
Invasive Populations of the Emerald Ash Borer Agrilus planipennis Fairmaire, 1888 (Coleoptera: Buprestidae) in Saint Petersburg, Russia: A Hitchhiker?


 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
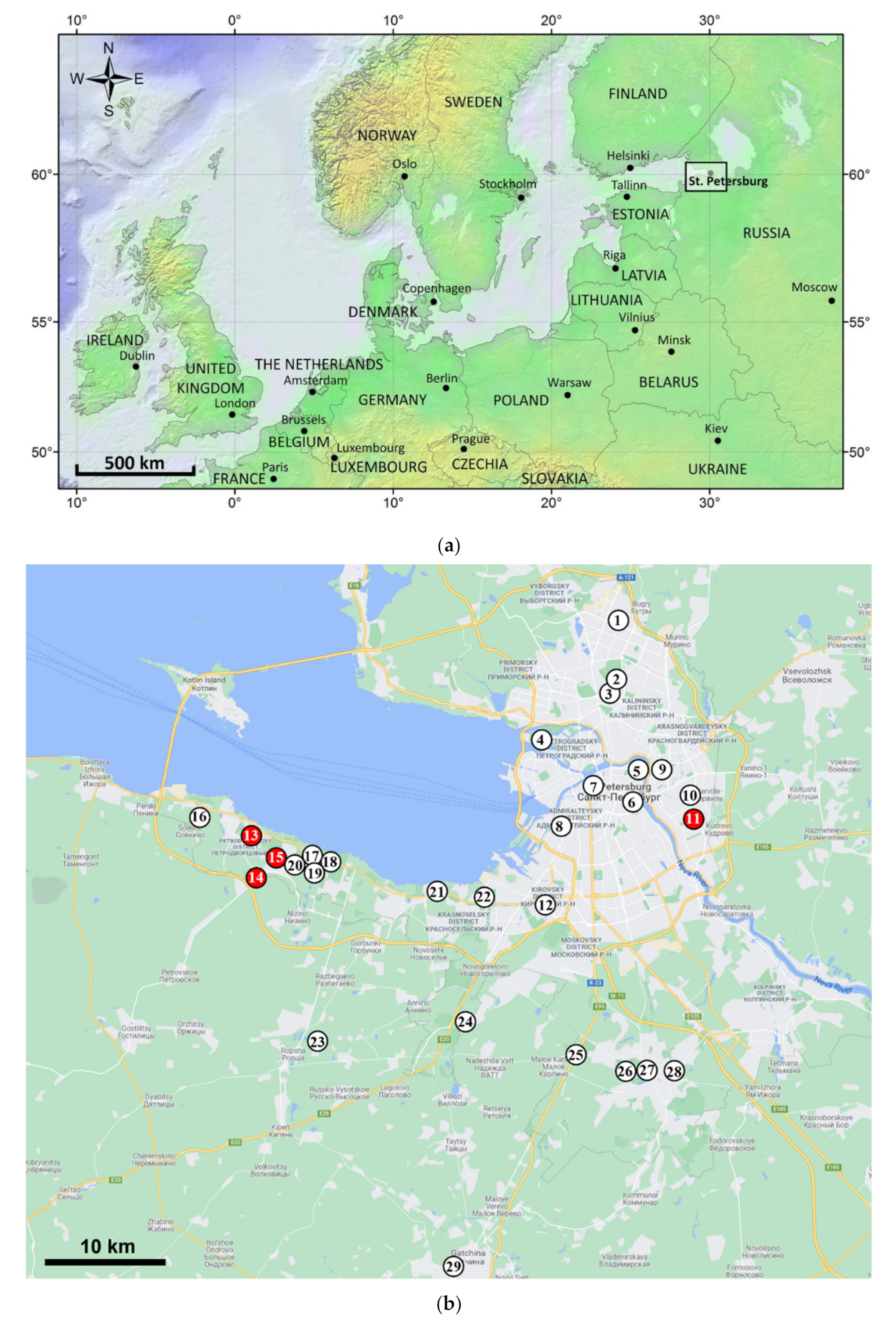
2.1. Survey
2.2. Dating Infestations of EAB
2.3. Examination of Sample Trees
2.4. Analysis of Temperatures
2.5. Statistical Analyses
3. Results
3.1. The Occurrence of EAB in Saint Petersburg
3.2. Ecology of a Local Population of EAB in the Nevsky District
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fyodorova, N.B. Trees and shrubs in Saint Petersburg and monitoring of their health status. Mosc. State For. Univ. Bull.–Lesn. Vestn. 2009, 5, 202–206. (In Russian) [Google Scholar]
- Fyodorova, N.B.; Osypov, D.V.; Grigoriev, A.S. Results of monitoring of communal trees and shrubs in Saint Petersburg in 2007. In Environmental Protection, Nature Management and Environmental Safety in Saint Petersburg in 2007; Golubeva, D.A., Sorokina, N.D., Eds.; Sesame-Print: Saint Petersburg, Russia, 2008; pp. 347–355. (In Russian) [Google Scholar]
- Herms, D.A.; McCullough, D.G. Emerald ash borer invasion of North America: History, biology, ecology, impacts, and management. Annu. Rev. Entomol. 2014, 59, 13–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orlova-Bienkowskaja, M.J.; Drogvalenko, A.N.; Zabaluev, I.A.; Sazhnev, A.S.; Peregudova, E.Y.; Mazurov, S.G.; Komarov, E.V.; Struchaev, V.V.; Martynov, V.V.; Nikulina, T.V.; et al. Current range of Agrilus planipennis Fairmaire, an alien pest of ash trees, in European Russia and Ukraine. Ann. For. Sci. 2020, 77, 29. [Google Scholar] [CrossRef]
- Volkovitsh, M.G.; Bieńkowski, A.O.; Orlova-Bienkowskaja, M.J. Emerald ash borer approaches the borders of the European union and Kazakhstan and is confirmed to infest European ash. Forests 2021, 12, 691. [Google Scholar] [CrossRef]
- Volkovitsh, M.G.; Suslov, D.V. The first record of the emerald ash borer, Agrilus planipennis Fairmaire (Coleoptera: Buprestidae), in Saint Petersburg signals a real threat to the palace and park ensembles of Peterhof and Oranienbaum. In Dendrobiotic Invertebrates and Fungi and their Role in Forest Ecosystems. The Kataev Memorial Readings—XI. In Proceedings of the All-Russia Conference with International Participation, Saint Petersburg, Russia, 24–27 November 2020; Musolin, D.L., Kirichenko, N.I., Selikhovkin, A.V., Eds.; Saint Petersburg State Forest Technical University: Saint Petersburg, Russia, 2020; pp. 121–122. [Google Scholar]
- Musolin, D.L.; Selikhovkin, A.V.; Peregudova, E.Y.; Popovichev, B.G.; Mandelshtam, M.Y.; Baranchikov, Y.N.; Vasaitis, R. North-westward expansion of the invasive range of emerald ash borer, Agrilus planipennis Fairmaire (Coleoptera: Buprestidae) towards the EU: From Moscow to Saint Petersburg. Forests 2021, 12, 502. [Google Scholar] [CrossRef]
- Selikhovkin, A.V.; Popovichev, B.G.; Mandelshtam, M.Y.; Vasaitis, R.; Musolin, D.L. The frontline of invasion: The current northern limit of the invasive range of Emerald ash borer, Agrilus planipennis, in European Russia. Balt. For. 2017, 23, 309–315. Available online: https://www.balticforestry.mi.lt/bf/index.php?option=com_content&view=article&catid=14&id=482 (accessed on 31 August 2021).
- Schowalter, D.N.; Saville, R.J.; Orton, E.S.; Buggs, R.J.A.; Bonello, P.; Brown, J.K.M. Resistance of European ash (Fraxinus excelsior) saplings to larval feeding by the emerald ash borer (Agrilus planipennis). Plants People Planet 2020, 2, 41–46. [Google Scholar] [CrossRef]
- Baranchikov, Y.N.; Seraya, L.G.; Grinash, M.N. All European ash species are susceptible to emerald ash borer Agrilus planipennis Fairmaire (Coleoptera: Buprestidae)—A Far Eastern invader. Sib. For. J. 2014, 6, 80–85, (In Russian with English Summary). [Google Scholar]
- Musolin, D.L.; Selikhovkin, A.V.; Shabunin, D.A.; Zviagintsev, V.B.; Baranchikov, Y.N. Between ash dieback and Emerald ash borer: Two Asian invaders in Russia and the future of ash in Europe. Balt. For. 2017, 23, 316–333. Available online: https://www.balticforestry.mi.lt/bf/PDF_Articles/2017-23%5B1%5D/Baltic%20Forestry%202017.1_316-333.pdf (accessed on 31 August 2021).
- Baranchikov, Y.N. Dating of the beginning of the Agrilus planipennis Fairmaire (Coleoptera: Buprestidae) invasion to St. Petersburg using internet technology. In Dendrobiotic Invertebrates and Fungi and their Role in Forest Ecosystems. The Kataev Memorial Readings—XI. Proceedings of the All-Russia Conference with International Participation, Saint Petersburg, Russia, 24–27 November 2020; Musolin, D.L., Kirichenko, N.I., Selikhovkin, A.V., Eds.; Saint Petersburg State Forest Technical University: Saint Petersburg, Russia, 2020; pp. 70–71. [Google Scholar]
- Gninenko, Y.I.; Klyukin, M.S.; Khegay, I.V. Distribution speed of emerald ash borer in Russia. In Emerald Ash Borer—Distribution and Protection Measures in the USA and Russia; Gninenko, Y.I., Ed.; VNIILM: Pushkino, Russia, 2016; pp. 57–62. (In Russian) [Google Scholar]
- Orlova-Bienkowskaja, M.J.; Bieńkowski, A.O. Minimum winter temperature as a limiting factor of the potential spread of Agrilus planipennis, an alien pest of ash trees, in Europe. Insects 2020, 11, 258. [Google Scholar] [CrossRef] [PubMed]
- Specialized Datasets for Climatic Studies. Available online: http://aisori-m.meteo.ru (accessed on 31 August 2021).
- Økland, B.; Flø, D.; Schroeder, M.; Zach, P.; Cocos, D.; Martikainen, P.; Siitonen, J.; Mandelshtam, M.Y.; Musolin, D.L.; Neuvonen, S.; et al. Range expansion of the small spruce bark beetle Ips amitinus: A newcomer in northern Europe. Agric. For. Entomol. 2019, 21, 286–298. [Google Scholar] [CrossRef] [Green Version]
- Baranchikov, Y.; Mozolevskaya, E.; Yurchenko, G.; Kenis, M. Occurrence of the emerald ash borer, Agrilus planipennis in Russia and its potential impact on European forestry. EPPO Bull. 2008, 38, 233–238. [Google Scholar] [CrossRef]
- Vasaitis, R.; Enderle, R. (Eds.) Dieback of European Ash (Fraxinus spp.)—Consequences and Guidelines for Sustainable Management; Swedish University of Agricultural Sciences: Uppsala, Sweden, 2017; p. 320. Available online: http://www.slu.se/globalassets/ew/org/inst/mykopat/forskning/stenlid/dieback-of-european-ash.pdf (accessed on 31 August 2021).
- Menkis, A.; Bakys, R.; Åslund, M.S.; Davydenko, K.; Elfstrand, M.; Stenlid, J.; Vasaitis, R. Identifying Fraxinus excelsior tolerant to ash dieback: Visual field monitoring versus a molecular marker. For. Pathol. 2019, 50, e12572. [Google Scholar] [CrossRef] [Green Version]
- Enderle, R.; Stenlid, J.; Vasaitis, R. An overview of ash (Fraxinus spp.) and ash dieback disease in Europe. CAB Rev. 2019, 14, 025. [Google Scholar] [CrossRef] [Green Version]
- Semizer-Cuming, D.; Krutovsky, K.V.; Baranchikov, Y.N.; Kjӕr, E.D.; Williams, S.G. Saving the world’s ash forests calls for international cooperation now. Nat. Ecol. Evol. 2019, 3, 141–144. [Google Scholar] [CrossRef] [PubMed]
- McKinney, L.V.; Nielsen, L.R.; Collinge, D.B.; Thomsen, I.M.; Hansen, J.K.; Kjær, E.D. The ash dieback crisis: Genetic variation in resistance can prove a long-term solution. Plant Pathol. 2014, 63, 485–499. [Google Scholar] [CrossRef]
- Evans, H.F.; Williams, D.; Hoch, G.; Loomans, A.; Marzano, M. Developing a European Toolbox to manage potential invasion by emerald ash borer (Agrilus planipennis) and bronze birch borer (Agrilus anxius), important pests of ash and birch. Forestry 2020, 93, 187–196. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| No. | GPS Coordinates of the Locality | Ash Species a | Plantation Category | No. of Investigated Ash Trees | ||
|---|---|---|---|---|---|---|
| Visually from the Ground | Of Those, Felled and Examined in Detail | Attacked by EAB | ||||
| PLANTINGS IN URBAN ENVIRONMENT | ||||||
| 3 | 59°59′34″ N, 30°20′33″ E | F. ex. | city park | 20 | 0 | 0 |
| 4 | 59°58′16″ N, 30°14′48″ E | F. ex. | city park | 6 | 2 | 0 |
| 5 | 59°56′41″ N, 30°22′49″ E | F. ex. | square | 1 | 1 | 0 |
| 6 | 59°55′51″ N, 30°22′36″ E | F. ex. | square | 5 | 1 | 0 |
| 7 | 59°54′49″ N, 30°17′17″ E | F. ex. | square | 4 | 1 | 0 |
| 8 | 59°54′13″ N, 30°15′39″ E | F. ex. | city park | 15 | 0 | 0 |
| 10 | 59°51′59″ N, 30°21′50″ E | F. ex. | city park | 6 | 1 | 0 |
| 22 | 59°50′48″ N, 30°08′54″ E | F. ex. | city park | 15 | 0 | 0 |
| 26 | 59°43′23″ N, 30°23′41″ E | F. ex. | city park | 20 | 0 | 0 |
| 27 | 59°43′04″ N, 30°22′11″ E | F. ex. | city park | 30 | 4 | 0 b |
| 28 | 59°43′10″ N, 30°25′26″ E | F. ex. | alley | 25 | 0 | 0 |
| Total F. ex. | 147 | 10 | 0 | |||
| 1 | 60°03′32″ N, 30°20′59″ E | F. p. | street | 10 | 0 | 0 |
| 2 | 60°00′00″ N, 30°21′01″ E | F. p. | street | 3 | 0 | 0 |
| 9.1 c | 59°55′35″ N, 30°24′26″ E | F. p. | square | 1 | 1 | 0 |
| 9.2 | 59°55′58″ N, 30°24′38″ E | F. p. | city park | 16 | 1 | 0 |
| 12 | 59°50′47″ N, 30°15′32″ E | F. p. | street | 4 | 1 | 0 |
| 24 | 59°43′56″ N, 30°05′32″ E | F. p. | city park | 10 | 3 | 0 |
| 25 | 59°43′50″ N, 30°17′35″ E | F. p. | roadside | 40 | 3 | 0 |
| Total F. p. | 84 | 9 | 0 | |||
| EAB invasion, Nevsky District | ||||||
| 11.1 c | 59°54′31″ N, 30°27′43″ E | F. p. | city park | 28 | 19 | 19 |
| 11.2 | 59°54′38″ N, 30°27′59″ E | F. ex. | alley | 35 | 8 | 8 |
| EAB invasion, Petrodvortsovy District | ||||||
| 13.1 c | 59°54′08″ N, 29°49′02″ E | F. p. | street | 63 | 63 | 63 |
| 13.2 | 59°54′08″ N, 29°49′02″ E | F. ex. | street | 3 | 1 | 1 |
| 14 | 59°51′49″ N, 29°48′38″ E | F. p. | roadside | 15 | 15 | 15 |
| 15.1 c | 59°53′01″ N, 29°52′00″ E | F. p. | square | 20 | 0 | 4 |
| 15.2 | 59°53′36″ N, 29°51′54″ E | F. p. | roadside | 12 | 0 | 1 |
| Total F. p. + F. ex. | 110 + 3 | 78 + 1 | 83 + 1 | |||
| PETERHOF STATE MUSEUM RESERVE | ||||||
| 16 | 59°54′47″ N, 29°44′36″ E | F. ex. | park | 600 | 0 | 0 |
| 17 | 59°53′12″ N, 29°54′30″ E | F. ex. | park | 1500 | 4 | 0 b |
| 18 | 59°52′52″ N, 29°56′22″ E | F. ex. | park | 275 | 0 | 0 |
| 19 | 59°52′37″ N, 29°54′29″ E | F. ex. + F. p. | park | 7 + 2 | 1 + 0 | 0 |
| 20 | 59°53′33″ N, 29°51′59″ E | F. ex. | park | 25 | 0 | 0 |
| 21 | 59°51′13″ N, 30°02′46″ E | F. ex. + F. p. | park | 2 + 1 | 0 | 0 |
| Total F. ex. + F. p. | 2409 + 3 | 5 | 0 | |||
| GATCHINA STATE MUSEUM RESERVE | ||||||
| 23 | 59°43′24″ N, 29°51′31″ E | F. ex. | park | 300 | 12 | 0 b |
| 29 | 59°33′47″ N, 30°06′52″ E | F. ex. | park | 250 | 35 | 0 b |
| Total F. ex. | 550 | 47 | 0 | |||
| THROUGHOUT THE STUDY | ||||||
| F. ex., no. (%) | 3144 | 71 | 9 (0.3) | |||
| F. p., no. (%) | 225 | 106 | 102 (45.3) | |||
| ALL, no. (%) | 3369 | 177 | 111 (3.3) | |||
| Tree No./Years Of Infestation | Ash Species a | DBH b (cm) | Height (m) | Colonized Stem Height, Min–Max (m) | Larval Density (no./1 dm2 of Bark) c | Number per Tree | Percent | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Larval Galleries | Exit Holes | Viable Larvae | Emerged Adult Beetles | Emerged Adult Beetles + Viable Larvae d | ||||||
| 1/2015–2018 | F. p. | 16 | 8.5 | 0–4 | 0.34 | 58 | 26 | 0 | 44.8 | 44.8 |
| 1/2019–2020 | + 0 e | 0 | 0 | 0 | 0 | 0 | 0 | |||
| 2/2015–2018 | F. p. | 16 | 10.2 | 0–4 | 0.53 | 92 | 44 | 0 | 47.8 | 47.8 |
| 2/2019–2020 | + 1 e | 0.29 | 50 | 10 | 12 | 0 | 44.0 | |||
| 3/2015–2018 | F. p. | 20 | 12.4 | 0–4 | 0.25 | 64 | 48 | 0 | 75.0 | 75.0 |
| 3/2019–2020 | + 2 e | 0.04 | 10 | 0 | 4 | 0 | 40.0 | |||
| 4/2015–2018 | F. p. | 21 | 9.3 | 0–2 | 0.14 | 30 | 12 | 0 | 40.0 | 40.0 |
| 4/2019–2020 | + 7 e | 0.04 | 32 | 22 | 8 | 0 | 93.8 | |||
| 5/2019–2020 | F. ex. | 11 | 7.2 | 0–6 | 0.29 | 43 | 8 | 26 | 0 | 79.1 |
| 6/2019–2020 | F. ex. | 12 | 6.4 | 0–5 | 0.71 | 156 | 37 | 75 | 0 | 71.8 |
| 7/2019–2020 | F. ex. | 11 | 5.5 | 1–3 | 0.77 | 60 | 4 | 3 | 0 | 11.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Selikhovkin, A.V.; Musolin, D.L.; Popovichev, B.G.; Merkuryev, S.A.; Volkovitsh, M.G.; Vasaitis, R. Invasive Populations of the Emerald Ash Borer Agrilus planipennis Fairmaire, 1888 (Coleoptera: Buprestidae) in Saint Petersburg, Russia: A Hitchhiker? Insects 2022, 13, 191. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020191
Selikhovkin AV, Musolin DL, Popovichev BG, Merkuryev SA, Volkovitsh MG, Vasaitis R. Invasive Populations of the Emerald Ash Borer Agrilus planipennis Fairmaire, 1888 (Coleoptera: Buprestidae) in Saint Petersburg, Russia: A Hitchhiker? Insects. 2022; 13(2):191. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020191
Chicago/Turabian StyleSelikhovkin, Andrey V., Dmitry L. Musolin, Boris G. Popovichev, Sergey A. Merkuryev, Mark G. Volkovitsh, and Rimvys Vasaitis. 2022. "Invasive Populations of the Emerald Ash Borer Agrilus planipennis Fairmaire, 1888 (Coleoptera: Buprestidae) in Saint Petersburg, Russia: A Hitchhiker?" Insects 13, no. 2: 191. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020191