
DNA Barcoding versus Morphological Variability of Pterostichus brevicornis brevicornis (Kirby, 1837) (Coleoptera, Carabidae) in the Arctic and Subarctic

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Collection, DNA Extraction, PCR, and Sequencing
2.2. Phylogenetic and Phylogeographic Analyses
2.3. Population Genetic Analysis
2.4. Morphological Studies
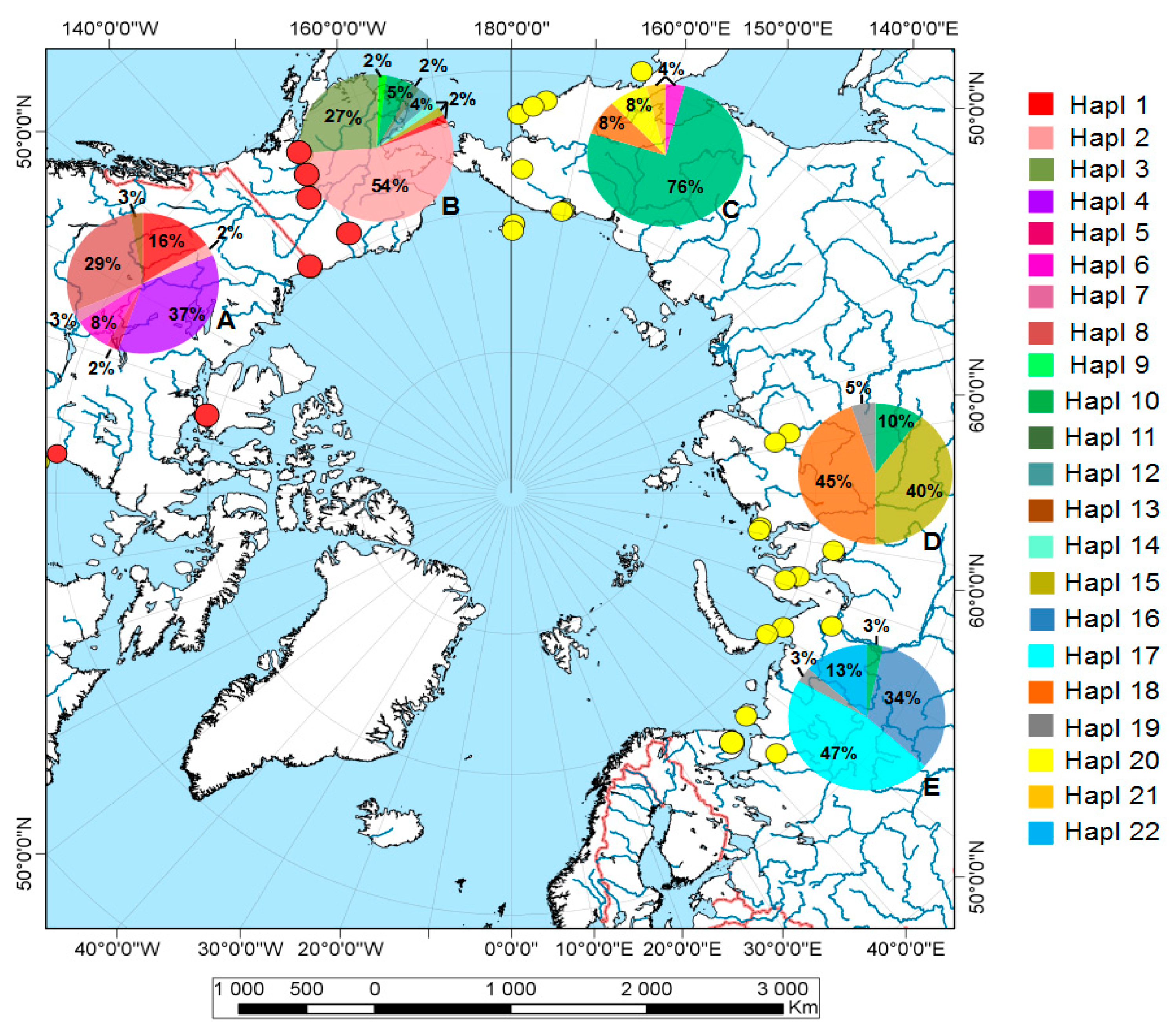
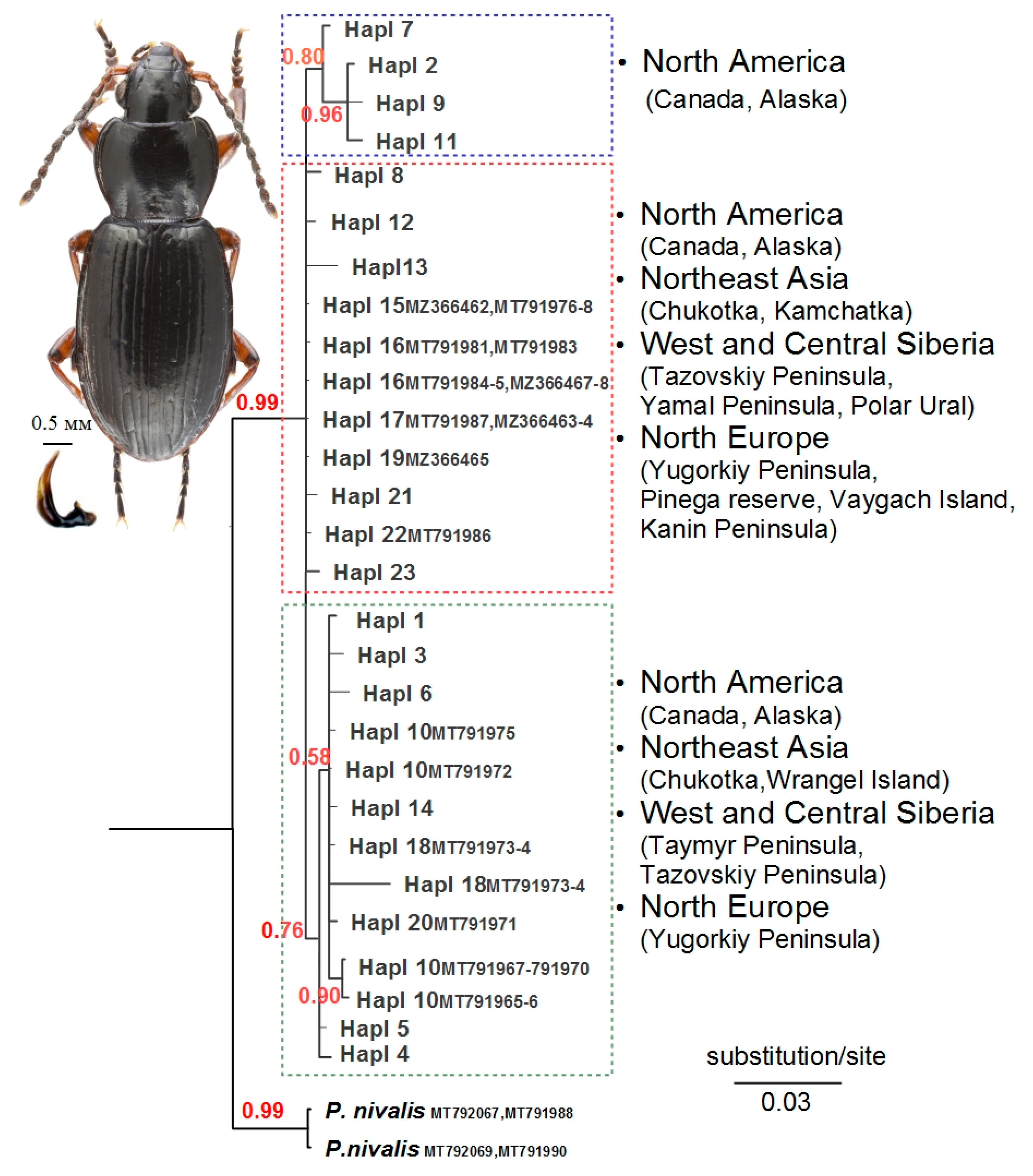
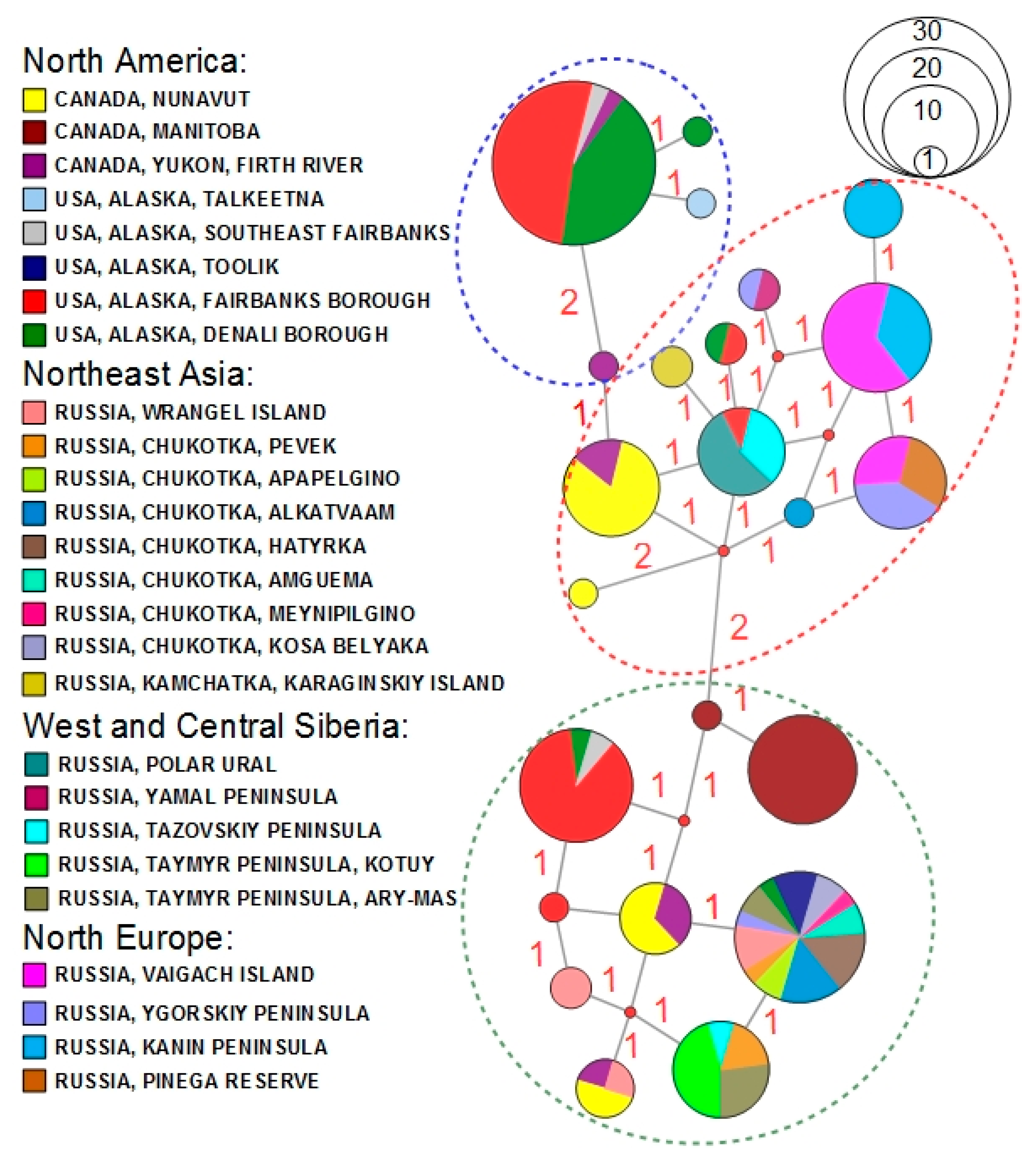
3. Results
3.1. Phylogenetic Reconstruction, Phylogeography, and Population Genetics
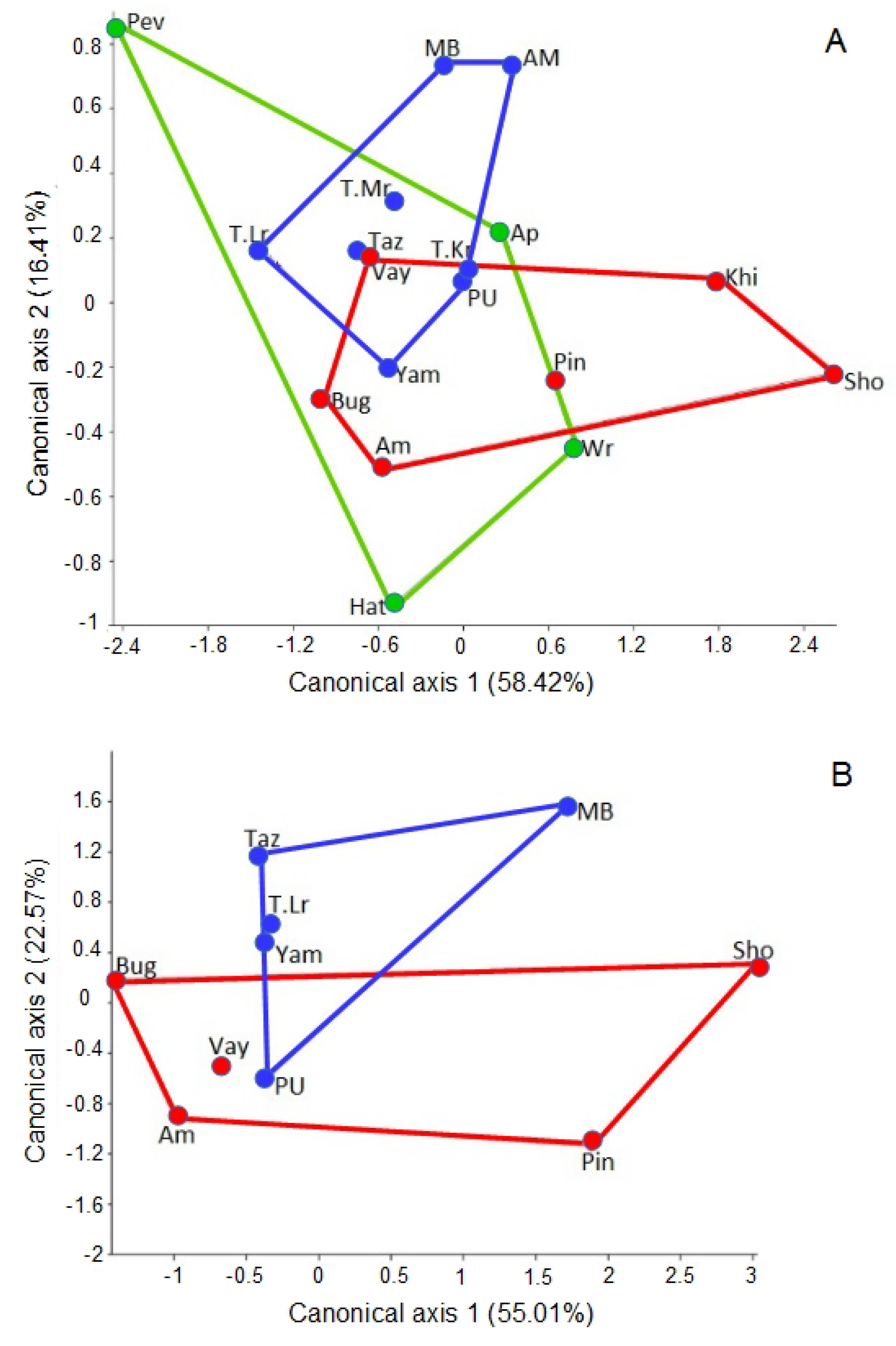
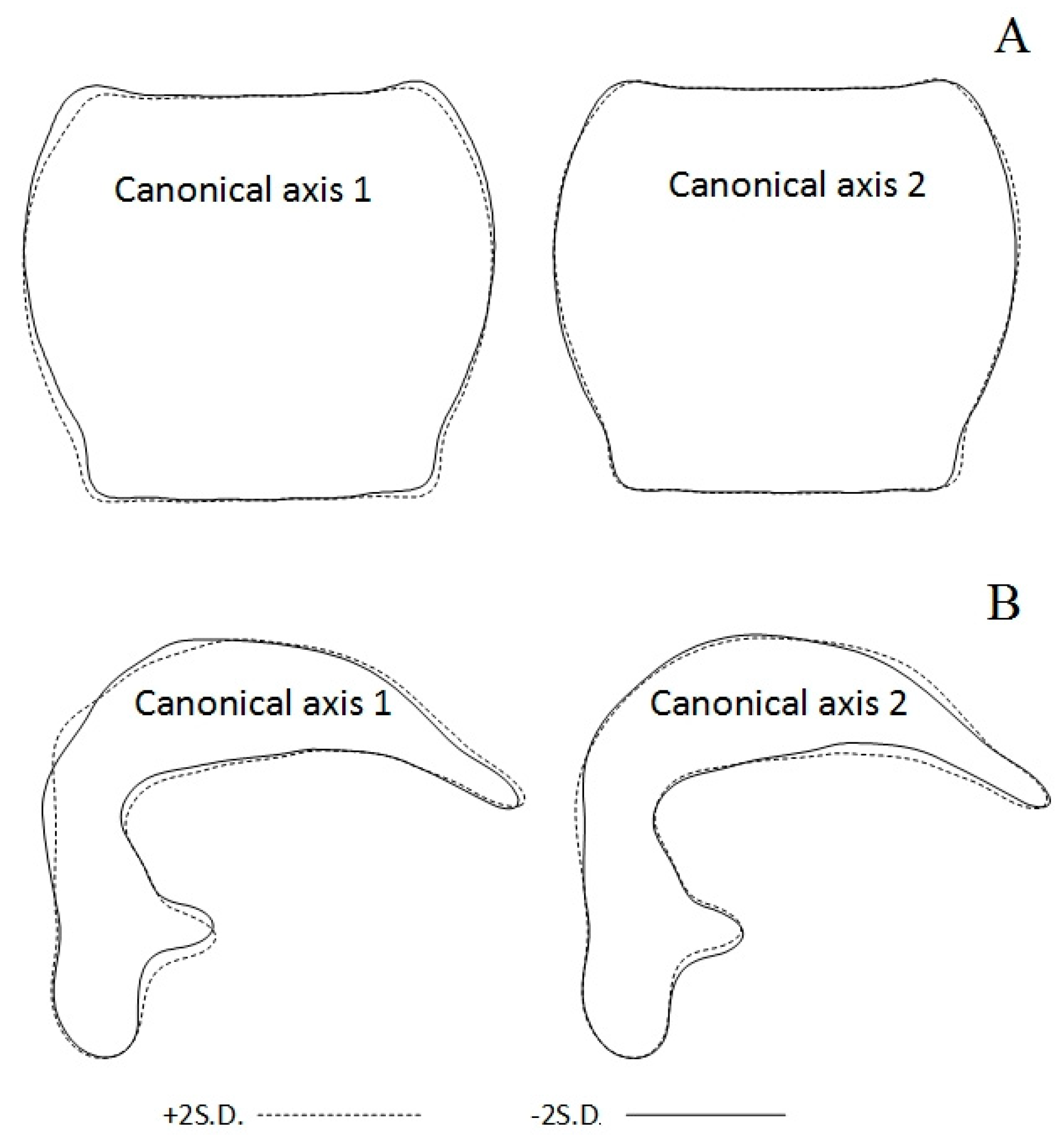
3.2. Shape Variation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chernov, Y.I.; Makarova, O.L.; Penev, L.D.; Khruleva, O.A. Beetles (Insecta, Coleoptera) in the Arctic Fauna: Communication 1. Faunal Composition. Entomol. Rev. 2014, 94, 438–478. [Google Scholar] [CrossRef]
- Chernov, Y.I.; Makarov, K.V.; Eremin, P.K. Family of Ground Beetles in the Arctic Fauna; Communication 1. Entomol. Rev. 2000, 80, 1009–1021. [Google Scholar]
- Chernov, Y.I.; Makarov, K.V.; Eremin, P.K. Family of Ground Beetles in the Arctic Fauna: Communication 2. Entomol. Rev. 2001, 81, 108–117. [Google Scholar]
- Löbl, I.; Löbl, D. (Eds.) Catalogue of Palaearctic Coleoptera. Volume 1. Revised and Updated Edition. Archostemata–Myxophaga–Adephaga; Brill: Leiden, The Netherlands; Boston, MA, USA, 2017; p. 1443. [Google Scholar] [CrossRef]
- Bousquet, Y. Supraspecific Classification of the Nearctic Pterostichini (Coleoptera: Carabidae); Association des Entomologists Amateurs du Québec: Varennes, QC, Canada, 1999; Volume 9, 292p. [Google Scholar]
- Bousquet, Y. Tribe Pterostichini Bonelli, 1810. In Catalogue of Palaearctic Coleoptera 1; Löbl, I., Smetana, A., Eds.; Apollo Books: Stenstrup, Denmark, 2003; pp. 469–520. [Google Scholar]
- Makarov, K.V.; Kryzhanovskij, O.L.; Belousov, I.A.; Kabak, I.I.; Kataev, B.M.; Shilenkov, V.G.; Matalin, A.V.; Fedorenko, D.N.; Komarov, E.V. A Taxonomic List of the Ground-Beetles (Carabidae) of Russia. 2020. Available online: http://www.zin.ru/Animalia/Coleoptera/rus/car_rus.htm (accessed on 10 August 2021).
- Danks, H.U. Arctic Arthropods; Publ. Entomol. Soc.: Ottawa, ON, Canada, 1981; 608p. [Google Scholar]
- Poppius, B. Zur Kenntnis der Pterostichen-Untergattung Cryobius Chaud. Acta Soc. pro Faun. Flora Fenn. 1906, 28, 1–280. [Google Scholar]
- Ball, G.E. A Revision of the North American Species of the subgenus Cryobius Chaudoir (Pterostichus, Carabidae, Coleoptera). Opusc. Entomol. 1966, 28, 165. [Google Scholar]
- Kryzhanovskij, O.L.; Belousov, I.A.; Kabak, I.I.; Kataev, B.M.; Makarov, K.V.; Shilinkov, V.G. A Check-List of the Ground-Beetles of Russia and Adjacent Lands (Insecta, Coleoptera, Carabidae); Pensoft Publishers: Sofia, Bulgaria; Moscow, Russia, 1995; 271p. [Google Scholar]
- Erjiomin, P.K. Palaearctic Species of the Pterostichus (Cryobius) brevicornis Group (Coleoptera, Carabidae). Zool. Zhurnal 1998, 77, 295–302. (In Russian) [Google Scholar]
- Will, K.W.; Gill, A.S. Phylogeny and classification of Hypherpes auctorum (Coleoptera: Carabidae: Pterostichini: Pterostichus). Ann. Carnegie Mus. 2008, 77, 93–127. [Google Scholar] [CrossRef]
- Gibson, C.M.; Kao, R.H.; Blevins, K.K.; Travers, P.D. Integrative Taxonomy for Continental-Scale Terrestrial Insect Observations. PLoS ONE 2012, 7, e37528. [Google Scholar] [CrossRef]
- Woodcock, T.S.; Boyle, E.E.; Roughley, R.E.; Kevan, P.G.; Labbee, R.N.; Smith, A.B.T.; Goulet, H.; Steinke, D.; Adamowicz, S.J. The diversity and biogeography of the Coleoptera of Churchill: Insights from DNA barcoding. BMC Ecol. 2013, 13, 40. [Google Scholar] [CrossRef] [Green Version]
- Hebert, P.D.N.; Ratnasingham, S.; Zakharov, E.V.; Telfer, A.C.; Levesque-Beaudin, V.; Milton, M.A.; Pedersen, S.; Jannetta, P.; Dewaard, J.R. Counting animal species with DNA barcodes: Canadian insects. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150333. [Google Scholar] [CrossRef] [Green Version]
- Sikes, D.S.; Bowser, M.; Morton, J.M.; Bickford, C.; Meierotto, S.; Hildebrandt, K. Building a DNA barcode library of Alaska’s non-marine arthropods. Genome 2017, 60, 248–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pentinsaari, M.; Blagoev, G.; Hogg, I.D.; Levesque-Beaudin, V.; Perez, K.; Sobel, C.N.; vanDenBrink, B.; Borisenko, A. A DNA Barcoding Survey of an Arctic Arthropod Community: Implications for Future Monitoring. Insects 2020, 11, 46. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, C.; Jordal, B.; Serrano, J. Molecular phylogeny of the tribe Sphodrini (Coleoptera: Carabidae) based on mitochondrial and nuclear markers. Mol. Phylogenet. Evol. 2009, 50, 44–58. [Google Scholar] [CrossRef] [PubMed]
- Sasakawa, K.; Kubota, K. Phylogeny and Genital Evolution of Carabid Beetles in the Genus Pterostichus and Its Allied Genera (Coleoptera: Carabidae) Inferred from Two Nuclear Gene Sequences. Ann. Entomol. Soc. Am. 2007, 100, 100–109. [Google Scholar] [CrossRef]
- Raupach, M.J.; Hannig, K.; Morinière, J.; Hendrich, L. A DNA barcode library for ground beetles of Germany: The genus Pterostichus Bonelli, 1810 and allied taxa (Insecta, Coleoptera, Carabidae). ZooKeys 2020, 980, 93–117. [Google Scholar] [CrossRef] [PubMed]
- Khobrakova, L.T.; Shilenkov, V.G.; Dudko, R.Y. The Ground Beetles (Coleoptera, Carabidae) of Buryatia; Buryat Scientific Center SB RAS Press: Ulan-Ude, Russia, 2014; 380p. (In Russian) [Google Scholar]
- Morita, S. Pterostichine carabid beetles of the subgenus Cryobius (Coleoptera, Carabidae) from north Japan. Elytra 2002, 30, 73–89. [Google Scholar]
- Baust, J.G. Temperature-induced neural adaptation: Motoneuron discharge in the Alaskan beetle Pterostichus brevicornis (Carabidae). Comp. Biochem. Physiol. 1972, A41, 205–213. [Google Scholar] [CrossRef]
- Baust, J.G.; Miller, L.K. Variations in glycerol content and its influence on cold hardiness in the Alaskan Carabid beetle Pterostichus brevicornis. J. Insect Physiol. 1970, 16, 979–990. [Google Scholar] [CrossRef]
- Kaufmann, T. Hibernation in the Arctic beetle, Pterostichus brevicornis, in Alaska. J. Kansas Entomol. Soc. 1971, 44, 81–92. [Google Scholar]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring, NY, USA, 1989; 1659p. [Google Scholar]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Hajibabaei, M.; Janzen, D.H.; Burns, J.M.; Hallwachs, W.; Hebert, P. DNA barcodes distinguish species of tropical Lepidoptera. Proc. Natl. Acad. Sci. USA 2006, 103, 968–971. [Google Scholar] [CrossRef] [Green Version]
- Park, J.-K.; Foighil, D.Ó. Sphaeriid and Corbiculid Clams Represent Separate Heterodont Bivalve Radiations into Freshwater Environments. Mol. Phylogenet. Evol. 2000, 14, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Jovelin, R.; Justine, J.-L. Phylogenetic relationships within the polyopisthocotylean monogeneans (Platyhelminthes) inferred from partial 28S rDNA sequences. Int. J. Parasitol. 2001, 31, 393–401. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT; Nucleic Acids Symposium Series; Information Retrieval Ltd.: London, UK, 1999; Volume 41, pp. 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Villesen, P. FaBox: An online toolbox for fasta sequences. Mol. Ecol. Notes 2007, 7, 965–968. [Google Scholar] [CrossRef]
- Talavera, G.; Castresana, J. Improvement of Phylogenies after Removing Divergent and Ambiguously Aligned Blocks from Protein Sequence Alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [Green Version]
- Posada, D. jModelTest: Phylogenetic Model Averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop, GCE 2010, New Orleans, LA, USA, 14 November 2010. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree v1.4.2. 2018. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 20 July 2021).
- Bandelt, H.J.; Forster, P.; Rohl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Garnier, S.; Magniez-Jannin, F.; Rasplus, J.-Y.; Alibert, P. When morphometry meets genetics: Inferring the phylogeography of Carabus solieri using Fourier analyses of pronotum and male genitalia. J. Evol. Biol. 2005, 18, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Kergoat, G.J.; Alvarez, N. Assessing the phylogenetic usefulness of a previously neglected morphological structure through elliptic Fourier analyses: A case study in Bruchus seed-beetles (Coleoptera: Chrysomelidae: Bruchinae). Syst. Entomol. 2008, 33, 289–300. [Google Scholar] [CrossRef] [Green Version]
- Polihronakis, M. Hierarchical comparative analysis of genetic and genitalic geographical structure: Testing patterns of male and female genital evolution in the scarab beetle Phyllophaga hirticula (Coleoptera: Scarabaeidae). Biol. J. Linn. Soc. 2008, 96, 135–149. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Wenzel, J.W. Mosaic pattern of genital divergence in three populations of Schistocerca lineata Scudder, 1899 (Orthoptera: Acrididae: Cyrtacanthacridinae). Biol. J. Linn. Soc. 2008, 94, 289–301. [Google Scholar] [CrossRef] [Green Version]
- Zúñiga-Reinoso, Á.; Benítez, H.A. The overrated use of the morphological cryptic species concept: An example with Nyctelia darkbeetles (Coleoptera: Tenebrionidae) using geometric morphometrics. Zool. Anz. J. Comp. Zool. 2015, 255, 47–53. [Google Scholar] [CrossRef]
- Tatsuta, H.; Takahashi, K.H.; Sakamaki, Y. Geometric morphometrics in entomology: Basics and applications. Entomol. Sci. 2018, 21, 164–184. [Google Scholar] [CrossRef] [Green Version]
- Iwata, H. SHAPE: A Computer Program Package for Quantitative Evaluation of Biological Shapes Based on Elliptic Fourier Descriptors. J. Hered. 2002, 93, 384–385. [Google Scholar] [CrossRef] [Green Version]
- Installation Guide, ZEN 2 (Blue edition). Available online: https://www.zeiss.com/content/dam/Microscopy/Downloads/Pdf/FAQs/zen2-blue-edition_installation-guide.pdf (accessed on 15 December 2020).
- Adobe Photoshop CC 2014. Available online: https://adobe-photoshop.ruprograms.com/adobe-photoshop-cc-2014 (accessed on 15 December 2020).
- StatSoft.ru. Available online: http://statsoft.ru/products/new-features/STATISTICA10.php (accessed on 15 December 2020).
- Hammer, Ø. PAST: Paleontological Statistics Version 3.06: Reference Manual; Natural History Museum University of Oslo: Oslo, Norway, 2015; 225p. [Google Scholar]
- Bolotov, I.N.; Aksenova, O.; Bespalaya, Y.; Gofarov, M.; Kondakov, A.; Paltser, I.S.; Stefansson, A.; Travina, O.; Vinarski, M. Origin of a divergent mtDNA lineage of a freshwater snail species, Radix balthica, in Iceland: Cryptic glacial refugia or a postglacial founder event? Hydrobiologia 2017, 787, 73–98. [Google Scholar] [CrossRef]
- Bolotov, I.N.; Tatarinov, A.G.; Filippov, B.Y.; Gofarov, M.Y.; Kondakov, A.V.; Kulakova, O.I.; Potapov, G.S.; Zubryi, N.A.; Spitsyn, V.M. The distribution and biology of Pararctia subnebulosa (Dyar, 1899) (Lepidoptera: Erebidae: Arctiinae), the largest tiger moth species in the High Arctic. Polar Biol. 2015, 38, 905–911. [Google Scholar] [CrossRef]
- Potapov, G.S.; Kondakov, A.V.; Filippov, B.; Gofarov, M.; Kolosova, Y.S.; Spitsyn, V.M.; Tomilova, A.A.; Zubrii, N.; Bolotov, I.N. Pollinators on the polar edge of the Ecumene: Taxonomy, phylogeography, and ecology of bumble bees from Novaya Zemlya. ZooKeys 2019, 866, 85–115. [Google Scholar] [CrossRef] [PubMed]
- Potapov, G.S.; Berezin, M.V.; Kolosova, Y.S.; Kondakov, A.V.; Tomilova, A.A.; Spitsyn, V.M.; Zheludkova, A.A.; Zubrii, N.A.; Filippov, B.Y.; Bolotov, I.N. The last refugia for a polar relict pollinator: Isolates of Bombus glacialis on Novaya Zemlya and Wrangel Island indicate its broader former range in the Pleistocene. Polar Biol. 2021, 44, 1691–1709. [Google Scholar] [CrossRef]
- Elias, S.A. Advances in Quaternary Entomology. In Developments in Quaternary Sciences; van der Meer, J.J.M., Ed.; Elsevier Publisher: Amsterdam, The Netherlands, 2010; Volume 12, pp. 1–288. [Google Scholar]
- Hughes, R.A.; Inouye, B.D.; Johnson, M.T.J.; Underwood, N.; Vellend, M. Ecological consequences of genetic diversity. Ecol. Lett. 2008, 11, 609–623. [Google Scholar] [CrossRef] [PubMed]
- Sota, T.; Nagata, N. Diversification in a fluctuating island setting: Rapid radiation of Ohomopterus ground beetles in the Japanese Islands. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 3377–3390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujisawa, T.; Sasabe, M.; Nagata, N.; Takami, Y.; Sota, T. Genetic basis of species-specific genitalia reveals role in species diversification. Sci. Adv. 2019, 5, eaav9939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hebert, P.D.N.; Ratnasingham, S.; De Waard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. B Boil. Sci. 2003, 270 (Suppl. S1), S96–S99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demari-Silva, B.; Multini, L.C.; Suesdek, L.; Oliveira, T.M.P.; Sallum, M.A.M.; Marrelli, M.T. Wing Morphometry and Genetic Variability between Culex coronator and Culex usquatus (Diptera: Culicidae), Two Sibling Species of the Coronator Group. J. Med. Entomol. 2017, 54, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Nice, C.C.; Shapiro, A.M. Molecular and morphological divergence in the butterfly genus Lycaeides (Lepidoptera: Lycaenidae) in North America: Evidence of recent speciation. J. Evol. Biol. 1999, 12, 936–950. [Google Scholar] [CrossRef] [Green Version]
- Raupach, M.J.; Astrin, J.J.; Hannig, K.; Peters, M.K.; Stoeckle, M.Y.; Wägele, J.-W. Molecular species identification of Central European ground beetles (Coleoptera: Carabidae) using nuclear rDNA expansion segments and DNA barcodes. Front. Zool. 2010, 7, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paill, W.; Koblmüller, S.; Friess, T.; Gereben-Krenn, B.-A.; Mairhuber, C.; Raupach, M.; Zangl, L. Relicts from Glacial Times: The Ground Beetle Pterostichus adstrictus Eschscholtz, 1823 (Coleoptera: Carabidae) in the Austrian Alps. Insects 2021, 12, 84. [Google Scholar] [CrossRef] [PubMed]
- Wachi, N.; Matsubayashi, K.W.; Maeto, K. Application of next-generation sequencing to the study of non-model insects. Entomol. Sci. 2018, 21, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Wang, G.; Li, M.; Peng, Z.; Ali, H.; Xu, L.; Gurr, G.M.; Hou, Y. Development of Single Nucleotide Polymorphism (SNP) Markers for Analysis of Population Structure and Invasion Pathway in the Coconut Leaf Beetle Brontispa longissima (Gestro) Using Restriction Site-Associated DNA (RAD) Genotyping in Southern China. Insects 2020, 11, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, S.R.; Cnaani, J.; Kinch, L.N.; Grishin, N.V.; Hines, H.M. A combined RAD-Seq and WGS approach reveals the genomic basis of yellow color variation in bumble bee Bombus terrestris. Sci. Rep. 2021, 11, 7996. [Google Scholar] [CrossRef] [PubMed]
- Suchan, T.; Espíndola, A.; Rutschmann, S.; Emerson, B.C.; Gori, K.; Dessimoz, C.; Arrigo, N.; Ronikier, M.; Alvarez, N. Assessing the potential of RAD-sequencing to resolve phylogenetic relationships within species radiations: The fly genus Chiastocheta (Diptera: Anthomyiidae) as a case study. Mol. Phylogenet. Evol. 2017, 114, 189–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svendsen, J.I.; Alexanderson, H.; Astakhov, V.I.; Demidov, I.; Dowdeswell, J.A.; Funder, S.; Gataullin, V.; Henriksen, M.; Hjort, C.; Houmark-Nielsen, M.; et al. Late Quaternary ice sheet history of northern Eurasia. Quat. Sci. Rev. 2004, 23, 1229–1271. [Google Scholar] [CrossRef]
- Vesović, N.; Ivanović, A.; Ćurčić, S. Sexual size and shape dimorphism in two ground beetle taxa, Carabus (Procrustes) coriaceus cerisyi and C. (Morphocarabus) kollari praecellens (Coleoptera: Carabidae)—A geometric morphometric approach. Arthropod Struct. Dev. 2019, 49, 1–9. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Populations | North Europe | West and Central Siberia | Northeast of Asia | Alaska | North Canada |
|---|---|---|---|---|---|
| North Europe | 0 | 0.59 | 0.83 | 0.55 | 0.49 |
| West and Central Siberia | 0.95 ± 0.31 | 0 | 0.27 | 0.35 | 0.19 |
| Northeast of Asia | 1.24 ± 0.41 | 0.57 ± 0.20 | 0 | 0.57 | 0.47 |
| Alaska | 1.01 ± 0.33 | 0.98 ± 0.30 | 1.09 ± 0.33 | 0 | 0.28 |
| North Canada | 0.76 ± 0.27 | 0.72 ± 0.23 | 0.73 ± 0.25 | 0.82 ± 0.26 | 0 |
| Parameters | North Europe | West and Central Siberia | Northeast of Asia | Alaska | North Canada |
|---|---|---|---|---|---|
| Sample size, N | 30 | 20 | 25 | 55 | 38 |
| No. of haplotypes | 5 | 4 | 5 | 9 | 8 |
| Haplotype diversity (h ± SD) | 0.674 ± 0.055 | 0.658 ± 0.070 | 0.423 ± 0.119 | 0.634 ± 0.055 | 0.767 ± 0.042 |
| Nucleotide diversity (π ± SD), % | 0.208 ± 0.15 | 0.640 ± 0.38 | 0.207 ± 0.15 | 0.638 ± 0.36 | 0.541 ± 0.32 |
| Body Units | Effect | Wilks’s Lambda | df1, df2 | F | p |
|---|---|---|---|---|---|
| Pronotum | Sex | 0.916 | 5, 59 | 1.248 | 0.223 |
| Population | 0.110 | 45, 267 | 2.272 | 0.0000 | |
| Sex × Population | 0.364 | 45, 267 | 1.502 | 0.056 | |
| Aedeagus | Population | 0.109 | 45, 155 | 2.207 | 0.0002 |
| Shape Variables | CA1 | CA2 | CA3 | CA4 | CA5 |
|---|---|---|---|---|---|
| Pronotum | |||||
| PC1 | 0.026 | −0.701 | 0.458 | 0.052 | 0.544 |
| PC2 | −0.337 | −0.063 | −0.058 | −0.933 | 0.091 |
| PC3 | 0.041 | −0.019 | −0.708 | 0.081 | 0.700 |
| PC4 | −0.657 | −0.334 | −0.244 | 0.557 | −0.295 |
| PC5 | −0.275 | 0.597 | 0.432 | 0.198 | 0.584 |
| Cumulative proportion | 0.584 | 0.748 | 0.882 | 0.958 | 1.000 |
| Aedeagus | |||||
| PC1 | 0.099 | −0.011 | −0.717 | −0.674 | −0.149 |
| PC2 | −0.087 | 0.794 | −0.350 | 0.432 | −0.231 |
| PC3 | −0.222 | 0.004 | 0.246 | −0.277 | −0.902 |
| PC4 | 0.763 | 0.321 | 0.452 | −0.214 | −0.254 |
| PC5 | 0.238 | −0.414 | −0.405 | 0.604 | −0.493 |
| Cumulative proportion | 0.550 | 0.786 | 0.889 | 0.972 | 1.000 |
| Pronotum | Aedeagus | |||
|---|---|---|---|---|
| r | p | r | p | |
| Simple Mantel Tests | ||||
| Shape × Gen | 0.089 | 0.287 | 0.199 | 0.288 |
| Shape × Geo | 0.218 | 0.147 | 0.600 | 0.033 |
| Partial Mantel Tests | ||||
| Shape × Gen (Geo) | −0.147 | 0.789 | 0.332 | 0.091 |
| Shape × Geo (Gen) | 0.253 | 0.124 | 0.579 | 0.032 |
| Geo × Gen (Shape) | 0.347 | 0.006 | 0.332 | 0.093 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zubrii, N.A.; Filippov, B.Y.; Kondakov, A.V.; Khruleva, O.A.; Rybalov, L.B.; Vikhreva, D.V. DNA Barcoding versus Morphological Variability of Pterostichus brevicornis brevicornis (Kirby, 1837) (Coleoptera, Carabidae) in the Arctic and Subarctic. Insects 2022, 13, 204. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020204
Zubrii NA, Filippov BY, Kondakov AV, Khruleva OA, Rybalov LB, Vikhreva DV. DNA Barcoding versus Morphological Variability of Pterostichus brevicornis brevicornis (Kirby, 1837) (Coleoptera, Carabidae) in the Arctic and Subarctic. Insects. 2022; 13(2):204. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020204
Chicago/Turabian StyleZubrii, Natalia Andreevna, Boris Yurevich Filippov, Alexander Vasilevich Kondakov, Olga Arturovna Khruleva, Leonid Borisovich Rybalov, and Darya Vitalievna Vikhreva. 2022. "DNA Barcoding versus Morphological Variability of Pterostichus brevicornis brevicornis (Kirby, 1837) (Coleoptera, Carabidae) in the Arctic and Subarctic" Insects 13, no. 2: 204. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020204