
Illumina Short-Read Sequencing of the Mitogenomes of Novel Scarites subterraneus Isolates Allows for Taxonomic Refinement of the Genus Scarites Fabricius 1775, within the Carabidae Family


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Species Isolation
2.2. DNA Extraction
2.3. DNA Sequencing and Annotation
2.4. Phylogenetic Trees and ANI Calculations
3. Results and Discussion
3.1. Species Identification Based on Morphology
3.2. Genome Assembly
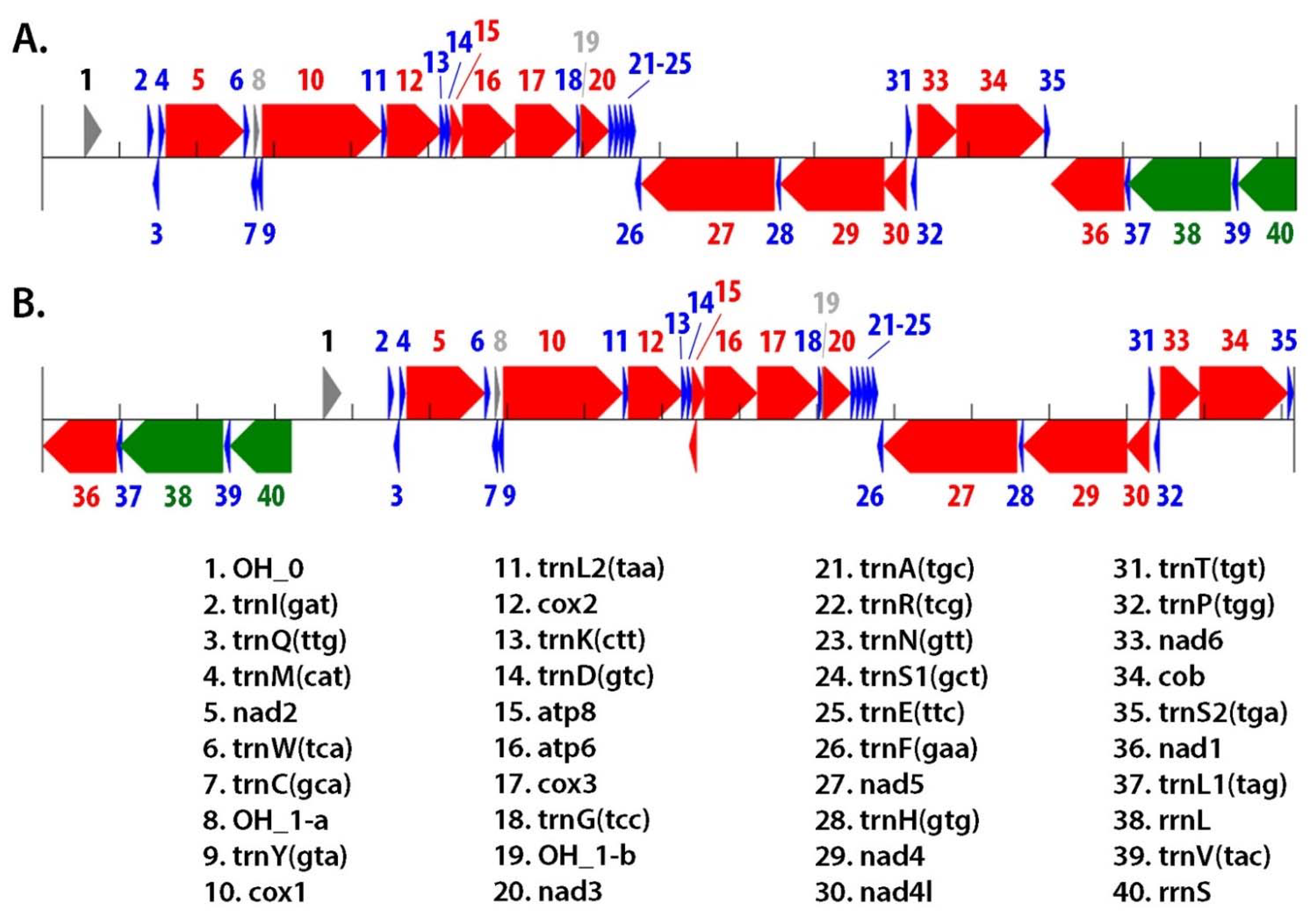
3.3. Mitogenomes Summary
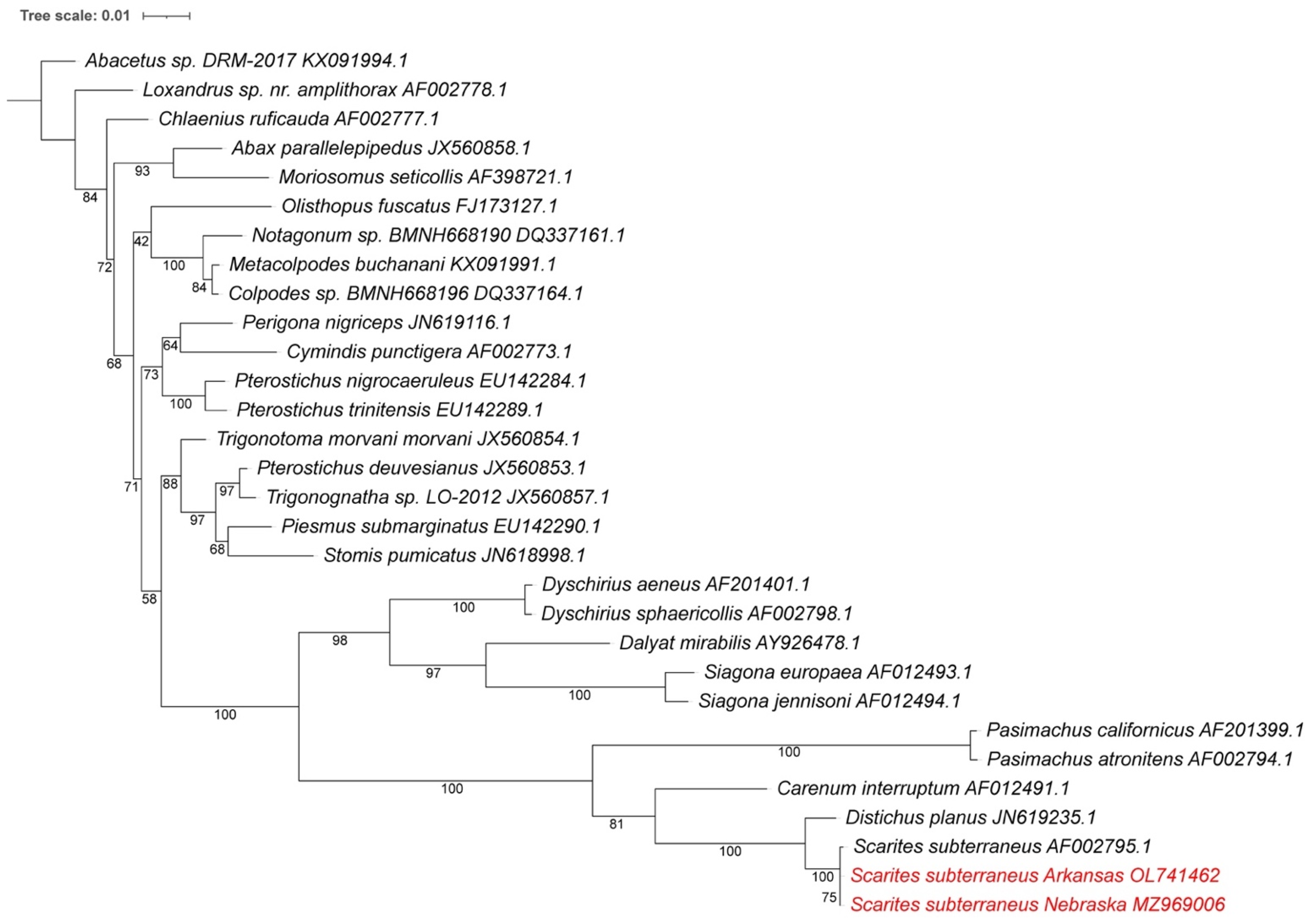
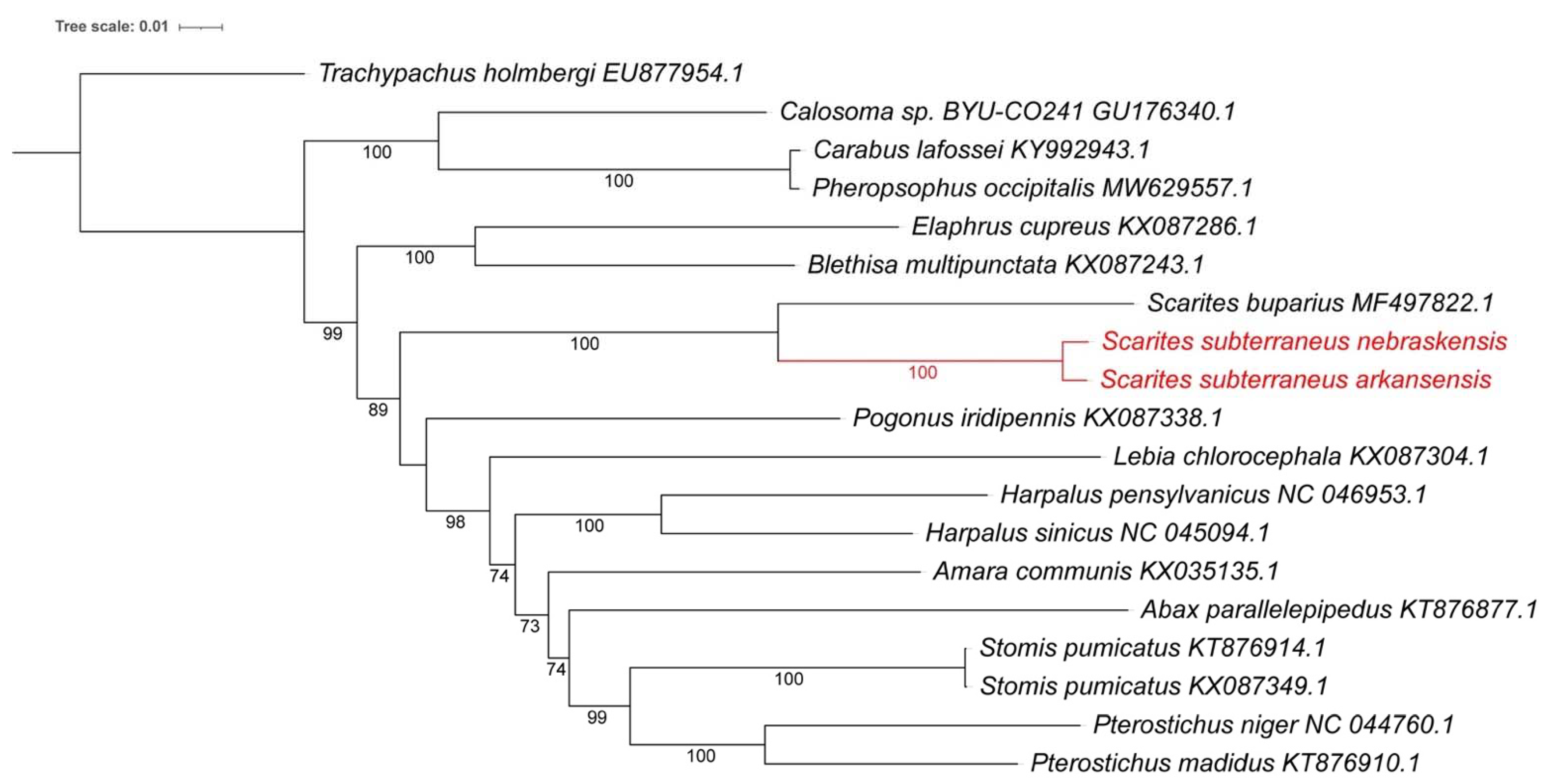
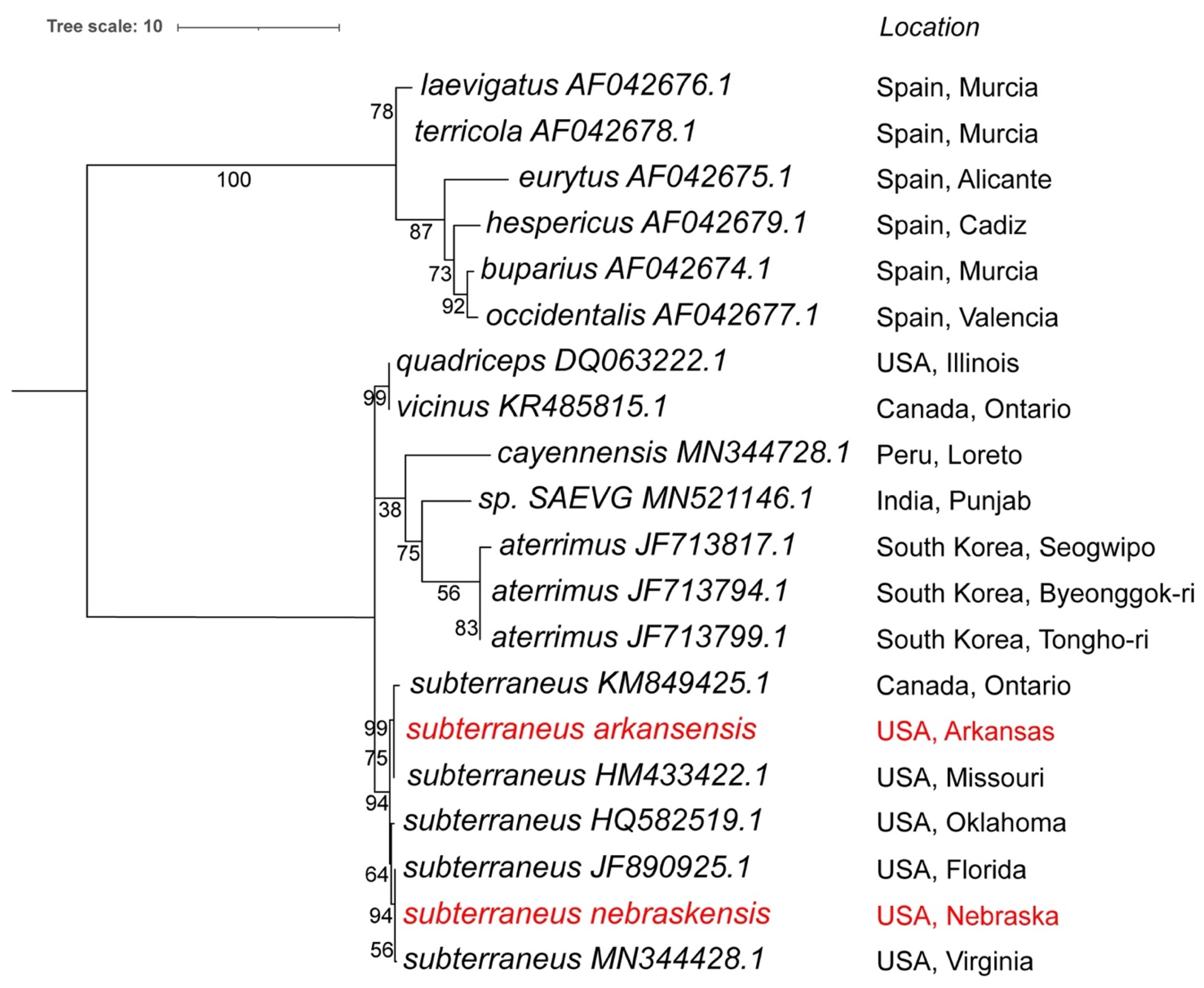
3.4. Mitogenome Phylogenetic Analysis
3.5. Microbial Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hlavac, T.F. A Review of the Species of Scarites (Antilliscaris) (Coleoptera: Carabidae) With Notes on Their Morphology and Evolution. Psyche A J. Èntomol. 1969, 76, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Bousquet, Y.; Larochelle, A. Catalogue of the Geadephaga (coleoptera: Trachypachidae, Rhysodidae, Carabidae including Cicindelini) of America north of Mexico. Memoirs Èntomol. Soc. Can. 1993, 125, 397. [Google Scholar] [CrossRef]
- Arnett, R.H., Jr.; Thomas, M.C. (Eds.) American Beetles, Volume I: Archostemata, Myxophaga, Adephaga, Polyphaga: Staphyliniformia; CRC Press LLC: Boca Raton, FL, USA, 2000. [Google Scholar]
- Ratcliffe, B.C.; Paulsen, M.J. The Scarabaeoid Beetles of Nebraska (Coleoptera: Scarabaeoidea). Bull. Univ. Neb. State Mus. 2008, 22, 1–570. [Google Scholar]
- Bousquet, Y. Catalogue of Geadephaga (Coleoptera: Adephaga) of America, north of Mexico. ZooKeys 2012, 245, 1–1722. [Google Scholar] [CrossRef] [Green Version]
- Lundgren, J.G. Ground beetles as weed control agents: Effects of farm management on granivory. Am. Èntomol. 2005, 51, 224–226. [Google Scholar] [CrossRef] [Green Version]
- Brooks, D.R.; Bater, J.E.; Clark, S.J.; Monteith, D.T.; Andrews, C.; Corbett, S.J.; Beaumont, D.A.; Chapman, J. Large carabid beetle declines in a United Kingdom monitoring network increases evidence for a widespread loss in insect biodiversity. J. Appl. Ecol. 2012, 49, 1009–1019. [Google Scholar] [CrossRef]
- Raupp, M.J. An Unusual but Not Unpleasant Home Invasion by a Beneficial Beetle: Big-Headed Ground Beetle, Scarites Subterraneus. 2020. Available online: http://bugoftheweek.com/blog/2020/10/12/an-unusual-but-not-unpleasant-home-invasion-by-a-beneficial-beetle-big-headed-ground-beetle-scarites-subterraneus (accessed on 24 December 2021).
- Bousquet, Y.; Skelley, P.E. Description of a New Species ofScaritesFabricius (Coleoptera: Carabidae) from Florida. Coleopt. Bull. 2010, 64, 45–49. [Google Scholar] [CrossRef]
- Erwin, T.L. Biodiversity at its utmost: Tropical Forest beetles. In Biodiversity II: Understanding and Protecting Our Biological Resources; Wilson, M.L., Don, E., Wilson, E.O., Eds.; Joseph Henry Press: Washington, DC, USA, 1997; pp. 27–40. [Google Scholar]
- Schowalter, T.D. Insects and Sustainability of Ecosystem Services; CRC Press: Boca Raton, FL, USA, 2013. [Google Scholar] [CrossRef]
- Slade, E.M.; Bagchi, R.; Keller, N.; Philipson, C.D. When do more species maximize more ecosystem services? Trends Plant Sci. 2019, 24, 790–793. [Google Scholar] [CrossRef]
- McKenna, D.D. Beetle genomes in the 21st century: Prospects, progress and priorities. Curr. Opin. Insect Sci. 2018, 25, 76–82. [Google Scholar] [CrossRef]
- Li, F.; Zhao, X.; Li, M.; He, K.; Huang, C.; Zhou, Y.; Li, Z.; Walters, J.R. Insect genomes: Progress and challenges. Insect Mol. Biol. 2019, 28, 739–758. [Google Scholar] [CrossRef] [Green Version]
- Löbl, I.; Smetana, A. (Eds.) Catalogue of Palaearctic Coleoptera. I; Apollo Books: Stenstrup, Denmark, 2003. [Google Scholar]
- Ober, K.A. Phylogenetic relationships of the carabid subfamily Harpalinae (Coleoptera) based on molecular sequence data. Mol. Phylogenetics Evol. 2002, 24, 228–248. [Google Scholar] [CrossRef]
- Ober, K.A.; Maddison, D.R. Phylogenetic Relationships of Tribes Within Harpalinae (Coleoptera: Carabidae) as Inferred from 28S Ribosomal DNA and theWinglessGene. J. Insect Sci. 2008, 8, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Gustafson, G.T.; Baca, S.M.; Alexander, A.M.; Short, A.E.Z. Phylogenomic analysis of the beetle suborder Adephaga with comparison of tailored and generalized ultraconserved element probe performance. Syst. Èntomol. 2019, 45, 552–570. [Google Scholar] [CrossRef]
- Misof, B.; Liu, S.; Meusemann, K.; Peters, R.S.; Donath, A.; Mayer, C.; Frandsen, P.B.; Ware, J.; Flouri, T.; Beutel, R.G.; et al. Phylogenomics resolves the timing and pattern of insect evolution. Science 2014, 346, 763–767. [Google Scholar] [CrossRef]
- Cameron, S.L. How to sequence and annotate insect mitochondrial genomes for systematic and comparative genomics research. Syst. Èntomol. 2014, 39, 400–411. [Google Scholar] [CrossRef] [Green Version]
- López-López, A.; Vogler, A.P. The mitogenome phylogeny of Adephaga (Coleoptera). Mol. Phylogenetics Evol. 2017, 114, 166–174. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Li, X.; Song, N.; Tang, H.; Yin, X. The Mitochondrial Genome of Amara aulica (Coleoptera, Carabidae, Harpalinae) and Insights into the Phylogeny of Ground Beetles. Genes 2020, 11, 181. [Google Scholar] [CrossRef] [Green Version]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data; Babraham Institute: Cambridge, UK, 2010; Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 24 December 2021).
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comp. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Zerbino, D.R.; Birney, E. Velvet: Algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 2008, 18, 821–829. [Google Scholar] [CrossRef] [Green Version]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PloS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [Green Version]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L.; et al. Improvements to PATRIC, the all-bacterial Bioinformatics Database and Analysis Resource Center. Nucleic Acids Res. 2016, 45, D535–D542. [Google Scholar] [CrossRef] [PubMed]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol. Phylogenetics Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef]
- Donath, A.; Jühling, F.; Al-Arab, M.; Bernhart, S.H.; Reinhardt, F.; Stadler, P.F.; Middendorf, M.; Bernt, M. Improved annotation of protein-coding genes boundaries in metazoan mitochondrial genomes. Nucleic Acids Res. 2019, 47, 10543–10552. [Google Scholar] [CrossRef] [PubMed]
- Timmermans, M.J.; Vogler, A.P. Phylogenetically informative rearrangements in mitochondrial genomes of Coleoptera, and monophyly of aquatic elateriform beetles (Dryopoidea). Mol. Phylogenetics Evol. 2012, 63, 299–304. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Wood, D.E.; Salzberg, S.L. Kraken: Ultrafast metagenomic sequence classification using exact alignments. Genome Biol. 2014, 15, R46. [Google Scholar] [CrossRef] [Green Version]
- Ondov, B.D.; Bergman, N.H.; Phillippy, A.M. Interactive metagenomic visualization in a Web browser. BMC Bioinform. 2011, 12, 385. [Google Scholar] [CrossRef] [Green Version]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [Green Version]
- Richter, M.; Rosselló-Móra, R.; Oliver Glöckner, F.O.; Peplies, J. JSpeciesWS: A web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 2016, 32, 929–931. [Google Scholar] [CrossRef] [PubMed]
- Nichols, S.W. Two new flightless species of Scarites s. str. inhabiting Florida and the West Indies (Coleoptera: Carabidae:Scaritini). Proc. Entomol. Soc. Wash. 1986, 88, 257–264. [Google Scholar]
- Yang, M.; Hu, B.; Zhou, L.; Liu, X.; Shi, Y.; Song, L.; Wei, Y.; Cao, J. First mitochondrial genome from Yponomeutidae (Lepidoptera, Yponomeutoidea) and the phylogenetic analysis for Lepidoptera. ZooKeys 2019, 879, 137–156. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-P.; Zhao, J.-L.; Su, T.-J.; Luo, A.-R.; Zhu, C.-D. The complete mitochondrial genome of Choristoneura longicellana (Lepidoptera: Tortricidae) and phylogenetic analysis of Lepidoptera. Gene 2016, 591, 161–176. [Google Scholar] [CrossRef]
- Galian, J.; de la Rua, P.; Serrano, J.; Juan, C.; Hewitt, G.M. Phylogenetic relationships in West Mediterranean Scaritina (Coleoptera: Carabidae) inferred from mitochondrial COI sequences and karyotype analysis. J. Zool. Syst. Evol. Res. 1999, 37, 85–92. [Google Scholar] [CrossRef]
- Somvanshi, V.S.; Lang, E.; Sträubler, B.; Spröer, C.; Schumann, P.; Ganguly, S.; Saxena, A.K.; Stackebrandt, E. Providencia vermicola sp. nov., isolated from infective juveniles of the entomopathogenic nematode Steinernema thermophilum. Int. J. Syst. Evol. Microbiol. 2006, 56, 629–633. [Google Scholar] [CrossRef] [Green Version]
- Juneja, P.; Lazzaro, B.P. Providencia sneebia sp. nov. and Providencia burhodogranariea sp. nov., isolated from wild Drosophila melanogaster. Int. J. Syst. Evol. Microbiol. 2009, 59, 1108–1111. [Google Scholar] [CrossRef] [Green Version]
- O’Hara, C.M.; Brenner, F.W.; Miller, J.M. Classification, identification, and clinical significance of Proteus, Providencia, and Morganella. Clin. Microbiol. Rev. 2000, 13, 534–546. [Google Scholar] [CrossRef]
- Benedetti, P.; Rassu, M.; Pavan, G.; Sefton, A.; Pellizzer, G. Septic shock, pneumonia, and soft tissue infection due to Myroides odoratimimus: Report of a case and review of Myroides infections. Infection 2011, 39, 161–165. [Google Scholar] [CrossRef]
- Ravindran, C.; Vasudevan, L.; Kannan, S.R.B.V. Disease symptoms and growth performance of grey mullet infected withMyroides odoratimimus. Aquac. Res. 2016, 48, 5122–5125. [Google Scholar] [CrossRef]
- Pompilio, A.; Galardi, G.; Gherardi, G.; Verginelli, F.; Geminiani, C.; Pilloni, A.P.; Catalanotti, P.; Di Bonaventura, G. Infection of recurrent calcaneal ulcer caused by a biofilm-producer Myroides odoratimimus strain. Folia Microbiol. 2017, 63, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Dharne, M.; Gupta, A.; Rangrez, A.; Ghate, H.; Patole, M.; Shouche, Y. Antibacterial activities of multi drug resistant Myroides odoratimimus bacteria isolated from adult flesh flies (Diptera: Sarcophagidae) are independent of metallo beta-lactamase gene. Braz. J. Microbiol. 2008, 39, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Deguenon, J.M.; Travanty, N.; Zhu, J.; Carr, A.; Denning, S.; Reiskind, M.; Watson, D.W.; Roe, R.M.; Ponnusamy, L. Exogenous and endogenous microbiomes of wild-caught Phormia regina (Diptera: Calliphoridae) flies from a suburban farm by 16S rRNA gene sequencing. Sci. Rep. 2019, 9, 20365. [Google Scholar] [CrossRef] [PubMed]
- Ballinger, M.J.; Perlman, S.J. The defensive Spiroplasma. Curr. Opin. Insect Sci. 2019, 32, 36–41. [Google Scholar] [CrossRef] [Green Version]
- Harumoto, T.; Lemaitre, B. Male-killing toxin in a bacterial symbiont of Drosophila. Nature 2018, 557, 252–255. [Google Scholar] [CrossRef]
- Weng, Y.; Francoeur, C.B.; Currie, C.R.; Kavanaugh, D.H.; Schoville, S.D. A high-quality carabid genome assembly provides insights into beetle genome evolution and cold adaptation. Mol. Ecol. Resour. 2021, 21, 2145–2165. [Google Scholar] [CrossRef]
- Colson, P.; De Lamballerie, X.; Yutin, N.; Asgari, S.; Bigot, Y.; Bideshi, D.K.; Cheng, X.-W.; Federici, B.A.; Van Etten, J.L.; Koonin, E.V.; et al. “Megavirales”, a proposed new order for eukaryotic nucleocytoplasmic large DNA viruses. Arch. Virol. 2013, 158, 2517–2521. [Google Scholar] [CrossRef]
- Williams, T.; Bergoin, M.; van Oers, M.M. Diversity of large DNA viruses of invertebrates. J. Invertebr. Pathol. 2016, 147, 4–22. [Google Scholar] [CrossRef]
- Borisenko, A.V.; Sones, J.E.; Hebert, P.D.N. The front-end logistics of DNA barcoding: Challenges and prospects. Mol. Ecol. Resour. 2009, 9, 27–34. [Google Scholar] [CrossRef]
- Gilbert, M.T.P.; Moore, W.; Melchior, L.; Worobey, M. DNA Extraction from Dry Museum Beetles without Conferring External Morphological Damage. PLoS ONE 2007, 2, e272. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kyndt, E.C.; Kyndt, J.A. Illumina Short-Read Sequencing of the Mitogenomes of Novel Scarites subterraneus Isolates Allows for Taxonomic Refinement of the Genus Scarites Fabricius 1775, within the Carabidae Family. Insects 2022, 13, 190. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020190
Kyndt EC, Kyndt JA. Illumina Short-Read Sequencing of the Mitogenomes of Novel Scarites subterraneus Isolates Allows for Taxonomic Refinement of the Genus Scarites Fabricius 1775, within the Carabidae Family. Insects. 2022; 13(2):190. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020190
Chicago/Turabian StyleKyndt, Elliot C., and John A. Kyndt. 2022. "Illumina Short-Read Sequencing of the Mitogenomes of Novel Scarites subterraneus Isolates Allows for Taxonomic Refinement of the Genus Scarites Fabricius 1775, within the Carabidae Family" Insects 13, no. 2: 190. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020190