
Comparative Screening of Mexican, Rwandan and Commercial Entomopathogenic Nematodes to Be Used against Invasive Fall Armyworm, Spodoptera frugiperda

 , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Origin and Handling of the Fall Armyworm and Nematodes
2.2. Step 1: Screening of Mexican Entomopathogenic Nematodes against the Fall Armyworm
2.3. Step 2: Comparing the Virulence of Mexican, Rwandan and Commercial Entomopathogenic Nematodes against the Fall Armyworm
2.4. Step 3: Assessing the Dose-Dependent Effectiveness of the Most Promising Entomopathogenic Nematodes against Different Stages of the Fall Armyworm
2.5. Data Analyses
3. Results
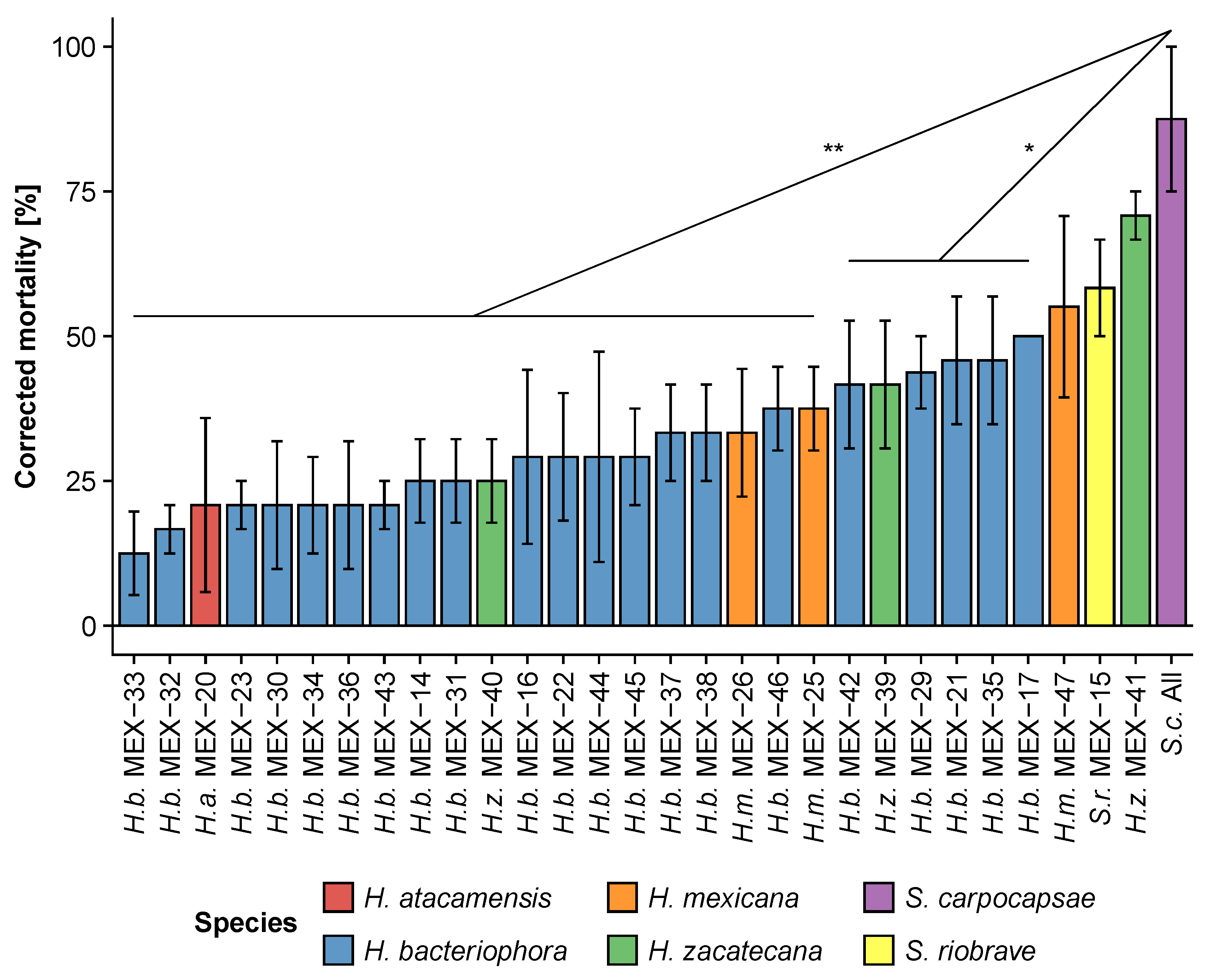
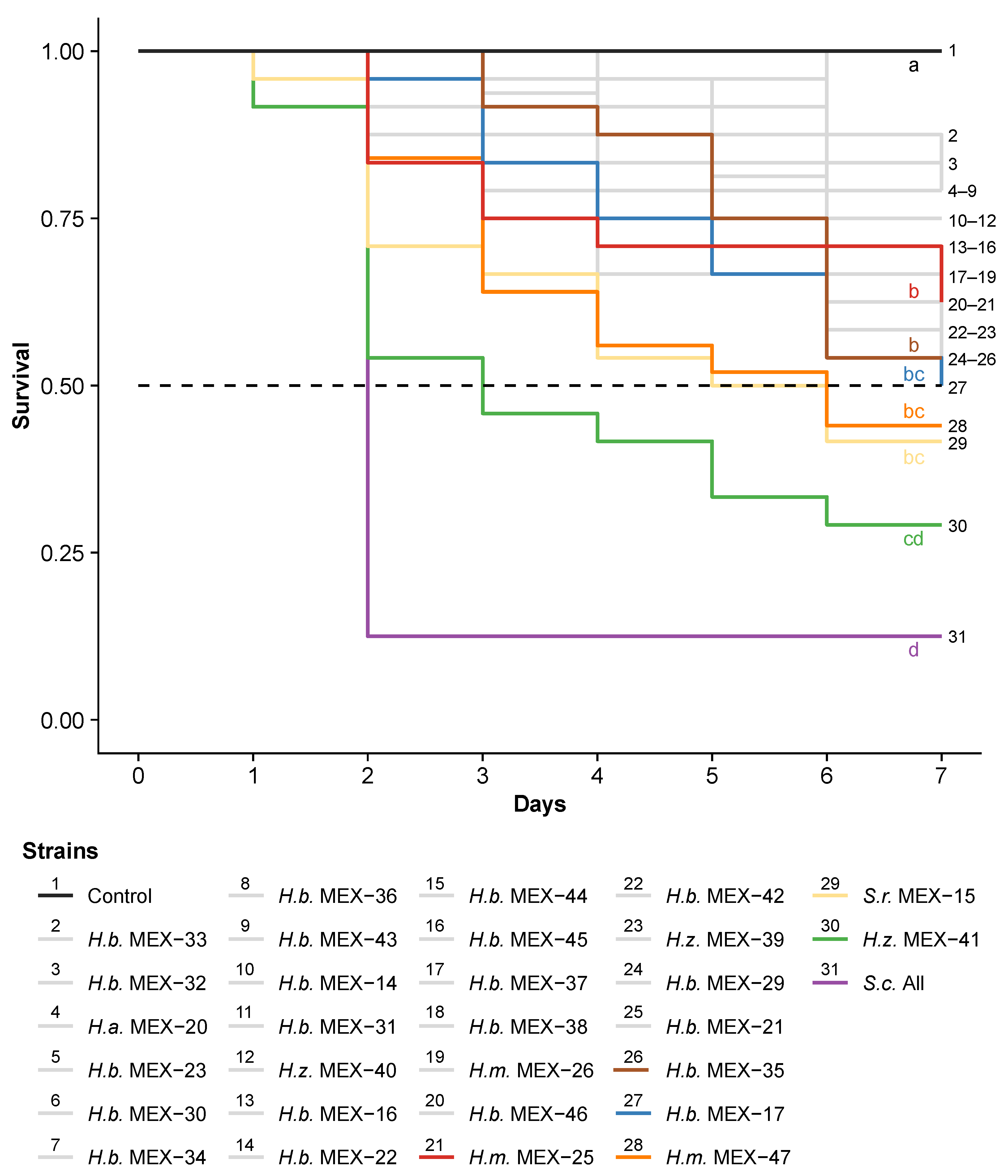
3.1. Step 1: Screening of Mexican Entomopathogenic Nematodes against the Fall Armyworm
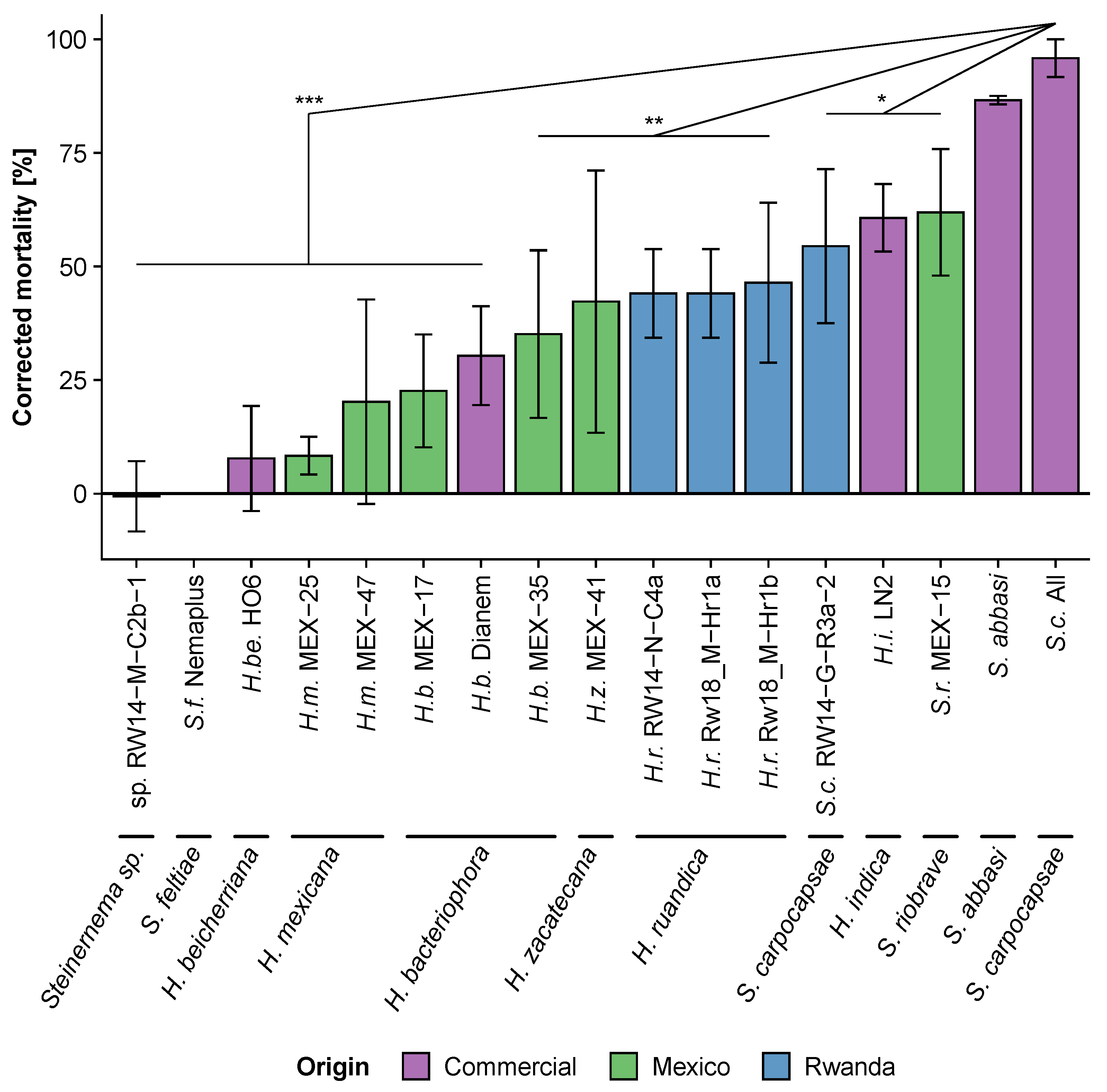
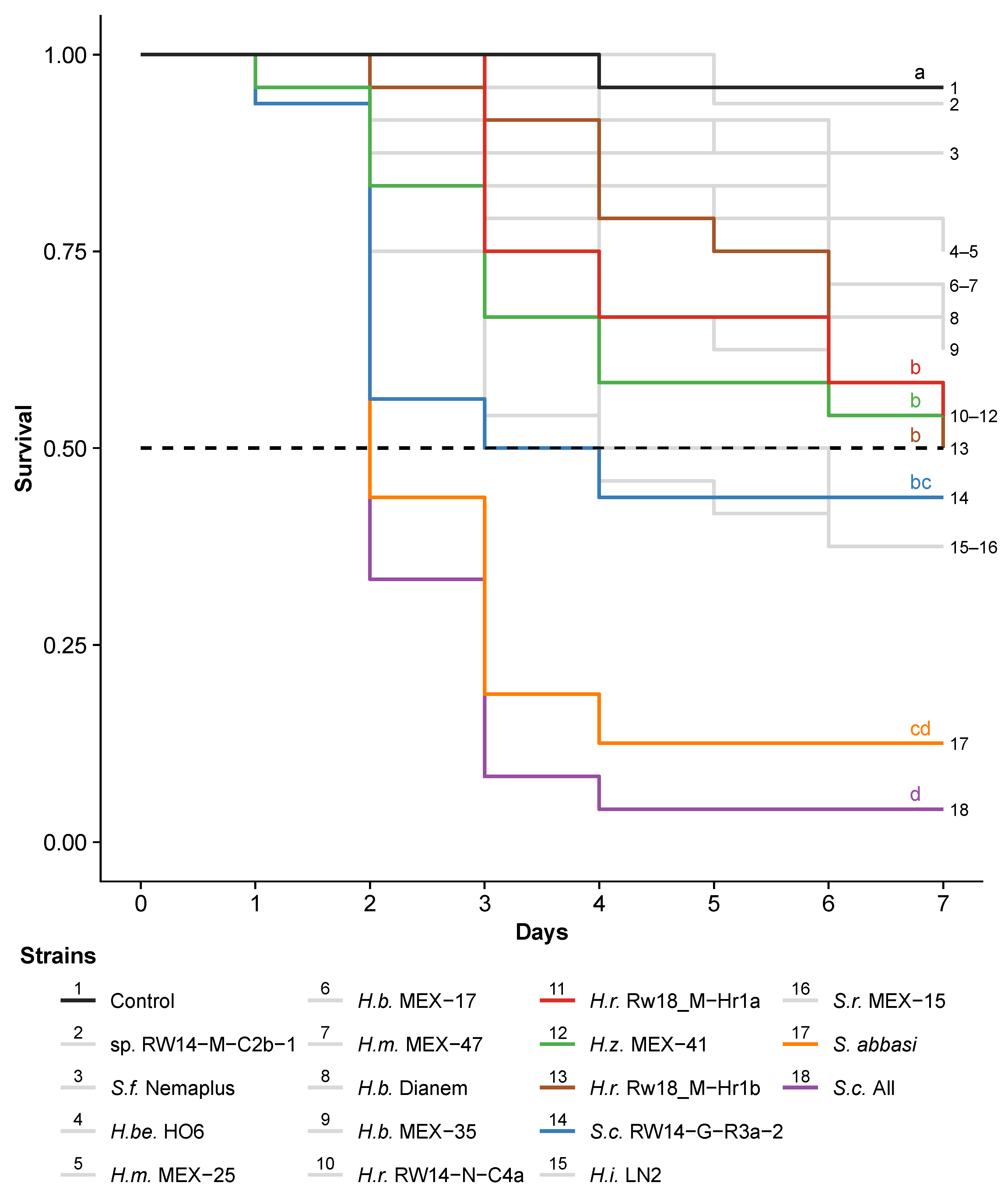
3.2. Step 2: Comparing the Virulence of Mexican, Rwandan and Commercial Entomopathogenic Nematodes against the Fall Armyworm
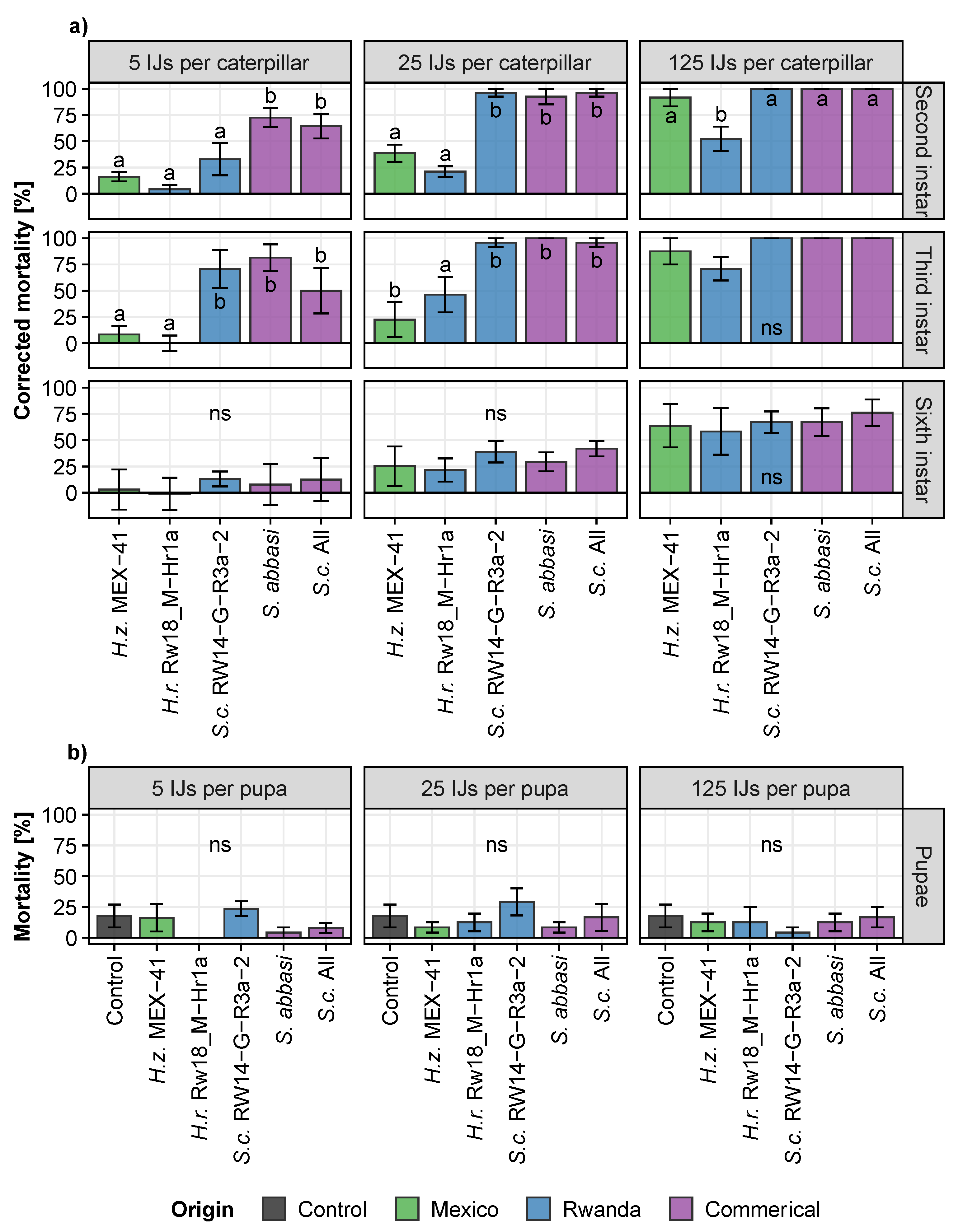
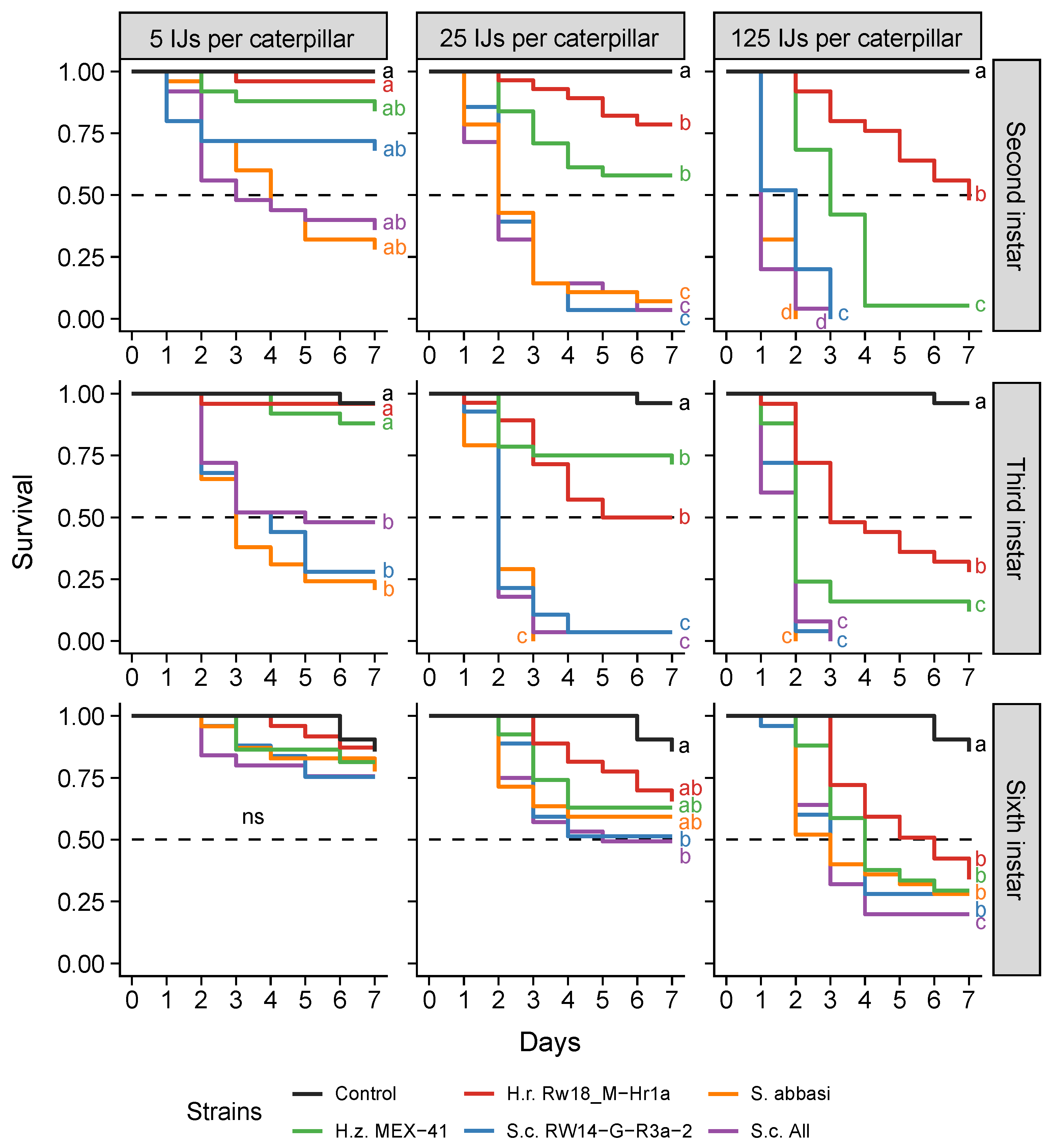
3.3. Step 3: Assessing the Dose-Dependent Effectiveness of the Most Promising Entomopathogenic Nematodes against Different Stages of the Fall Armyworm
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Luginbill, P. The Fall Army Worm; US Department of Agriculture Technical Bulletin No 34; US Department of Agriculture: Washington, DC, USA, 1928; p. 92.
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First report of outbreaks of the fall armyworm Spodoptera frugiperda (J E Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef] [Green Version]
- Sharanabasappa, D.; Kalleshwara Swamy, C.M.; Asokan, R.; Swamy, H.M.M.; Maruthi, M.S.; Pavithra, H.B.; Hegbe, K.; Navi, S.; Prabhu, S.T.; Goergen, G.E. First report of the Fall armyworm, Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae), an alien invasive pest on maize in India. Pest Manag. Hortic. Ecosyst. 2018, 24, 23–29. [Google Scholar]
- Cock, M.J.W.; Beseh, P.K.; Buddie, A.G.; Cafá, G.; Crozier, J. Molecular methods to detect Spodoptera frugiperda in Ghana, and implications for monitoring the spread of invasive species in developing countries. Sci. Rep. 2017, 7, 4103. [Google Scholar] [CrossRef]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J.; et al. Fall Armyworm: Impacts and Implications for Africa. Outlooks Pest Manag. 2017, 28, 196–201. [Google Scholar] [CrossRef] [Green Version]
- Baudron, F.; Zaman-Allah, M.A.; Chaipa, I.; Chari, N.; Chinwada, P. Understanding the factors influencing fall armyworm (Spodoptera frugiperda J.E. Smith) damage in African smallholder maize fields and quantifying its impact on yield. A case study in Eastern Zimbabwe. Crop Prot. 2019, 120, 141–150. [Google Scholar] [CrossRef]
- Rwomushana, I.; Bateman, M.; Beale, T.; Beseh, P.; Cameron, K.; Chiluba, M.; Clottey, V.; Davis, T.; Day, R.; Early, R.; et al. Fall Armyworm: Impacts and Implications for Africa. Evidence Note Update, October 2018. Available online: https://www.invasive-species.org/wp-content/uploads/sites/2/2019/02/FAW-Evidence-Note-October-2018.pdf (accessed on 24 November 2021).
- Hruska, A.J.; Gould, F. Fall Armyworm (Lepidoptera: Noctuidae) and Diatraea lineolata (Lepidoptera: Pyralidae): Impact of larval population level and temporal occurrence on maize yield in Nicaragua. J. Econ. Entomol. 1997, 90, 611–622. [Google Scholar] [CrossRef]
- Wan, J.; Huang, C.; Li, C.-Y.; Zhou, H.-X.; Ren, Y.-L.; Li, Z.-Y.; Xing, L.-S.; Zhang, B.; Qiao, X.; Liu, B.; et al. Biology, invasion and management of the agricultural invader: Fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Integr. Agric. 2021, 20, 646–663. [Google Scholar] [CrossRef]
- Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Day, R.; Early, R.; Godwin, J.; et al. Fall Armyworm: Impacts and Implications for Africa; CABI Publishing: Wallingford, UK, 2017. [Google Scholar]
- Tambo, J.A.; Day, R.K.; Lamontagne-Godwin, J.; Silvestri, S.; Beseh, P.K.; Oppong-Mensah, B.; Phiri, N.A.; Matimelo, M. Tackling fall armyworm (Spodoptera frugiperda) outbreak in Africa: An analysis of farmers’ control actions. Int. J. Pest Manag. 2020, 66, 298–310. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Wu, Q.-L.; Zhang, H.-W.; Wu, K.-M. Spread of invasive migratory pest Spodoptera frugiperda and management practices throughout China. J. Integr. Agric. 2021, 20, 637–645. [Google Scholar] [CrossRef]
- Georgis, R.; Koppenhöfer, A.M.; Lacey, L.A.; Bélair, G.; Duncan, L.W.; Grewal, P.S.; Samish, M.; Tan, L.; Torr, P.; van Tol, R.W.H.M. Successes and failures in the use of parasitic nematodes for pest control. Biol. Control 2006, 38, 103–123. [Google Scholar] [CrossRef]
- Hominick, W.H. Biogeography. In Entomopathogenic Nematology; Gaugler, R., Ed.; CABI Publishing: New York, NY, USA, 2002; pp. 115–143. [Google Scholar]
- Ehlers, R.U.; Hokkanen, H.M.T. Insect biocontrol with non-endemic entomopathogenic nematodes (Steinernema and Heterorhabditis spp.): Conclusions and recommendations of a combined OECD and COST workshop on scientific and regulatory policy issues. Biocontrol Sci. Technol. 1996, 6, 295–302. [Google Scholar] [CrossRef]
- Ehlers, R.-U. Mass production of entomopathogenic nematodes for plant protection. Appl. Microbiol. Biotechnol. 2001, 56, 623–633. [Google Scholar] [CrossRef]
- Andaló, V.; Santos, V.; Moreira, G.F.; Moreira, C.C.; Junior, A.M. Evaluation of entomopathogenic nematodes under laboratory and greenhouses conditions for the control of Spodoptera frugiperda Avaliação de nematoides entomopatogênicos em condições de laboratório e casa-de-vegetação visando ao. Cienc. Rural 2010, 40, 1860–1866. [Google Scholar] [CrossRef] [Green Version]
- Caccia, M.G.; Del Valle, E.; Doucet, M.E.; Lax, P. Susceptibility of Spodoptera frugiperda and Helicoverpa gelotopoeon (Lepidoptera: Noctuidae) to the entomopathogenic nematode Steinernema diaprepesi (Rhabditida: Steinernematidae) under laboratory conditions. Chil. J. Agric. Res. 2014, 74, 123–126. [Google Scholar] [CrossRef]
- Fuxa, J.R.; Richter, A.R.; Acudelo-Silva, F. Effect of host age and nematode strain on susceptibility of Spodoptera frugiperda to Steinernema feltiae. J. Nematol. 1988, 20, 91–95. [Google Scholar]
- Kaya, H.K.; Gaugler, R. Entomopathogenic Nematodes. Annu. Rev. Entomol. 1993, 38, 181–206. [Google Scholar] [CrossRef]
- Richter, A.R.; Fuxa, J.R. Effect of Steinernema feltiae on Spodoptera frugiperda and Heliothis zea (Lepidoptera: Noctuidae) in Corn. J. Econ. Entomol. 1990, 83, 1286–1291. [Google Scholar] [CrossRef]
- Acharya, R.; Hwang, H.-S.; Mostafiz, M.M.; Yu, Y.-S.; Lee, K.-Y. Susceptibility of various developmental stages of the fall armyworm, Spodoptera frugiperda, to entomopathogenic nematodes. Insects 2020, 11, 868. [Google Scholar] [CrossRef]
- Glazar, I.; Galper, S.; Sharon, E. Virulence of the nematode (Steinernematids and Heterorhabditis)-bacteria (Xenorhabdus spp.) complex to the Egyptian cotton leafworm Spodoptera littoralis (Lepidoptera: Noctuidae). J. Invertebr. Pathol. 1991, 57, 94–100. [Google Scholar] [CrossRef]
- Dhakal, M.; Nguyen, K.B.; Hunt, D.J.; Ehlers, R.-U.; Spiridonov, S.E.; Subbotin, S.A. Molecular identification, phylogeny and phylogeography of the entomopathogenic nematodes of the genus Heterorhabditis Poinar, 1976: A multigene approach. Nematology 2020, 23, 451–466. [Google Scholar] [CrossRef]
- Machado, R.A.R.; Bhat, A.H.; Abolafia, J.; Muller, A.; Bruno, P.; Fallet, P.; Arce, C.C.M.; Turlings, T.C.J.; Bernal, J.S.; Kajuga, J.; et al. Multi-locus phylogenetic analyses uncover species boundaries and reveal the occurrence of two new entomopathogenic nematode species, Heterorhabditis ruandica n. sp. and Heterorhabditis zacatecana n. sp. J. Nematol. 2021, 53, 1–42. [Google Scholar] [CrossRef]
- Hominick, W.M.; Reid, A.P.; Bohan, D.A.; Briscoe, B.R. Entomopathogenic nematodes: Biodiversity, geographical distribution and the convention on biological diversity. Biocontrol Sci. Technol. 1996, 6, 317–332. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Machado, R.A.R.; Doan, C.V.; Arce, C.C.M.; Hu, L.; Robert, C.A.M. Entomopathogenic nematodes increase predation success by inducing cadaver volatiles that attract healthy herbivores. eLife 2019, 8, e46668. [Google Scholar] [CrossRef]
- Bruno, P.; Machado, R.A.R.; Glauser, G.; Köhler, A.; Campos-Herrera, R.; Bernal, J.; Toepfer, S.; Erb, M.; Robert, C.A.M.; Arce, C.C.M.; et al. Entomopathogenic nematodes from Mexico that can overcome the resistance mechanisms of the western corn rootworm. Sci. Rep. 2020, 10, 8257. [Google Scholar] [CrossRef]
- Yan, X.; Waweru, B.; Qiu, X.; Hategekimana, A.; Kajuga, J.; Li, H.; Edgington, S.; Umulisa, C.; Han, R.; Toepfer, S. New entomopathogenic nematodes from semi-natural and small-holder farming habitats of Rwanda. Biocontrol Sci. Technol. 2016, 26, 820–834. [Google Scholar] [CrossRef] [Green Version]
- Fallet, P.; Machado, R.A.R.; Toepfer, S.; Ye, W.; Kajuga, J.; Waweru, B.; Bazagwira, D.; Turlings, T.C.J. A Rwandan survey of entomopathogenic nematodes that can potentially be used to control the fall armyworm. IOBC-WPRS Bull. 2020, 150, 87–90. [Google Scholar]
- Abbott, W.S. A Method of Computing the Effectiveness of an Insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- White, G.F. A method for obtaining infective nematode larvae form cultures. Science 1927, 66, 302–303. [Google Scholar] [CrossRef]
- Gelman, A.; Su, Y.-S. Arm: Data Analysis Using Regression and Multilevel/Hierarchical Models. R Package Version 1.11-2. 2020. Available online: https://CRAN.R-project.org/package=arm (accessed on 7 November 2021).
- Lenth, R.V. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. R Package Version 1.6.1. 2021. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 7 November 2021).
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Therneau, T. A Package for Survival Analysis in R. R Package Version 3.2-11. 2021. Available online: https://CRAN.R-project.org/package=survival (accessed on 7 November 2021).
- Kassambara, A.; Kosinski, M.; Biecek, P. Survminer: Drawing Survival Curves Using ‘ggplot2’. R Package Version 0.4.9. 2021. Available online: https://CRAN.R-project.org/package=survminer (accessed on 7 November 2021).
- Zeileis, A.; Kleiber, C.; Jackman, S. Regression models for count data in R. J. Stat. Softw. 2008, 27, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Riga, E.; Whistlecraft, J.; Potter, J. Potential of controlling insect pests of corn using entomopathogenic nematodes. Can. J. Plant Sci. 2001, 81, 783–787. [Google Scholar] [CrossRef]
- Garcia, L.C.; Raetano, C.G.; Leite, L.G. Application technology for the entomopathogenic nematodes Heterorhabditis indica and Steinernema sp. (Rhabditida: Heterorhabditidae and Steinernematidae) to control Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae) in corn. Neotrop. Entomol. 2008, 37, 305–311. [Google Scholar] [CrossRef] [Green Version]
- Salvadori, J.D.M.; Defferrari, M.S.; Ligabue-Braun, R.; Yamazaki Lau, E.; Salvadori, J.R.; Carlini, C.R. Characterization of entomopathogenic nematodes and symbiotic bacteria active against Spodoptera frugiperda (Lepidoptera: Noctuidae) and contribution of bacterial urease to the insecticidal effect. Biol. Control 2012, 63, 253–263. [Google Scholar] [CrossRef] [Green Version]
- Garcia-del-Pino, F.; Alabern, X.; Morton, A. Efficacy of soil treatments of entomopathogenic nematodes against the larvae, pupae and adults of Tuta absoluta and their interaction with the insecticides used against this insect. BioControl 2013, 58, 723–731. [Google Scholar] [CrossRef]
- Batalla-Carrera, L.; Morton, A.; García-del-Pino, F. Efficacy of entomopathogenic nematodes against the tomato leafminer Tuta absoluta in laboratory and greenhouse conditions. BioControl 2010, 55, 523–530. [Google Scholar] [CrossRef]
- James, M.; Malan, A.P.; Addison, P. Surveying and screening South African entomopathogenic nematodes for the control of the Mediterranean fruit fly, Ceratitis capitata (Wiedemann). Crop Prot. 2018, 105, 41–48. [Google Scholar] [CrossRef]
- Du Preez, F.; Malan, A.P.; Addison, P. Potential of in vivo- and in vitro-cultured entomopathogenic nematodes to infect Lobesia vanillana (Lepidoptera: Tortricidae) under laboratory conditions. PLoS ONE 2021, 16, e0242645. [Google Scholar] [CrossRef]
- Ramos-Rodríguez, O.; Campbell, J.F.; Ramaswamy, S.B. Pathogenicity of three species of entomopathogenic nematodes to some major stored-product insect pests. J. Stored Prod. Res. 2006, 42, 241–252. [Google Scholar] [CrossRef]
- Cagnolo, S.R.; Peschiutta, M.L.; Bertolotti, M.A. Susceptibility of adults of Anticarsia gemmatalis Hübner, 1818 (Lepidoptera: Noctuidae) to the entomopathogenic nematode Steinernema rarum (Doucet, 1986) Mamiya, 1988 (Rhabditida: Steinernematidae) under laboratory conditions. Nematology 2011, 13, 373–376. [Google Scholar] [CrossRef]
- Toepfer, S.; Hatala-Zseller, I.; Ehlers, R.-U.; Peters, A.; Kuhlmann, U. The effect of application techniques on field-scale efficacy: Can the use of entomopathogenic nematodes reduce damage by western corn rootworm larvae? Agric. For. Entomol. 2010, 12, 389–402. [Google Scholar] [CrossRef]
- Georgis, R. Formulation and application technology. In Entomopathogenic Nematodes in Biological Control; Gaugler, R., Kaya, H.K., Eds.; CRC Press: Boca Raton, FL, USA, 1990; pp. 173–191. [Google Scholar]
- Fallet, P.; Bazagwira, D.; Bustos, C.; Kajuga, J.; Mukundwa, I.P.; Karangwa, P.; Toepfer, S.; Turlings, T.C.J. Laboratory and field trials reveal the potential of a gel formulation of entomopathogenic nematodes as biocontrol against the fall armyworm (Spodoptera frugiperda). bioRxiv 2022. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.I.; Stuart, R.J.; McCoy, C.W. Targeted improvement of Steinernema carpocapsae for control of the pecan weevil, Curculio caryae (Horn) (Coleoptera: Curculionidae) through hybridization and bacterial transfer. Biol. Control 2005, 34, 215–221. [Google Scholar] [CrossRef]
- Hiltpold, I.; Baroni, M.; Toepfer, S.; Kuhlmann, U.; Turlings, T.C.J. Selection of entomopathogenic nematodes for enhanced responsiveness to a volatile root signal helps to control a major root pest. J. Exp. Biol. 2010, 213, 2417–2423. [Google Scholar] [CrossRef] [Green Version]
- Ehlers, R.-U.; Oestergaard, J.; Hollmer, S.; Wingen, M.; Strauch, O. Genetic selection for heat tolerance and low temperature activity of the entomopathogenic nematode–bacterium complex Heterorhabditis bacteriophora–Photorhabdus luminescens. BioControl 2005, 50, 699–716. [Google Scholar] [CrossRef]
- Nimkingrat, P.; Khanam, S.; Strauch, O.; Ehlers, R.-U. Hybridisation and selective breeding for improvement of low temperature activity of the entomopathogenic nematode Steinernema feltiae. BioControl 2013, 58, 417–426. [Google Scholar] [CrossRef]
- Bosa O, C.F.; Chávez, D.; Torres, L.; París, A.; Villamizar, L.; Cotes, A.M. Evaluación de aislamientos nativos de Nomuraea rileyi para el control de Spodoptera frugiperda (Lepidoptera: Noctuidae). Rev. Colomb. Entomol. 2004, 30, 93–97. [Google Scholar]
- García, C.G.; González, M.B.M.; Bautista, N.M. Pathogenicity of isolates of entomopathogenic fungi against Spodoptera frugiperda (Lepidoptera: Noctuidae) and Epilachna varivestis (Coleoptera: Coccinellidae). Patogenicidad de aislamientos de hongos entomopatógenos contra Spodoptera frugiperda (Lepidoptera: Noctuidae) y Epilachna varivestis (Coleoptera: Coccinellidae). Rev. Colomb. Entomol. 2011, 37, 217–222. [Google Scholar]
- Guo, J.; Wu, S.; Zhang, F.; Huang, C.; He, K.; Babendreier, D.; Wang, Z. Prospects for microbial control of the fall armyworm Spodoptera frugiperda: A review. BioControl 2020, 65, 647–662. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Origin | Strain | Species (Authority) | Source |
|---|---|---|---|
| Commercial | NA (Dianem®) | Heterorhabditis bacteriophora (Poinar) | Provided by e-nema GmbH, Schwentinental, Germany |
| NA | Steinernema abbasi (Elawad, Ahmad and Reid) | ||
| All (Nemastar®) | Steinernema carpocapsae (Weiser) | ||
| NA (Nemaplus®) | Steinernema feltiae (Filipjev) | ||
| H06 (HR-HB®) | Heterorhabditis beicherriana (Xing-Yue, Qi-Zhi, Nermut, Puza and Mracek) | Provided by Guangdong Academy of Sciences, China | |
| LN2 (HR-HI®) | Heterorhabditis indica (Poinar, Karunakar and David) | ||
| Mexico | MEX-20 | Heterorhabditis atacamensis (Edgington, Buddie, Moore, France, Merino and Hunt) | Collected by P. Bruno [28] |
| MEX-14 | Heterorhabditis bacteriophora | ||
| MEX-16 | |||
| MEX-17 | |||
| MEX-21 | |||
| MEX-22 | |||
| MEX-23 | |||
| MEX-29 | |||
| MEX-30 | |||
| MEX-31 | |||
| MEX-32 | |||
| MEX-33 | |||
| MEX-34 | |||
| MEX-35 | |||
| MEX-36 | |||
| MEX-37 | |||
| MEX-38 | |||
| MEX-42 | |||
| MEX-43 | |||
| MEX-44 | |||
| MEX-45 | |||
| MEX-46 | |||
| MEX-25 | Heterorhabditis mexicana (Nguyen, Sharpiro-Ilan, Stuart, McCoy, James and Adams) | ||
| MEX-26 | |||
| MEX-47 | |||
| MEX-39 | Heterorhabditis zacatecana (Machado, Bhat, Abolafia, Muller, Bruno, Fallet, Arce, Turlings, Bernal, Kajuga, Waweru and Toepfer) | ||
| MEX-40 | |||
| MEX-41 | |||
| MEX-15 | Steinernema riobrave (Cabanillas, Poinar and Raulston) | ||
| Rwanda | RW14-N-C4a | Heterorhabditis ruandica (Machado, Bhat, Abolafia, Muller, Bruno, Fallet, Arce, Turlings, Bernal, Kajuga, Waweru and Toepfer) | Provided by RAB, Rwanda [29] |
| RW14-G-R3a-2 | Steinernema carpocapsae | ||
| RW14-M-C2b-1 | Steinernema sp. (closely related to S. feltiae) | ||
| Rw18_M-Hr1a | Heterorhabditis ruandica | Provided by RAB, Rwanda [30] | |
| Rw18_M-Hr1b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fallet, P.; De Gianni, L.; Machado, R.A.R.; Bruno, P.; Bernal, J.S.; Karangwa, P.; Kajuga, J.; Waweru, B.; Bazagwira, D.; Degen, T.; et al. Comparative Screening of Mexican, Rwandan and Commercial Entomopathogenic Nematodes to Be Used against Invasive Fall Armyworm, Spodoptera frugiperda. Insects 2022, 13, 205. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020205
Fallet P, De Gianni L, Machado RAR, Bruno P, Bernal JS, Karangwa P, Kajuga J, Waweru B, Bazagwira D, Degen T, et al. Comparative Screening of Mexican, Rwandan and Commercial Entomopathogenic Nematodes to Be Used against Invasive Fall Armyworm, Spodoptera frugiperda. Insects. 2022; 13(2):205. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020205
Chicago/Turabian StyleFallet, Patrick, Lara De Gianni, Ricardo A. R. Machado, Pamela Bruno, Julio S. Bernal, Patrick Karangwa, Joelle Kajuga, Bancy Waweru, Didace Bazagwira, Thomas Degen, and et al. 2022. "Comparative Screening of Mexican, Rwandan and Commercial Entomopathogenic Nematodes to Be Used against Invasive Fall Armyworm, Spodoptera frugiperda" Insects 13, no. 2: 205. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020205