Potential Risk of Pollen from Genetically Modified MON 810 Maize Containing Cry1Ab Toxin to Protected Lepidopteran Larvae in the Pannonian Biogeographical Region—A Retrospective View


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
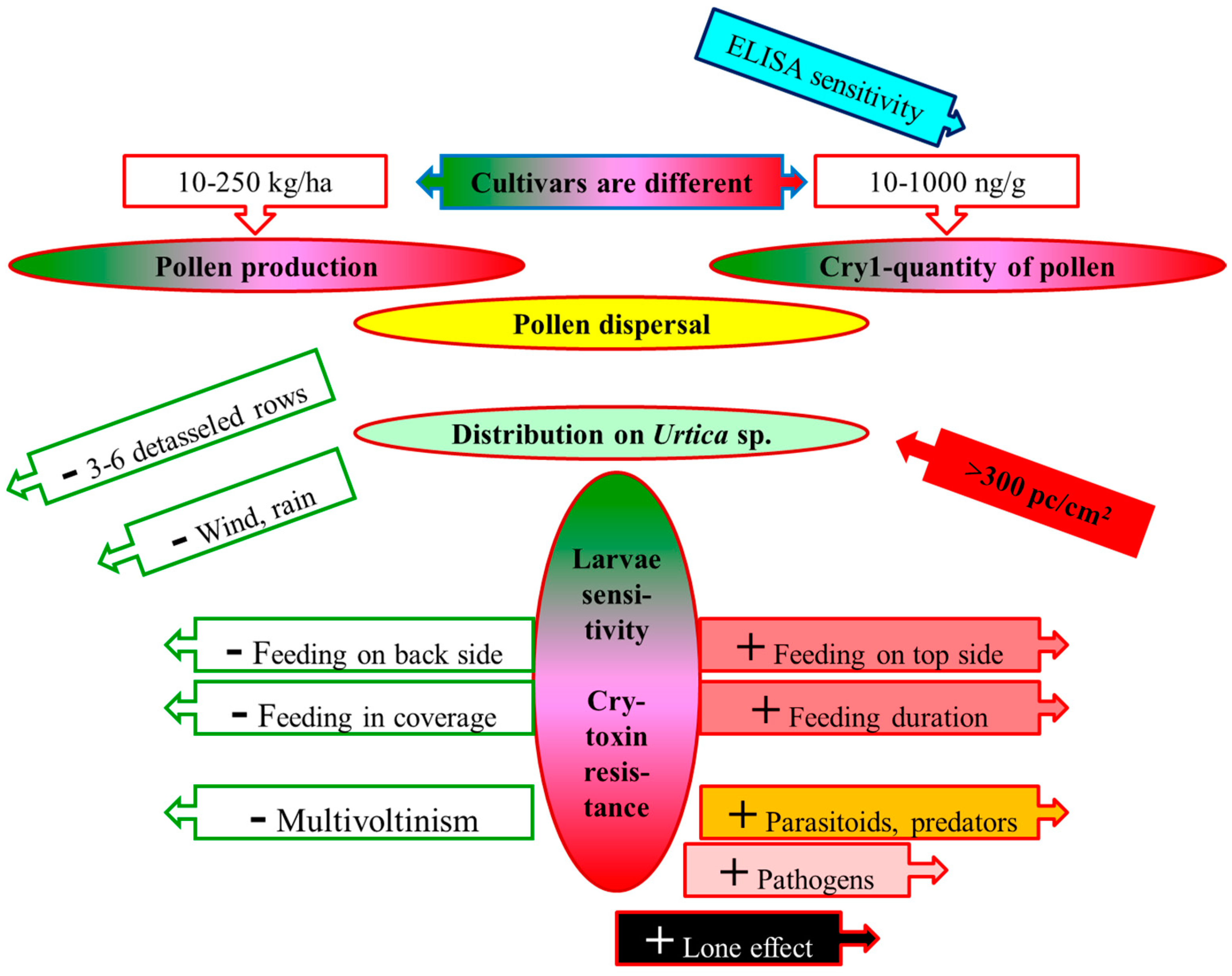
:Simple Summary
Abstract
1. Introduction
2. Pollen Production and Cry1Ab Toxin Content of MON 810 Cultivars
2.1. Differences between Genetically Modified Cultivars
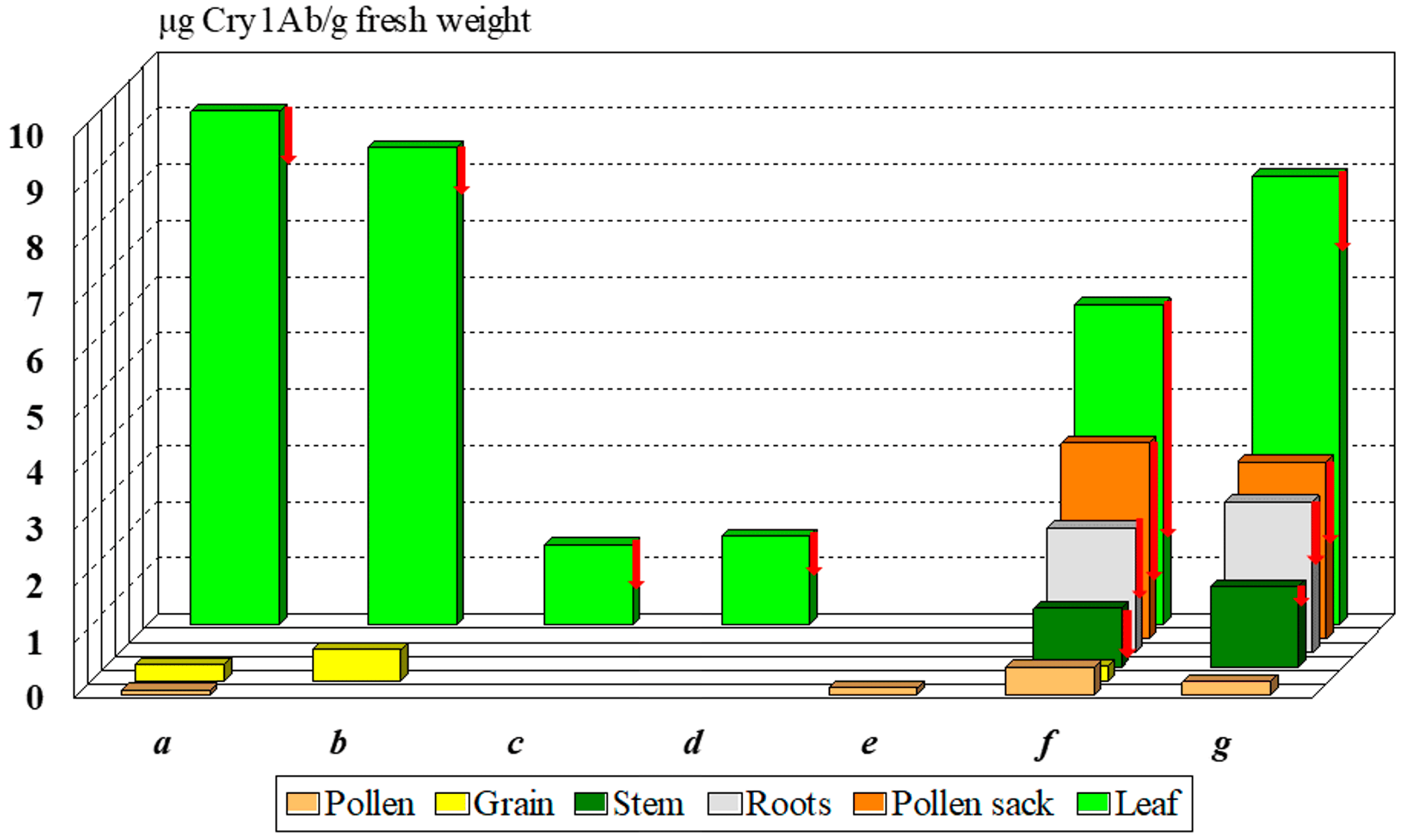
2.2. Cry1Ab Toxin Production Changes during Plant Development
2.3. Analytical Difficulties of Cry1Ab Determination
3. Frequent Weeds on Maize Field Edges in the Pannonian Biogeographical Region and Their Maize Pollen-Catching Capacity
4. Maize Pollen Distribution
4.1. Pollination Time of Maize in the Pannonian Biogeographical Region
4.2. Average Pollen Density of Maize Cultivars
5. Protected Lepidopteran Larvae near Edge of the Maize Field
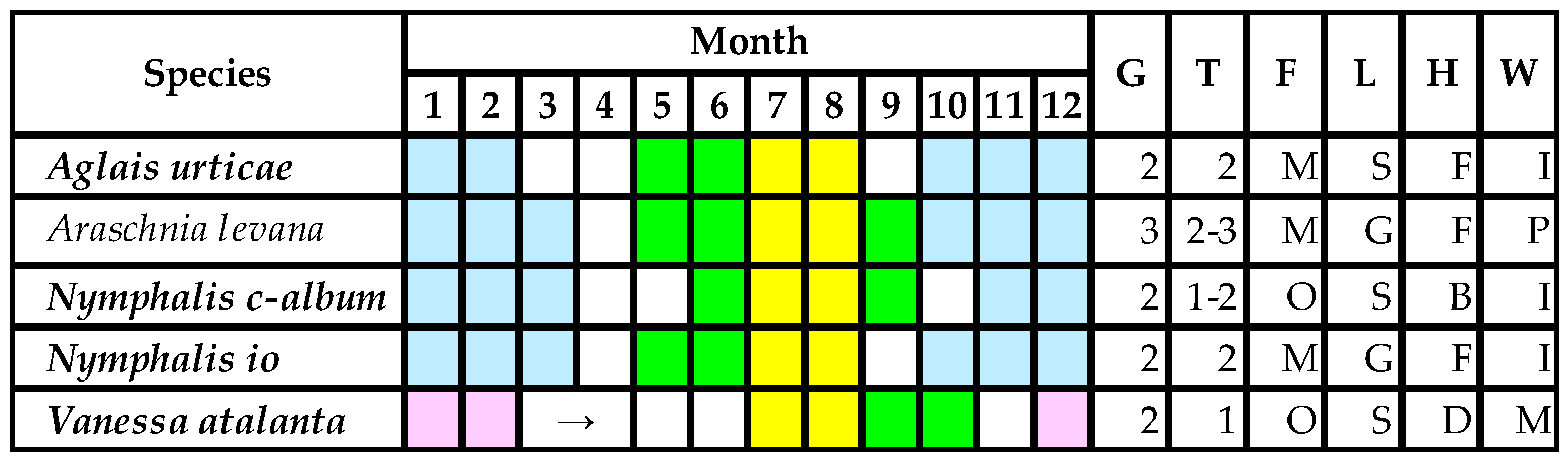
5.1. Lifestyles of Protected Lepidopteran Larvae Living on Urtica Species
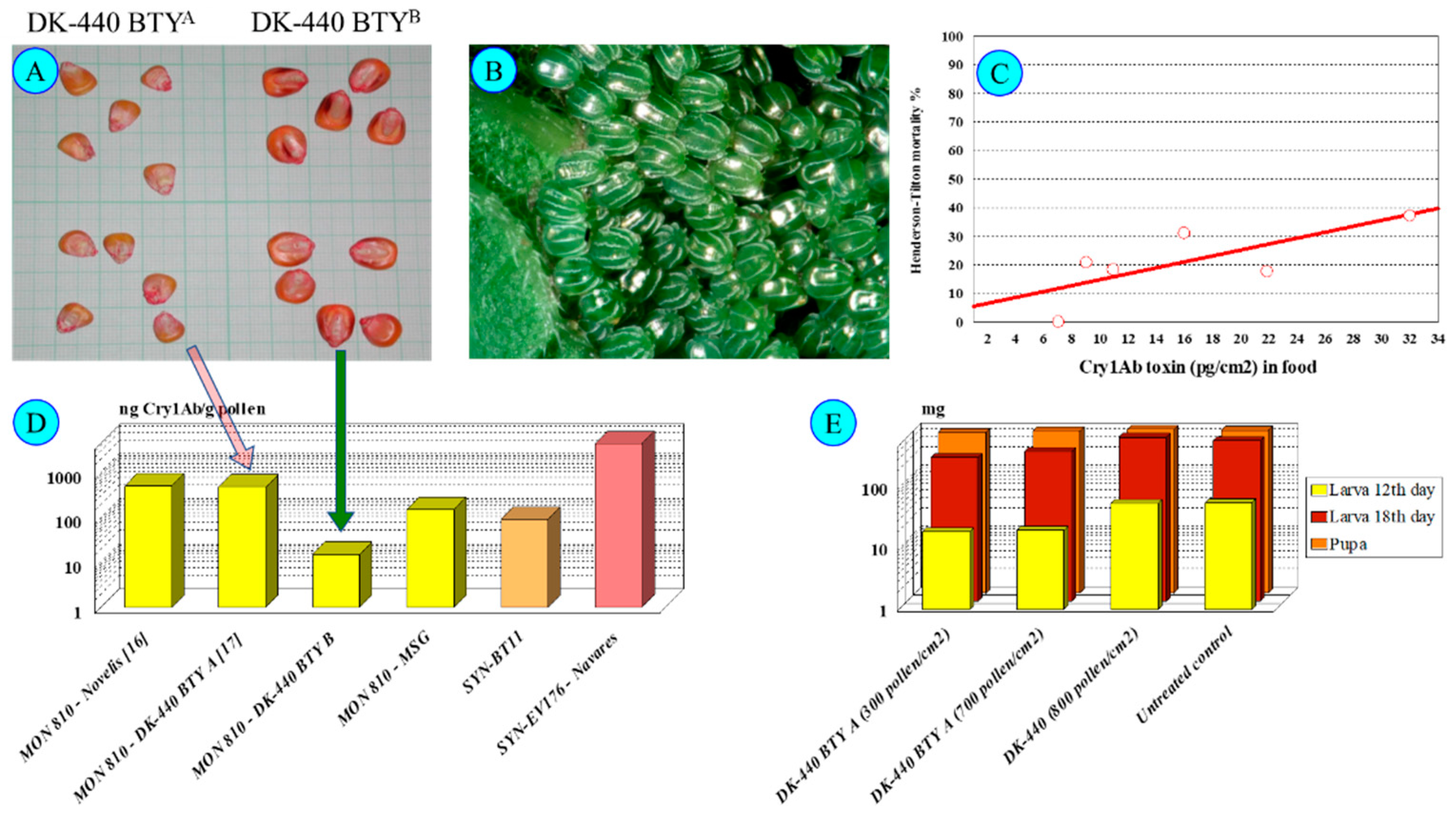
5.2. Toxicity Types of Cry1Ab on Nymphalis io Larvae
5.3. Variable Larval Sensitivities of Nymphalis io Subpopulations—Cry1 Toxin Resistance
6. Consequences for a Credible Hazard Assessment
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Losey, J.E.; Rayor, L.S.; Carter, M.A. Transgenic pollen harms monarch larvae. Nature 1999, 399, 214. [Google Scholar] [CrossRef] [PubMed]
- Felke, M.; Langenbruch, G.-A. Wirkung von Bt-mais-pollen auf raupen des tagpfauenauges in laborversuch. Gesunde Pflanz. 2003, 55, 1–7. (In German) [Google Scholar] [CrossRef]
- Pimentel, D.; Lach, L.; Zuniga, R.; Morrison, D. Environmental and economic costs of nonindigenous species in the United States. BioScience 2000, 50, 53–65. [Google Scholar] [CrossRef]
- Shelton, A.M.; Sears, M.K. The monarch butterfly controversy: Scientific interpretations of a phenomenon. Plant J. 2001, 27, 483–488. [Google Scholar] [CrossRef] [Green Version]
- Darvas, B.; Csóti, A.; Gharib, G.; Peregovits, L.; Ronkay, L.; Lauber, L.; Polgár, A.L. Some data to the risk analysis of Bt corn pollen and protected Lepidoptera species in Hungary. Növényvédelem 2004, 40, 441–449. (In Hungarian) [Google Scholar]
- Varga, Z. Conservation biogeographical aspects of the Pannonian Region. In Agrarian Biotechnology—First Generation GM Plants; Darvas, B., Ed.; Committee on Agriculture of the Hungarian Parliament: Budapest, Hungary, 2007; pp. 52–54. ISBN 978-963-87505-1-8. (In Hungarian) [Google Scholar]
- Hungarian Ministry of Environment. 13/2001. [V. 9.] Decree about Protected, Strictly Protected Plant and Animal Species, Strictly Protected Caves, and Plant and Animal Species of Community Interest; Hungarian Ministry of Environment: Budapest, Hungary, 2001. (In Hungarian) [Google Scholar]
- Hungarian Ministry of Environment and Water. 18/2008 [VI. 19.] Decree about Modification of 13/2001. [V. 9.] HME Decree about Protected, Strictly Protected Plant and Animal Species, Strictly Protected Caves, and Plant and Animal Species of Community Interest; Hungarian Ministry of Environment and Water: Budapest, Hungary, 2008. (In Hungarian) [Google Scholar]
- Darvas, B.; Székács, A. (Eds.) Hungarian Background on Views of 1st Generation Genetically Modified Plants; Agricultural Committee of the Hungarian Parliament: Budapest, Hungary, 2011; pp. 108–213. ISBN 978-963-9848-44-3. [Google Scholar]
- Székács, A.; Juracsek, J.; Polgár, L.A.; Darvas, B. Levels of expressed Cry1Ab toxin in genetically modified corn DK-440-BTY (YIELDGARD) and stubble. FEBS J. 2005, 272, 508. [Google Scholar]
- Székács, A.; Lauber, É.; Juracsek, J.; Darvas, B. Cry1Ab toxin production of MON 810 transgenic maize. Environ. Toxicol. Chem. 2010, 29, 182–190. [Google Scholar] [CrossRef]
- Székács, A.; Lauber, É.; Takács, E.; Darvas, B. Detection of Cry1Ab toxin in the leaves of MON 810 transgenic maize. Anal. Bioanal. Chem. 2010, 396, 2203–2211. [Google Scholar] [CrossRef]
- Székács, A. Cry1Ab toxin quantification in MON 810 maize. In GM-Crop Cultivation—Ecological Effects on a Landscape Scale. Theorie in der Ökologie; Breckling, B., Verhoeven, R., Eds.; Peter Lang: Frankfurt, Germany, 2013; Volume 17, pp. 123–124. ISBN 978-363-1628-70-6. [Google Scholar]
- Darvas, B.; Bánáti, H.; Takács, E.; Lauber, É.; Szécsi, A.; Székács, A. Relationships of Helicoverpa armigera, Ostrinia nubilalis and Fusarium verticillioides on MON 810 maize. Insects 2011, 2, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Bánáti, H.; Darvas, B.; Fehér-Tóth, S.; Czéh, Á.; Székács, A. Determination of mycotoxin production of Fusarium species in genetically modified maize varieties by quantitative flow immunocytometry. Toxins 2017, 9, 70. [Google Scholar] [CrossRef] [Green Version]
- Székács, A. Environmental analytical and ecotoxicological aspects of Bt maize in the Pannonian Biogeographical Region of the European Union. In GMOs: Implications for Biodiversity Conservation and Ecological Processes. Topics in Biodiversity and Conservation; Hawksworth, D.L., de Miranda, M.P., Chaurasia, A., Eds.; Springer Nature: Cham, Germany, 2020; Volume 19, pp. 149–172. ISBN 978-3-030-53182-9. [Google Scholar]
- Lauber, É. Cry1 Toxin Content and Pollen Effects of MON 810 Bt Maize on Hungarian Protected Butterflies. Ph.D. Thesis, Budapesti Corvinus Egyetem Kertészettudományi Kar, Budapest, Hungary, 2011; pp. 1–107. (In Hungarian). [Google Scholar]
- Székács, A.; Darvas, B. Environmental assessment of MON 810 maize in the Pannonian Biogeographical Region. Acta Phytopathol. Entomol. Hung. 2012, 47, 307–320. [Google Scholar] [CrossRef]
- Perry, N.J.; Devos, Y.; Arpaia, S.; Bartsch, D.; Gathmann, A.; Hails, R.S.; Kiss, J.; Lheureux, K.; Manachini, B.; Mestdagh, S.; et al. A mathematical model of exposure of non-target Lepidoptera to Bt-maize pollen expressing Cry1Ab within Europe. Proc. R. Soc. B 2010, 277, 1417–1425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holst, N.; Lang, A.; Lövei, G.L.; Otto, M. Increased mortality is predicted of Inachis io larvae caused by Bt-maize pollen in European farmland. Ecol. Model. 2013, 250, 126–133. [Google Scholar] [CrossRef]
- Fahsea, L.; Papastefanoua, P.; Otto, M. Estimating acute mortality of Lepidoptera caused by the cultivation of insect-resistant Bt maize—The LepiX model. Ecol. Model. 2018, 371, 50–59. [Google Scholar] [CrossRef]
- Baudrot, V.; Walker, E.; Lang, A.; Stefanescu, C.; Rey, J.-F.; Soubeyrand, S.; Messéan, A. When the average hides the risk of Bt-corn pollen on non-target Lepidoptera: Application to Aglais io in Catalonia. Ecotoxicol. Environ. Saf. 2021, 207, 111215. [Google Scholar] [CrossRef] [PubMed]
- Brookes, G.; Barfoot, P.; Melé, E.; Messeguer, J.; Bénétrix, F.; Bloc, D.; Foueillassar, X.; Fabié, A.; Poeydomenge, C. Genetically Modified Maize: Pollen Movement and Crop Coexistence; PG Economics Ltd.: Dorchester, UK, 2004. [Google Scholar]
- Sanders, P.R.; Elswick, E.N.; Groth, M.E.; Ledesma, B.E. Evaluation of Insect Protected Maize Lines in 1994 U.S. Field Test Location; Study 94-01-39-01, MSL-14179; Monsanto Company: St. Louis, MI, USA, 1995; EPA MRID No 43665502. [Google Scholar]
- AGBIOS. Safety Assesment of YieldGard Insect-Protected Event MON 810; Product Safety Description; AGBIOS: 2002. Available online: https://www.agbios.com (accessed on 13 February 2021).
- Bruns, H.A.; Abel, C.A. Nitrogen fertility effects on Bt δ-endotoxin and nitrogen concentrations of maize during early growth. Agron. J. 2003, 95, 207–211. [Google Scholar]
- Abel, C.A.; Adamczyk, J.J. Relative concentration of Cry1A in maize leaves and cotton bolls with diverse chlorophyll content and corresponding larval development of Fall Armyworm (Lepidoptera: Noctuidae) and Southwestern Corn Borer (Lepidoptera: Crambidae) on maize whorl leaf profiles. J. Econ. Entomol. 2004, 97, 1737–1744. [Google Scholar]
- Nguyen, H.T.; Jehle, J.J. Quantitative analysis of the seasonal and tissue-specific expression of Cry1Ab in transgenic maize Mon 810. J. Plant Dis. Prot. 2007, 114, 82–87. [Google Scholar] [CrossRef]
- Lang, A.; Ludy, C.; Vojtech, E. Dispersion and deposition of Bt maize pollen in field margins. J. Plant Dis. Prot. 2004, 111, 417–428. [Google Scholar]
- Takács, E.; Darvas, B.; Székács, A. Analytical difficulties and certain biological aspects of Cry1Ab toxin determination in MON 810 genetically modified maize. Acta Phytopathol. Entomol. Hung. 2012, 47, 293–306. [Google Scholar] [CrossRef]
- Székács, A.; Weiss, G.; Quist, D.; Takács, E.; Darvas, B.; Meier, M.; Swain, T.; Hilbeck, A. Inter-laboratory comparison of Cry1Ab toxin quantification in MON 810 maize by enzyme-immunoassay. Food Agric. Immunol. 2012, 23, 99–121. [Google Scholar] [CrossRef]
- Csóti, A. Data for the Risk Analysis for Protected Larvae and Bt-Corn Pollen in Hungary. Master’s Thesis, Budapesti Közgazdaságtudományi és Államigazgatási Egyetem, Kertészettudományi Kar, Budapest, Hungary, 2004; pp. 1–39. (In Hungarian). [Google Scholar]
- Pleasants, J.M.; Hellmich, R.L.; Dively, G.P.; Sears, M.K.; Stanley-Horn, D.E.; Matilla, H.R.; Foster, J.E.; Clark, P.; Jones, G.D. Corn pollen deposition on milkweeds in and near cornfield. Proc. Natl. Acad. Sci. USA 2001, 98, 11919–11924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarosz, N.; Loubet, B.; Huber, L. Modelling airborne concentration and deposition rate of maize pollen. Atmos. Environ. 2004, 38, 5555–5566. [Google Scholar] [CrossRef] [Green Version]
- Aylor, D.E. Quantifying maize pollen movement in a maize canopy. Agric. For. Meteorol. 2005, 131, 247–256. [Google Scholar] [CrossRef]
- Hofmann, F.; Otto, M.; Wosniok, W. Maize pollen deposition in relation to distance from the nearest pollen source under common cultivation—results of 10 years of monitoring (2001 to 2010). Environ. Sci. Eur. 2014, 26, 24. [Google Scholar] [CrossRef] [Green Version]
- Lang, A.; Lauber, É.; Darvas, B. Early-tier tests insufficient for GMO risk assessment. Nat. Biotechnol. 2007, 25, 35–36. [Google Scholar] [CrossRef]
- Székács, A.; Darvas, B. Comparative aspects of Cry toxin usage in insect control. In Advanced Technologies for Managing Insect Pests; Ishaaya, I., Palli, S.R., Horowitz, A.R., Eds.; Springer Science+Business Media: Dordrecht, The Netherlands, 2012; pp. 195–230. ISBN 978-94-007-4497-4. [Google Scholar]
- Müller, A.-K.; Schuppener, M.; Rauschen, S. Assessing the impact of Cry1Ab expressing corn pollen on larvae of Aglais urticae in a laboratory bioassay. Abs. IOBC Conf. 2011, 73, 55–60. [Google Scholar]
- Schuppener, M.; Mühlhause, J.; Müller, A.-K.; Rauschen, S. Environmental risk assessment for the small tortoiseshell Aglais urticae and a stacked Bt-maize with combined resistances against Lepidoptera and Chrysomelidae in central European agrarian landscapes. Molec. Ecol. 2012, 21, 4646–4662. [Google Scholar] [CrossRef]
- Lauber, É.; Darvas, B. Increased mortality of isolated first instar larvae of Inachis io (Lepidoptera). Acta Phytopathnol. Entomol. Hung. 2009, 44, 111–117. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Finson, N.; Groeters, F.R.; Moar, W.J.; Johnson, M.W.; Luo, K.; Adang, M.J. Reversal of resistance to Bacillus thuringiensis in Plutella xylostella. Proc. Natl. Acad. Sci. USA 1994, 91, 4120–4124. [Google Scholar] [CrossRef] [Green Version]
- Alves, A.P.; Spencer, T.A.; Tabashnik, B.E.; Siegfried, B.D. Inheritance of resistance to the Cry1Ab Bacillus thuringiensis toxin in Ostrinia nubilalis (Lepidoptera: Crambidae). J. Econ. Entomol. 2006, 99, 494–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabashnik, B.E.; Huang, F.; Ghimire, M.N.; Leonard, B.R.; Siegfried, B.D.; Rangasamy, M.; Yang, Y.; Wu, Y.; Gahan, L.J.; Heckel, D.G.; et al. Efficacy of genetically modified Bt toxins against insects with different genetic mechanisms of resistance. Nat. Biotechnol. 2011, 29, 1128–1131. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Brévault, T.; Carrière, Y. Insect resistance to Bt crops: Lessons from the first billion acres. Nat. Biotechnol. 2013, 31, 510–521. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Carrière, Y. Surge in insect resistance to transgenic crops and prospects for sustainability. Nat. Biotechnol. 2017, 35, 926–935. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Boyd, S.D.; Bulla, L.A.; Winkle, D.D. The defined toxin-binding region of the cadherin G-protein coupled receptor, BT-R1, for the active Cry1Ab toxin of Bacillus thuringiens. J. Proteom. Bioinform. 2018, 11, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Lang, A.; Lee, M.; Dolek, M.; Berchtold, J.; Otto, M. Laboratory tests with Lepidoptera to assess non-target effects of Bt maize pollen: Analysis of current studies and recommendations for a standardised design. Environ. Sci. Eur. 2019, 31, 39. [Google Scholar] [CrossRef]
- Lövei, G.L.; Lang, A.; Ferrante, M.; Bacle, V. Can the growing of transgenic maize threaten protected Lepidoptera in Europe? Insect Sci. 2021, 28, 1159–1168. [Google Scholar] [CrossRef]
- Perry, J.N.; Barberi, P.; Bartsch, D.; Birch, A.N.E.; Gathmann, A.; Kiss, J.; Manachini, B.; Nuti, M.; Rauschen, S.; Schiemann, J.; et al. Response to Kruse-Plass et al. regarding the risk to non-target lepidopteran larvae exposed to pollen from one or more of three Bt maize events (MON810, Bt11 and 1507). Environ. Sci. Eur. 2017, 29, 21. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority. Scientific opinion updating risk management recommendations to limit exposure of non-target Lepidoptera of conservation concern in protected habitats to Bt-maize pollen. EFSA J. 2015, 13, 4127. [Google Scholar]
- Hofmann, F.; Kruse-Plass, M.; Kuhn, U.; Otto, M.; Schlechtriemen, U.; Schröder, B.; Vögel, R.; Wosniok, W. Accumulation and variability of maize pollen deposition on leaves of European Lepidoptera host plants and relation to release rates and deposition determined by standardised technical sampling. Environ. Sci. Eur. 2016, 28, 14. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority. Relevance of a new scientific publication (Hofmann et al. 2016) for previous environmental risk assessment conclusions and risk management recommendations on the cultivation of Bt-maize events MON810, Bt11 and 1507. EFSA Suppl. Public 2016, 13, 1070E. [Google Scholar]
- Kruse-Plass, M.; Hofmann, F.; Kuhn, U.; Otto, M.; Schlechtriemen, U.; Schröder, B.; Vögel, R.; Wosniok, W. Reply to the EFSA (2016) on the relevance of recent publications (Hofmann et al. 2014, 2016) on environmental risk assessment and management of Bt-maize events (MON810, Bt11 and 1507). Environ. Sci. Eur. 2017, 29, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darvas, B.; Polgár, A.L. Novel-type insecticides: Specificity and effects on non-target organisms. In Insecticides with Novel Modes of Action: Mechanisms and Application; Ishaaya, I., Degheele, D., Eds.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1998; pp. 188–259. ISBN 3-540-63058-9. [Google Scholar]
- Melé, E.; Nadal, A.; Messeguer, J.; Melé-Messeguer, M.; Palaudelmàs, M.; Peñas, G.; Piferrer, X.; Capellades, G.; Serra, J.; Pla, M. Modeling gene flow distribution within conventional fields and development of a simplified sampling method to quantify adventitious GM contents in maize. Sci. Rep. 2015, 5, 17106. [Google Scholar] [CrossRef]
- Darvas, B.; Bánáti, H.; Takács, E.; Vajda, B.; Neszlényi, K.; Székács, A. Maize intraspecific hybridization—GM x non-GM variety. bioKontroll 2012, 3, 35–36. (In Hungarian) [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Darvas, B.; Gyurcsó, G.; Takács, E.; Székács, A. Potential Risk of Pollen from Genetically Modified MON 810 Maize Containing Cry1Ab Toxin to Protected Lepidopteran Larvae in the Pannonian Biogeographical Region—A Retrospective View. Insects 2022, 13, 206. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020206
Darvas B, Gyurcsó G, Takács E, Székács A. Potential Risk of Pollen from Genetically Modified MON 810 Maize Containing Cry1Ab Toxin to Protected Lepidopteran Larvae in the Pannonian Biogeographical Region—A Retrospective View. Insects. 2022; 13(2):206. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020206
Chicago/Turabian StyleDarvas, Béla, Gergő Gyurcsó, Eszter Takács, and András Székács. 2022. "Potential Risk of Pollen from Genetically Modified MON 810 Maize Containing Cry1Ab Toxin to Protected Lepidopteran Larvae in the Pannonian Biogeographical Region—A Retrospective View" Insects 13, no. 2: 206. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020206