Andricus cydoniae Giraud, 1859 Junior Synonym of Cynips conifica Hartig, 1843, as Experimentally Demonstrated (Hymenoptera: Cynipidae: Cynipini)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Abbreviations Used in the Text
- GCPC: Private collection of Giuliano Cerasa, Giuliana, Palermo, Italy
- SSPC: Private collection of Salvatore Sottile, Cinisello Balsamo, Milan, Italy
- MCLSS: Museo Civico Lentate Sul Seveso, Milan, Italy
2.2. Study Material Used in the Experiments
- (1)
- Asexual females placed in contact chamber (Experiment 1):
- (2)
- Sexual generation obtained from Experiment 1
- (3)
- Sexual generation obtained from Experiment 1 and used in the Experiment 2:
2.3. Additional Material Examined for Morphological Diagnosis
2.4. Laboratory Assays
2.4.1. Experiment 1
2.4.2. Experiment 2
2.5. Morphological Study
3. Results
3.1. Laboratory Assays
3.2. Gall
3.3. Similar Galls
3.4. Diagnosis of the Asexual Form
3.5. Diagnosis of the Sexual Form
3.6. Biology and Host Plant
3.7. Distribution
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stone, G.N.; Schönrogge, K.; Atkinson, R.J.; Bellido, D.; Pujade-Villar, J. The population biology of oak gall wasps (Hymenoptera: Cynipidae). Annu. Rev. Entomol. 2002, 47, 633–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csóka, G.; Stone, G.N.; Melika, G.G. Biology, ecology, and evolution of gall-inducing Cynipidae. In Biology, Ecology, and Evolution of Gall-Inducing Arthropods; Raman, A., Schaefer, C.W., Withers, T.M., Eds.; Science Publishers, Inc.: Enfield, NH, USA, 2005; Volume 2, pp. 573–642. [Google Scholar]
- Avtzis, D.N.; Melika, G.; Matošević, D.; Coyle, D.R. The Asian chestnut gall wasp Dryocosmus kuriphilus: A global invader and a successful case of classical biological control. J. Pest Sci. 2018, 92, 107–115. [Google Scholar] [CrossRef]
- Massa, B.; Cerasa, G.; Pagliano, G. Insecta Hymenoptera Cynipidae. In Checklist of the Italian Fauna; Version 1.0; LifeWatch Italia: Lecce, Italy, 2021; Available online: https://www.lifewatchitaly.eu/en/initiatives/checklist-fauna-italia-en/checklist/ (accessed on 31 May 2021).
- Pujade-Villar, J.; Bellido, D.; Segú, G.; Melika, G. Current state of knowledge of heterogony in Cynipidae (Hymenoptera: Cynipoidea). Sess. Conjunta Entomol. 2001, 11, 87–107. [Google Scholar]
- Walker, P. Andricus aries does have a sexual generation on Turkey oak. Cecidology 2001, 16, 94–95. [Google Scholar]
- Walker, P. Two new records for cynipid oak galls (Cynipidae: Hymenoptera) in Britain. Cecidology 2002, 17, 64–67. [Google Scholar]
- Pujade i Villar, J.; Folliot, R.; Bellido, D. The life cycle of Andricus hispanicus (Hartig, 1856) n. stat., a sibling species of A. kollari (Hartig, 1843) (Hymenoptera: Cynipidae). Butll. Inst. Cat. Hist. Nat. 2004, 83–95. Available online: https://raco.cat/index.php/ButlletiICHN/article/view/235821 (accessed on 31 May 2021).
- Folliot, R.; Ros-Farré, P.; Bellido, D.; Pujade-Villar, J. Alternation of generations in Andricus corruptrix (Schlechtendal): Comments on and description of a new sexual form (Hymenoptera: Cynipidae). Contrib. Zool. 2004, 73, 263–270. [Google Scholar] [CrossRef] [Green Version]
- Folliot, R.; Pujade-Villar, J. Males of Andricus hystrix Trotter, a new sexual form of Cynipidae (Hymenoptera). Bol. SEA 2006, 38, 157–160. [Google Scholar]
- Stone, G.N.; Atkinson, R.J.; Rokas, A.; Aldrey, J.-L.N.; Melika, G.; Ács, Z.; Csóka, G.; Hayward, A.; Bailey, R.; Buckee, C.; et al. Evidence for widespread cryptic sexual generations in apparently purely asexual Andricus gallwasps. Mol. Ecol. 2008, 17, 652–665. [Google Scholar] [CrossRef]
- Doutt, R.L. Heterogony in Dryocosmus (Hymenoptera, Cynipidae). Ann. Entomol. Soc. Am. 1959, 52, 69–74. [Google Scholar] [CrossRef]
- Nicholls, J.A.; Melika, G.; DeMartini, J.; Stone, G.N. New Species of Dryocosmus Giraud gallwasps from California (Hymenoptera: Cynipidae: Cynipini) galling Chrysolepis Hjelmq. (Fagaceae). Zootaxa 2018, 4532, 407–433. [Google Scholar] [CrossRef] [PubMed]
- Cerasa, G.; Lo Verde, G.; Caleca, V.; Massa, B.; Nicholls, J.A.; Melika, G. Description of the sexual generation of Dryocosmus destefanii (Hymenoptera: Cynipidae: Cynipini) and disclosure of its life cycle. Zootaxa 2020, 4742, 359–374. [Google Scholar] [CrossRef] [PubMed]
- Hartig, T. Zweiter nachtrag zur naturgeschichte der gallwespen. Z. Entomol. 1843, 4, 395–422. [Google Scholar]
- Rohwer, S.A.; Fagan, M.M. The type-species of the Genera of the Cynipoidea, or the gall wasps and parasitic Cynipoids. Proc. U. S. Natl. Mus. 1917, 53, 357–380. [Google Scholar] [CrossRef]
- Marsden-Jones, E.M. A Study of the life-cycle of Adleria kollari Hartig, the marble or devonshire gall. Trans. R. Entomol. Soc. Lond. 1953, 104, 195–221. [Google Scholar] [CrossRef]
- Giraud, J.E. Signalements de quelques espèces nouvelles de cynipides et de leurs galles. Verh. Zool.-Bot. Ver. Wien 1859, 9, 337–374. [Google Scholar]
- Cook, J.M.; Rokas, A.; Pagel, M.; Stone, G.N. Evolutionary shifts between host oak sections and host-plant organs in Andricus gallwasps. Evolution 2002, 56, 1821–1830. [Google Scholar] [CrossRef]
- Melika, G. Gall Wasps of Ukraine; Vestnik Zoologii, Schmalhausen Institute of Zoology: Kyiv, Ukraine, 2006; Volume 1–2, pp. 1–644. [Google Scholar]
- Dalla-Torre, K.W.; Kieffer, J.J. Cynipidae; Das Tierreich, 24; Friedlander & Sohn: Berlin, Germany, 1910; pp. 1–891. Available online: https://www.biodiversitylibrary.org/bibliography/1077 (accessed on 8 February 2022).
- Hadley, A. Combine ZP. 2011. Available online: http://hadleyweb.pwp.blueyonder.co.uk (accessed on 12 February 2015).
- Fergusson, N.D.M. The Cynipoid Families. In The Hymenoptera of Costa Rica; Hanson, P.E., Gauld, I.D., Eds.; Oxford University Press: Oxford, UK, 1995; pp. 247–265. [Google Scholar]
- Liljeblad, J.; Ronquist, F. A phylogenetic analysis of higher-level gall wasp relationships (Hymenoptera: Cynipidae). Syst. Entomol. 1998, 23, 229–252. [Google Scholar] [CrossRef]
- Melika, G.; Pujade-Villar, J.; Abe, Y.; Tang, C.-T.; Nicholls, J.; Wachi, N.; Ide, T.; Yang, M.-M.; Pénzes, Z.; Csóka, G. Palaearctic oak gallwasps galling oaks (Quercus) in the section Cerris: Re-Appraisal of generic limits, with descriptions of new genera and species (Hymenoptera: Cynipidae: Cynipini). Zootaxa 2010, 2470, 1. [Google Scholar] [CrossRef]
- Polidori, C.; Nieves-Aldrey, J.L. Diverse filters to sense: Great variability of antennal morphology and sensillar equipment in gall-wasps (Hymenoptera: Cynipidae). PLoS ONE 2014, 9, e101843. [Google Scholar] [CrossRef]
- Ronquist, F.; Nordlander, G. Skeletal Morphology of an Archaic Cynipoid, Ibalia rufipes (Hymenoptera: Ibaliidae). Entomol. Scandinavica. Suppl. 1989, 1–60. Available online: https://agris.fao.org/agris-search/search.do?recordID=SE8911554 (accessed on 31 May 2021).
- Harris, R.A. A glossary of surface sculpturing. Occas. Pap. Entomol. 1979, 28, 1–31. [Google Scholar]
- Eady, R.D. Some illustrations of microsculpture in the Hymenoptera. Roy. Ent. Soc. Lond. Proc. Ser. A Gen. Ent. 1968, 43, 66–72. [Google Scholar] [CrossRef]
- Yoder, M.J.; Mikó, I.; Seltmann, K.C.; Bertone, M.A.; Deans, A.R. A gross anatomy ontology for Hymenoptera. PLoS ONE 2010, 5, e15991. [Google Scholar] [CrossRef] [PubMed]
- Seltmann, K.; Yoder, M.; Miko, I.; Forshage, M.; Bertone, M.; Agosti, D.; Austin, A.; Balhoff, J.; Borowiec, M.; Brady, S.; et al. A Hymenopterists’ guide to the Hymenoptera anatomy ontology: Utility, clarification, and future directions. J. Hymenopt. Res. 2012, 27, 67–88. [Google Scholar] [CrossRef] [Green Version]
- ICZN. International Commission on Zoological Nomenclature. In International Code of Zoological Nomenclature, 4th ed.; The International Trust for Zoological Nomenclature: London, UK, 1999; pp. 1–305. Available online: https://www.iczn.org/assets/63c4277759/Updates-to-print-Code4.pdf (accessed on 31 May 2021).
- Uechi, N.; Yukawa, J.; Tokuda, M.; Maryana, N.; Ganaha-Kikumura, T.; Kim, W. Description of the Asian chili pod gall midge, Asphondylia capsicicola sp. n., with comparative notes on Asphondylia gennadii (Diptera: Cecidomyiidae) that induces the same sort of pod gall on the same host plant species in the Mediterranean region. Appl. Entomol. Zool. 2017, 52, 113–123. [Google Scholar] [CrossRef]
- Abe, Y.; Ide, T.; Su, C.-Y.; Zhu, D.-H. Leaf galls with the same morphology induced on the same plant species by two species of Latuspina (Hymenoptera: Cynipidae), with a description of a new species. Proc. Entomol. Soc. Wash. 2021, 123, 465–473. [Google Scholar] [CrossRef]
- Pujade, J.; Tavakoli, M.; Melika, G.; Ferrer Suay, M. Andricus synophri (Hymenoptera: Cynipidae), a New species of oak gallwasp from Iran. J. Insect Biodivers. Syst. 2015, 1, 1–10. [Google Scholar]
- Meier, F.; Engesser, R.; Forster, B.; Odermatt, O.; Angst, A.; Hölling, D. Forstschutz-Überblick 2014. WSL Berichte 2015, 23. Available online: https://www.dora.lib4ri.ch/wsl/islandora/object/wsl%3A9084/datastream/PDF/view (accessed on 31 May 2021).
- Marković, Č. Contribution to knowledge of the fauna of cynipid gall wasps (Hymenoptera: Cynipidae) of Mt. Jastrebac (Serbia). Acta Entomol. Serbica 2014, 19, 63–72. [Google Scholar]
- Kwast, E. A contribution to the fauna of Cynipidae (Insecta, Hymenoptera, Cynipidae) of Croatia with a description of an asexual female of Andricus korlevici (Kieffer, 1902) Nov. Comb. Nat. Croat. Period. Musei Hist. Nat. Croat. 2012, 21, 223–245. [Google Scholar]
- Tomasi, E. Indagine Cecidologica Dell’isola Di Cres-Lošinj (Cherso-Lussino) (Hrvatska, Adriatic Sea, NE). Atti Mus. Civ. Stor. Nat. Trieste 2019, 60, 153–268. [Google Scholar]
- Kollár, J. Gall-inducing arthropods associated with ornamental woody plants in a city park of Nitra (SW Slovakia). Acta Entomol. Serbica 2011, 16, 115–126. [Google Scholar]
- Katılmış, Y.; Kıyak, S. Checklist of Cynipidae of Turkey with a New Genus Record. J. Nat. Hist. 2008, 42, 2161–2167. [Google Scholar] [CrossRef]
- Katılmış, Y.; Azmaz, M. Investigation on the inquilines (Hymenoptera: Cynipidae, Synergini) of oak galls from inner western Anatolia, Turkey. Turk. J. Zool. 2015, 39, 168–173. [Google Scholar] [CrossRef]
- De Stefani, T. Produzioni patologiche sulle piante causate da animali. Agric. Calabr.-Siculo 1898, 23, 1–12. [Google Scholar]
- Rokas, A.; Melika, G.; Abe, Y.; Nieves-Aldrey, J.-L.; Cook, J.M.; Stone, G.N. Lifecycle closure, lineage sorting, and hybridization revealed in a phylogenetic analysis of european oak gallwasps (Hymenoptera: Cynipidae: Cynipini) using mitochondrial sequence data. Mol. Phylogenet. Evol. 2003, 26, 36–45. [Google Scholar] [CrossRef]
- Hood, G.R.; Zhang, L.; Topper, L.; Brandão-Dias, P.F.; Del Pino, G.A.; Comerford, M.S.; Egan, S.P. ‘Closing the Life Cycle’of Andricus quercuslanigera (Hymenoptera: Cynipidae). Ann. Entomol. Soc. Am. 2018, 111, 103–113. [Google Scholar] [CrossRef]
- Folliot, R. Contribution à l’étude de la biologie des Cynipides gallicoles (Hyménoptères, Cynipoidea). Ann. Sci. Nat. Zool. 1964, 4, 407–564. [Google Scholar]
- Lund, J.N.; Ott, J.R.; Lyon, R.J. Heterogony in Belenocnema treatae Mayr (Hymenoptera: Cynipidae). Proc. Entomol. Soc. Wash. 1998, 100, 755–763. [Google Scholar]
- Garbin, L.; Díaz, N.B.; Pujade-Villar, J. Experimental study of the reproductive cycle of Plagiotrochus amenti Kieffer, 1901 (Hymenoptera, Cynipoidea, Cynipidae), with comments on its taxonomy. Boln. Asoc. Esp. Ent. 2008, 32, 341–349. [Google Scholar]
- Adler, H. Les Cynipidae. 1re Partie. Introdution. La Génération Alternante Chez Les Cynipidae; Traduit et Annoté Par J. Lichtenstein: Paris, France, 1881; pp. 1–141. Available online: https://www.biodiversitylibrary.org/bibliography/11137 (accessed on 31 May 2021).
- Cerasa, G.; Lo Verde, G.; Caleca, V.; Massa, B.; Nicholls, J.A.; Melika, G. Description of Dryocosmus destefanii new species (Hymenoptera: Cynipidae: Cynipini) from Quercus suber L. in Italy. Zootaxa 2018, 4370, 535–548. [Google Scholar] [CrossRef] [PubMed]
- Cerasa, G.; Sottile, S.; Massa, B.; Lo Verde, G. Rediscovery of Cerroneuroterus apenninus (Trotter, 1923) (Hymenoptera: Cynipidae: Cynipini) a valid species from Italy with description and neotype designation. Zootaxa 2021, 4941, 399–414. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Features | A. conificus ♀ (=A. cydoniae) | A. multiplicatus ♀ |
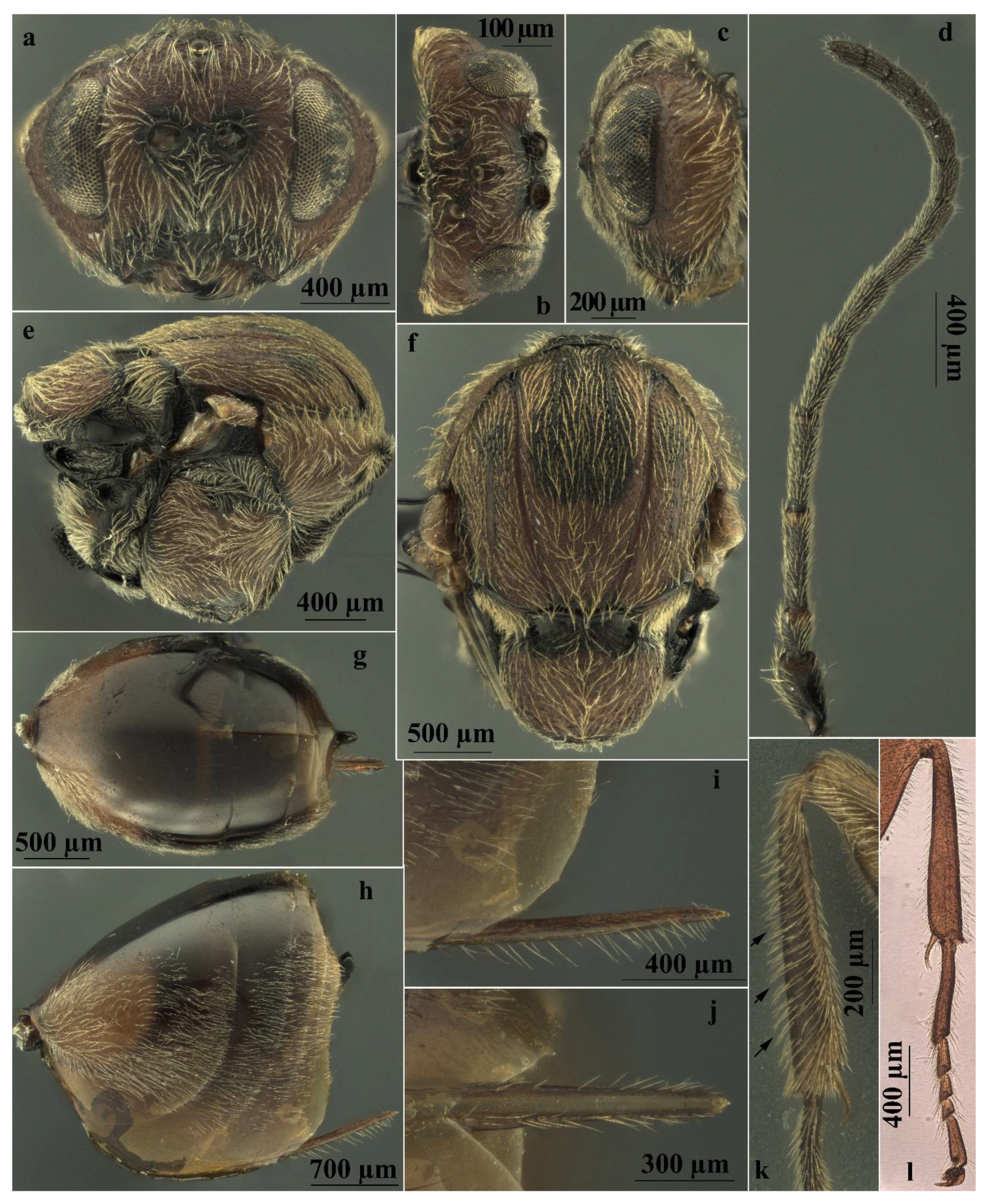
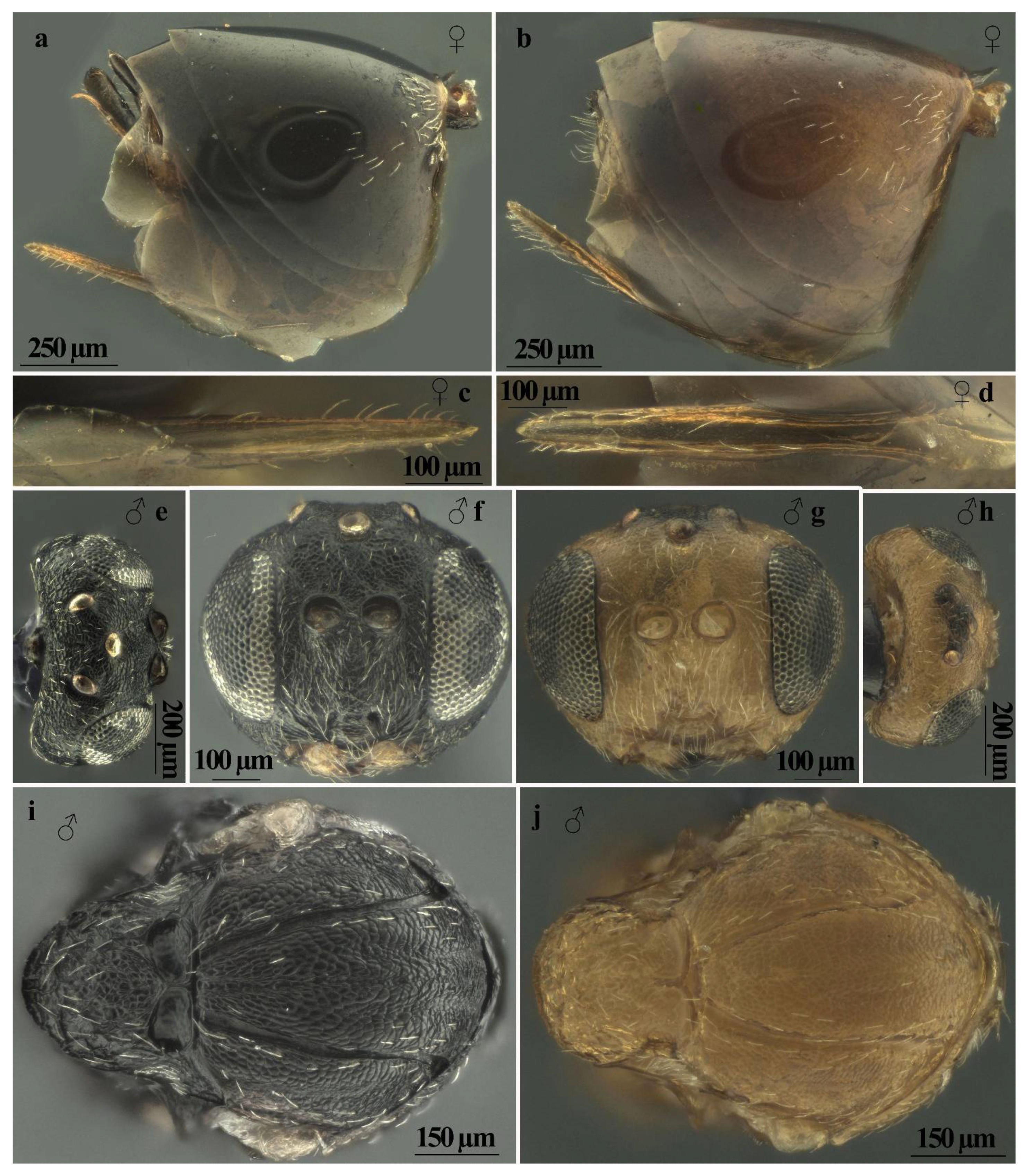
| Body color | Mostly dark brown to black, with yellow legs, except for proximal part of hind coxae being dark brown (Figure 3a) | Mostly reddish yellow, legs slightly lighter than body (Figure 4a) |
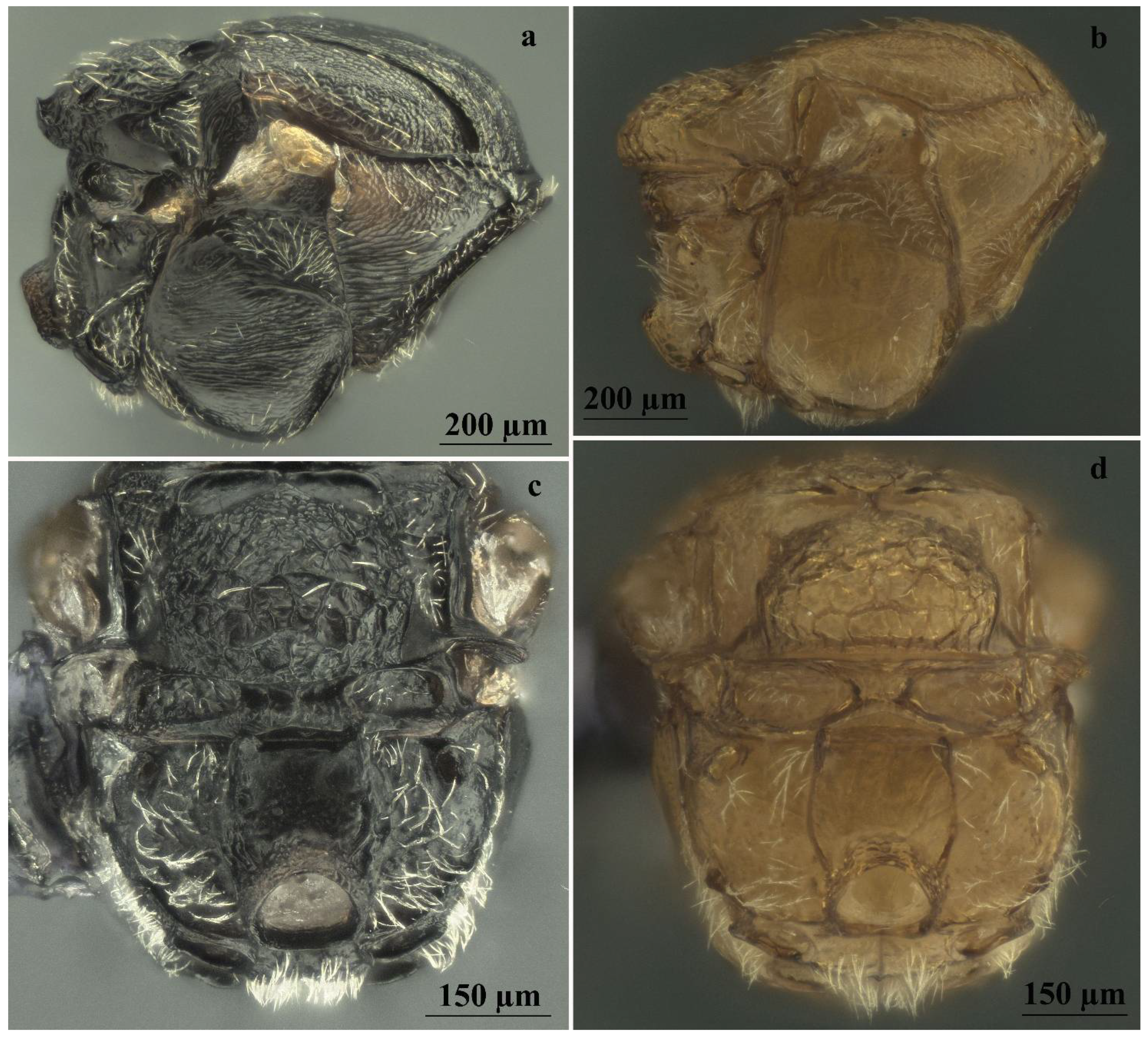
| Head sculpture | Frons, vertex, and occiput reticulate (Figure 6a,b) | Frons, vertex, and occiput uniformly coriaceous (Figure 6c,d) |
| Striae on mesopleuron | With very marked striae (Figure 6e) | With or without very indistinct striae (Figure 6f) |
| Sculpture, shape, and size of mesoscutellum | As long as is broad; uniformly strongly areolate-rugose with distinct mainly longitudinal sharp rugae with emarginate posterior margin (Figure 6e,j) | Broader than long; reticulate rugose around its limits, more delicate in the central part of disk with unemarginate posterior margin (Figure 6f,i) |
| Shape of scutellar foveae | Scutellar foveae subtriangular well-delimited posteriorly (Figure 6g,j) | Subrectangular not or very slightly delimited posteriorly (Figure 6h,i) |
| Mesoscutum sculpture | Deeply colliculate (Figure 6g) | Shallowly colliculate (Figure 6h) |
| White setae on prominent part of ventral spine of hypopygium | Very few, short (approximately as long as the median diameter of the hypopygium in lateral view), erect, and not extending behind apex of ventral spine (Figure 7a, c) | Few, long (about one and a half times the median diameter of the hypopygium in lateral view), curved, and slightly extending behind apex of spine (Figure 7b,d) |
| Features | A. conificus ♂ (=A. cydoniae) | A. multiplicatus ♂ |
| Body color | Mostly dark brown to black, with yellow legs, except for proximal part of hind coxae being dark brown (Figure 3b) | Mostly reddish yellow, legs slightly lighter than body (Figure 4b) |
| Ratio of diameter of torulus (including rims) to eye-torulus distance | Nearly 1.6 times eye-torulus distance (Figure 7f) | Equal to eye-torulus distance (Figure 7g) |
| Ratio of eye-torulus distance to distance between toruli | Nearly 1.4 times as large as distance between toruli (Figure 7f) | Nearly 3.0 times as large as distance between toruli (Figure 7g) |
| Frons and vertex sculpture | Rugose (Figure 7e,f) | Coriaceous (Figure 7g,h) |
| Shape of scutellar foveae | Subtriangular well-delimited posteriorly (Figure 7i) | Subrectangular Not or very slightly delimited posteriorly (Figure 7j) |
| Sculpture, shape, and size of mesoscutellum | Around its limits, strongly reticulate rugose; more delicate or colliculate in the central part of disk, with emarginate posterior margin (Figure 7i) | Around its limits, reticulate rugose; more delicate or coriaceous in the central part of disk, with unemarginate posterior margin (Figure 7j) |
| Mesoscutum sculpture | Deeply colliculate (Figure 7i) | Shallowly colliculate (Figure 7j) |
| Striae on mesopleuron | With very marked striae (Figure 8a) | With or without very indistinct striae (Figure 8b) |
| Ratio of breadth to height of metascutellum | More than 2.0 (Figure 8c) | Less than 1.5 (Figure 8d) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sottile, S.; Cerasa, G.; Massa, B.; Lo Verde, G. Andricus cydoniae Giraud, 1859 Junior Synonym of Cynips conifica Hartig, 1843, as Experimentally Demonstrated (Hymenoptera: Cynipidae: Cynipini). Insects 2022, 13, 200. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020200
Sottile S, Cerasa G, Massa B, Lo Verde G. Andricus cydoniae Giraud, 1859 Junior Synonym of Cynips conifica Hartig, 1843, as Experimentally Demonstrated (Hymenoptera: Cynipidae: Cynipini). Insects. 2022; 13(2):200. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020200
Chicago/Turabian StyleSottile, Salvatore, Giuliano Cerasa, Bruno Massa, and Gabriella Lo Verde. 2022. "Andricus cydoniae Giraud, 1859 Junior Synonym of Cynips conifica Hartig, 1843, as Experimentally Demonstrated (Hymenoptera: Cynipidae: Cynipini)" Insects 13, no. 2: 200. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020200