
- Downham, R.; Gavran, M. Australian Plantation Statistics 2017 Update; Australian Government Department of Agriculture and Water Resources: Canberra, Australia, 2017; p. 12.
- Avtzis, D.N.; Lubanga, U.K.; Lefoe, G.K.; Kwong, R.M.; Eleftheriadou, N.; Andreadi, A.; Elms, S.; Shaw, R.; Kenis, M. Prospects for classical biological control of Marchalina hellenica in Australia. BioControl 2020, 65, 413–423. [Google Scholar] [CrossRef]
- Ülgentürk, S.; Szentkirályi, F.; Uygun, N.; Fent, M.; Gaimari, S.D.; Civelek, H.; Ayhan, B. Predators of Marchalina hellenica (Hemiptera: Marchalinidae) on pine forests in Turkey. Phytoparasitica 2013, 41, 529–537. [Google Scholar] [CrossRef]
- Oǧuzoǧlu, S.; Avcl, M.; Ipekdal, K. Predators of the giant pine scale, Marchalina hellenica (Gennadius 1883; Hemiptera: Marchalinidae), out of its natural range in Turkey. Open Life Sci. 2021, 16, 682–694. [Google Scholar] [CrossRef] [PubMed]
- Gaimari, S.D.; Milonas, P.; Souliotis, C. Notes on the taxonomy, biology and distribution of Neoleucopis kartliana (Diptera: Chamaemyiidae). Folia Heyrovskyana A. Folia Heyrovskyana 2007, 15, 7–16. [Google Scholar]
- Hodgson, C.; Gounari, S. Morphology of Marchalina hellenica (Gennadius) (Hemiptera: Coccoidea: Marchalinidae) from Greece, with a discussion on the identity of M. caucasica Hadzibeyli from the Caucasus. Zootaxa 2006, 32, 1–32. [Google Scholar] [CrossRef]
- Teodorescu, I.; Maican, S. Diversity of entomofauna (Hymenoptera, Diptera, Coleoptera) in the Comana natural park (Romania). Rom. J. Biol. 2014, 59, 17–34. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Clark, R.C.; Brown, N.R. Studies of predators of the balsam woolly aphid, Adelges piceae (Ratz.) (Homoptera: Adelgidae): III. Field identification and some notes on the biology of Neoleucopis pinicola Mall. (Diptera: Chamaemyiidae). Can. Entomol. 1957, 89, 404–409. [Google Scholar] [CrossRef]
- Pschorn-Walcher, H.; Zwölfer, H. The predator complex of the white-fir woolly aphids (Genus Dreyfusia, Adelgidae). Z. Angew. Entomol. 1956, 39, 63–75. [Google Scholar] [CrossRef]
- Eichhorn, O. Problems of the population dynamics of silver fir woolly aphids, genus Adelges (=Dreyfusia), Adelgidae. Z. Angew. Entomol. 1968, 61, 157–214. [Google Scholar] [CrossRef]
- Altermatt, F. Climatic warming increases voltinism in european butterflies and moths. Proc. R. Soc. B Biol. Sci. 2010, 277, 1281–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawley, M.J. The population biology of invaders. Philosophical Transactions of the Royal Society of London B. Biol. Sci. 1986, 314, 711–731. [Google Scholar]
- Hokkanen, H.M.T.; Sailer, R.I. Success in classical biological control. Crit. Rev. Plant Sci. 1985, 3, 35–72. [Google Scholar] [CrossRef]
- Murdoch, W.W.; Swarbrick, S.L.; Briggs, C.J. Biological control: Lessons from a study of California red scale. Popul. Ecol. 2006, 48, 297–305. [Google Scholar] [CrossRef]
- Seehausen, M.L.; Afonso, C.; Jactel, H.; Kenis, M. Classical biological control against insect pests in Europe, North Africa, and the Middle East: What influences its success? NeoBiota 2021, 65, 169–191. [Google Scholar] [CrossRef]
- Satar, S.; Raspi, A.; Özdemir, I.; Tusun, A.; Karacaoğlu, M.; Benelli, G. Seasonal habits of predation and prey range in aphidophagous silver flies (Diptera Chamaemyiidae), an overlooked family of biological control agents. Bull. Insectol. 2015, 68, 173–180. [Google Scholar]
- Dietschler, N.J.; Bittner, T.D.; Trotter, R.T.; Fahey, T.J.; Whitmore, M.C. Biological Control of Hemlock Woolly Adelgid: Implications of Adult Emergence Patterns of Two Leucopis spp. (Diptera: Chamaemyiidae) and Laricobius nigrinus (Coleoptera: Derodontidae) Larval Drop. Environ. Entomol. 2021, 50, 803–813. [Google Scholar] [CrossRef]
- Gaimari, S.D.; Havill, N.P. A new genus of Chamaemyiidae (Diptera: Lauxanioidea) predaceous on Adelgidae (Hemiptera), with a key to chamaemyiid species associated with Pinaceae-feeding Sternorrhyncha. Zootaxa 2021, 5067, 1–39. [Google Scholar] [CrossRef]
- Fisher, R.A. The Genetical Theory of Natural Selection; Clarendon Press: Oxford, UK, 1930. [Google Scholar]
- Gaimari, S.D.; Turner, W.J. Behavioral observations on the adults and larvae of Leucopis ninae and L. gaimarii (Diptera: Chamaemyiidae), predators of Russian wheat aphid, Diuraphis noxia (Homoptera: Aphididae). J. Kans. Entomol. Soc. 1997, 70, 153–159. [Google Scholar]
- Wäckers, F.L.; van Rijn, P.C.J.; Heimpel, G.E. Honeydew as a food source for natural enemies: Making the best of a bad meal? Biol. Control 2008, 45, 176–184. [Google Scholar] [CrossRef] [Green Version]
- Crosskey, R.W. Introduction to the Diptera. In Medical Insects and Arachnids; Lane, R.P., Crosskey, R.W., Eds.; Springer: Dordrecht, The Netherlands, 1993; pp. 51–77. [Google Scholar]
- Jervis, M.A.; Heimpel, G.E.; Ferns, P.N.; Harvey, J.A.; Kidd, N.A.C. Life-history strategies in parasitoid wasps: A comparative analysis of “ovigeny”. J. Anim. Ecol. 2001, 70, 442–458. [Google Scholar] [CrossRef] [Green Version]
- Benelli, G.; Bonsignori, G.; Stefanini, C.; Raspi, A. First quantification of courtship behavior in a silver fly, Leucopis palumbii (Diptera: Chamaemyiidae): Role of visual, olfactory and tactile cues. J. Insect Behav. 2014, 27, 462–477. [Google Scholar] [CrossRef]
- Clausen, C.P. Chermidae. In Introduced Parasites and Predators of Arthropod Pests and Weeds: A World Review; Clausen, C.P., Ed.; USDA Agricultural Handbook No. 480; US Department of Agriculture: Washington, DC, USA, 1978; pp. 49–55. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eleftheriadou, N.; Lubanga, U.; Lefoe, G.; Seehausen, M.L.; Kenis, M.; Kavallieratos, N.G.; Avtzis, D.N. Phenology and Potential Fecundity of Neoleucopis kartliana in Greece. Insects 2022, 13, 143. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020143
Eleftheriadou N, Lubanga U, Lefoe G, Seehausen ML, Kenis M, Kavallieratos NG, Avtzis DN. Phenology and Potential Fecundity of Neoleucopis kartliana in Greece. Insects. 2022; 13(2):143. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020143
Chicago/Turabian StyleEleftheriadou, Nikoleta, Umar Lubanga, Greg Lefoe, M. Lukas Seehausen, Marc Kenis, Nickolas G. Kavallieratos, and Dimitrios N. Avtzis. 2022. "Phenology and Potential Fecundity of Neoleucopis kartliana in Greece" Insects 13, no. 2: 143. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020143