
Identification of a Cordyceps fumosorosea Fungus Isolate and Its Pathogenicity against Asian Citrus Psyllid, Diaphorina citri (Hemiptera: Liviidae)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Isolation of Entomopathogenic Fungus
2.2. Morphological Characterization and Inoculation
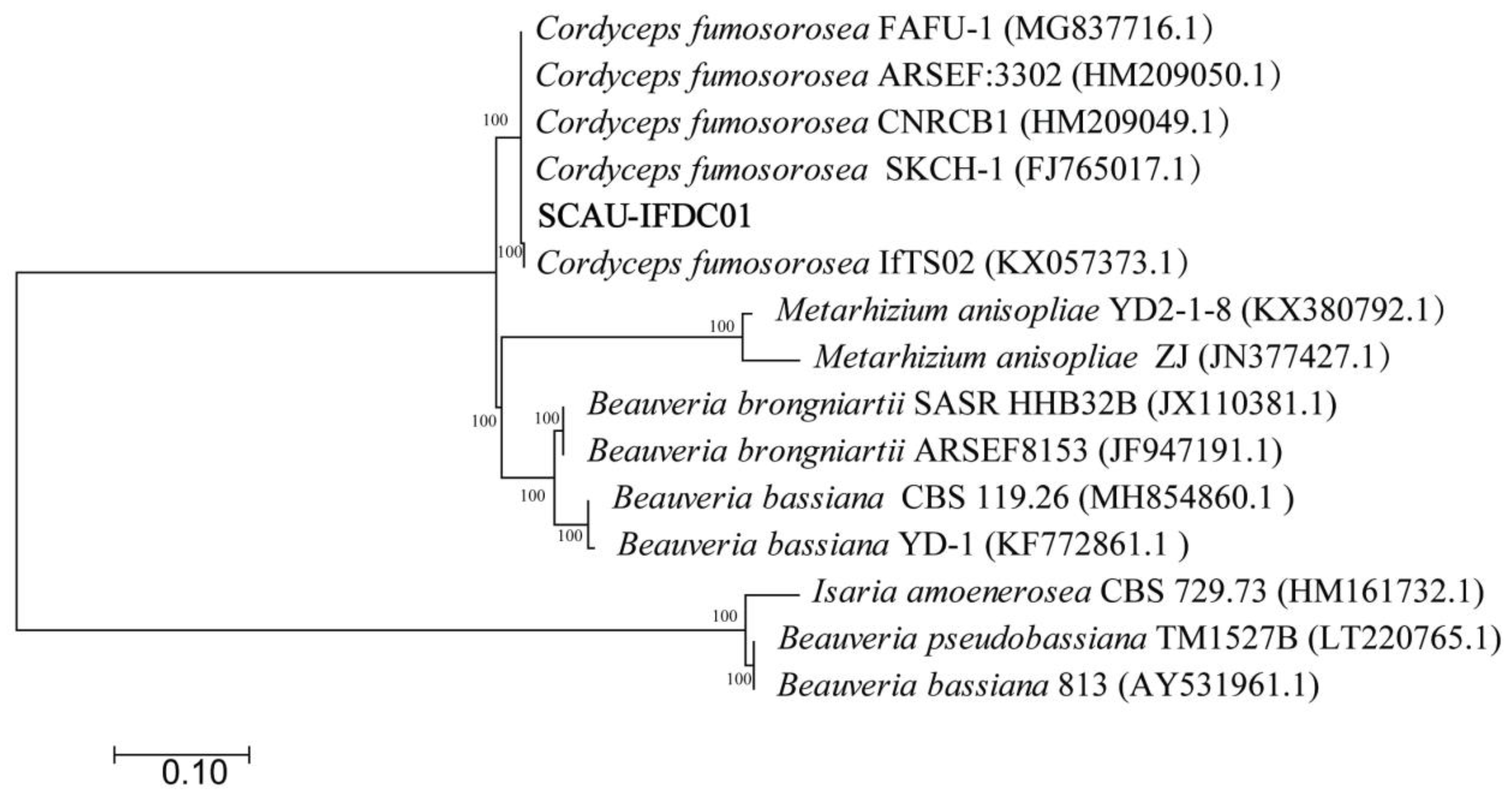
2.3. Indentification and Phylogenetic Tree of Fungal Isolate
2.4. Host Plants and Insect Colony
2.5. Laboratory Bioassays
2.6. Greenhouse Bioassays
2.7. Statistical Analyses
3. Results
3.1. Identification and Characterization of the Isolate
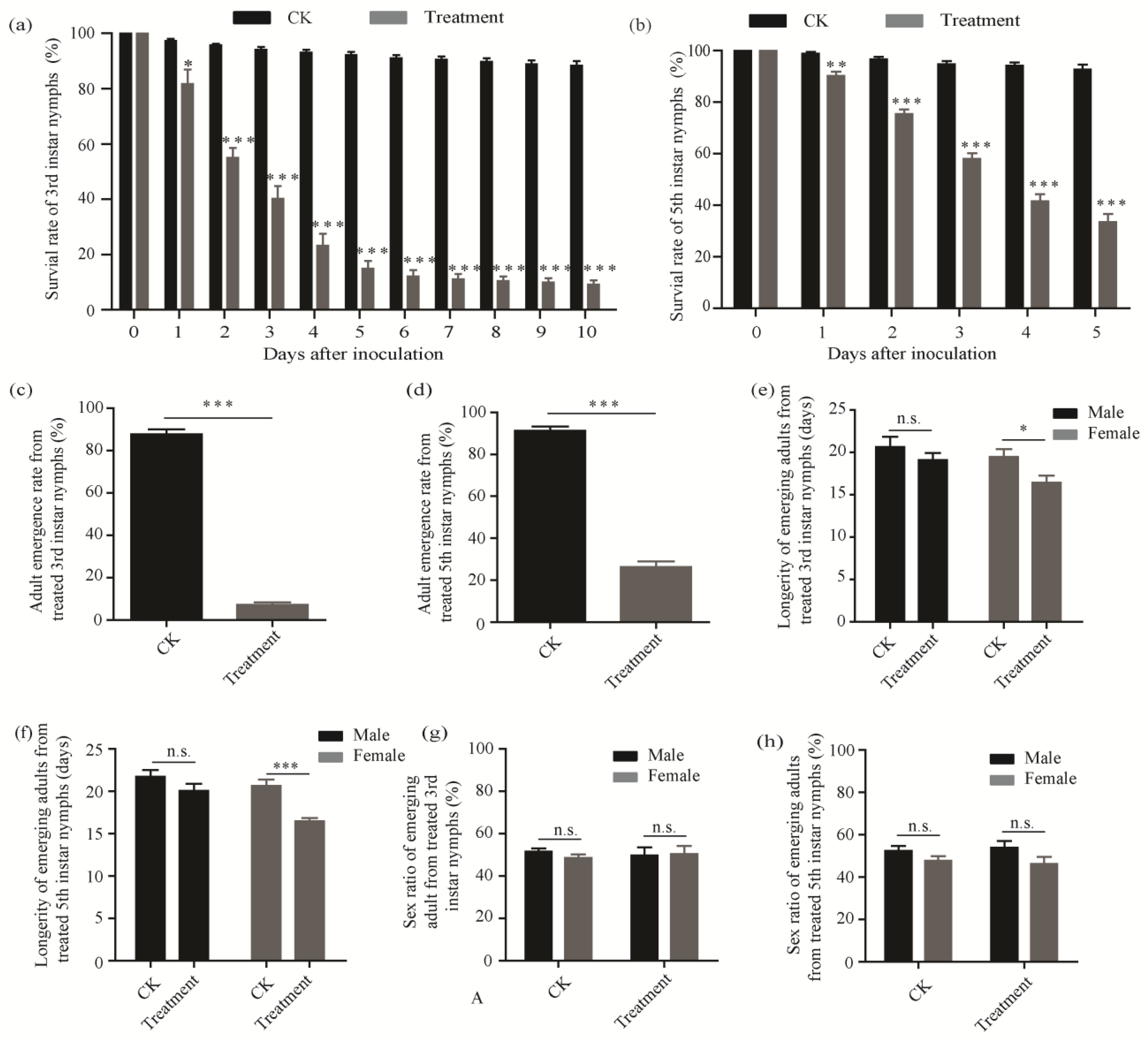
3.2. Pathogenicity of C. fumosorosea SCAU-CFDC01 against D. citri
3.3. Greenhouse Experiments
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bové, J.M. Huanglongbing: A destructive newly-emerging, century-old disease of citrus. J. Plant Path. 2006, 88, 7–37. [Google Scholar]
- Bassanezi, R.B.; Montesino, L.H.; Gasparoto, M.C.G.; Filho, A.B.; Amorim, L. Yield loss caused by huanglongbing in different sweet orange cultivars in so paulo, brazil. Eur. J. Plant Pathol. 2011, 130, 577–586. [Google Scholar] [CrossRef]
- Graa, J.V.D.; Douhan, G.W.; Halbert, S.E.; Keremane, M.L.; Lee, R.F.; Vidalakis, G.; Zhao, H. Huanglongbing: An overview of a complex pathosystem ravaging the world’s citrus. J. Integr. Plant Biol. 2016, 58, 373–387. [Google Scholar]
- Wang, N.; Pierson, E.A.; Setubal, J.C.; Xu, J.; Levy, J.G.; Zhang, Y.Z.; Li, J.Y.; Rangel, L.T.; Martins, J. The Candidatus Liberibacter-host interferface: Insights into pathogenesis mechanisms and disease control. Annu. Rev. Phytopathol. 2017, 55, 451–482. [Google Scholar] [CrossRef] [PubMed]
- Hall, D.G.; Richardson, M.L.; Ammar, E.D.; Halbert, S.E. Asian citrus psyllid, Diaphorina citri, vector of citrus huanglongbing disease. Entomol. Exp. Appl. 2013, 146, 207–223. [Google Scholar] [CrossRef]
- Yang, Y.P.; Huang, M.D.; Beattie, G.A.C.; Xia, Y.L.; Ouyang, G.C.; Xiong, J.J. Distribution, biology, ecology and control of the psyllid Diaphorina citri Kuwayama, a major pest of citrus: A status report for China. Int. J. Pest Manag. 2006, 552, 343–352. [Google Scholar] [CrossRef]
- Boina, D.R.; Onagbola, E.O.; Salyani, M.; Stelinski, L.L. Antifeedant and sublethal effects of imidacloprid on Asian citrus psyllid, Diaphorina citri. Pest Manag. Sci. 2009, 65, 870–877. [Google Scholar] [CrossRef]
- Huang, J.K.; Hu, R.F.; Pray, C.; Qiao, F.B.; Rozelle, S. Biotechnology as an alternative to chemical pesticides: A case study of Bt cotton in China. Agric. Econ. 2015, 29, 55–67. [Google Scholar] [CrossRef]
- Pardo, S.; Martinez, A.M.; Figueroa, J.I.; Chavarrieta, J.M.; Vinuela, E.; Rebollar-Alviter, A.; Miranda, M.A.; Valle, J.; Pineda, S. Insecticide resistance of adults and nymphs of Asian citrus psyllid populations from Apatzingan Valley, Mexico. Pest Manag. Sci. 2018, 74, 34–47. [Google Scholar] [CrossRef]
- Xiong, Q.; Xie, Y.P.; Zhu, Y.M.; Xue, J.L.; Li, J.; Fan, R.J. Morphological and ultrastructural characterization of Carposina sasakii larvae (Lepidoptera: Carposinidae) infected by Beauveria bassiana (Ascomycota: Hypocreales: Clavicipitaceae). Micron 2013, 44, 303–311. [Google Scholar] [CrossRef]
- Wang, C.S.; Wang, S.B. Insect pathogenic fungi: Genomics, molecular interactions, and genetic improvements. Ann. Rev. Entomol. 2017, 62, 73–90. [Google Scholar] [CrossRef] [PubMed]
- Wei, G.; Lai, Y.L.; Wang, G.D.; Chen, H.; Li, F.; Wang, S.B. Insect pathogenic fungus interacts with the gut microbiota to accelerate mosquito mortality. Proc. Natl. Acad. Sci. USA 2017, 114, 5994–5999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shipp, J.L.; Zhang, Y.; Hunt, D.W.A.; Ferguson, G. Influence of humidity and greenhouse microclimate on the efficacy of Beauveria bassiana (Balsamo) for control of greenhouse arthropod pests. Environ. Entomol. 2003, 32, 1154–1163. [Google Scholar] [CrossRef] [Green Version]
- Stauderman, K.; Avery, P.; Aristizábal, L.; Arthurs, S. Evaluation of Isaria fumosorosea (Hypocreales: Cordycipitaceae) for control of the Asian citrus psyllid, Diaphorina citri (Hemiptera: Psyllidae). Biocontrol Sci. Technol. 2012, 22, 747–761. [Google Scholar] [CrossRef]
- Lezama-Gutiérrez, R.; Molina-Ochoa, J.; Chávez-Flores, O.; Ángel-Sahagún, C.A.; Skoda, S.R.; Reyes-Martínez, G.; Barba-Reynoso, M.; Rebolledo-Domínguez, O.; Ruíz-Aguilar, G.M.L.; Foster, J.E. Use of the entomopathogenic fungi Metarhizium anisopliae, Cordyceps bassiana and Isaria fumosorosea to control Diaphorina citri (hemiptera: Psyllidae) in persian lime under field conditions. Int. J. Trop. Insect Sci. 2012, 32, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Orduño-Cruz, N.; Guzmán-Franco, A.W.; Rodríguez-Leyva, E.; Alatorre-Rosas, R.; González-Hernández, H.; Mora-Aguilera, G. In vivo selection of entomopathogenic fungal isolates for control of Diaphorina citri (Hemiptera: Liviidae). Biol. Control. 2015, 90, 1–5. [Google Scholar] [CrossRef]
- Ou, D.; Zhang, L.H.; Guo, C.F.; Chen, X.S.; Ali, S.; Qiu, B.L. Identifification of a new Cordyceps javanica fungus isolate and its toxicity evaluation against Asian citrus psyllid. MicrobiologyOpen 2019, 8, e00760. [Google Scholar] [CrossRef] [Green Version]
- Awan, U.A.; Meng, L.X.; Xia, S.; Raza, M.F.; Zhang, Z.Y.; Zhang, H.Y. Isolation, fermentation, and formulation of entomopathogenic fungi virulent against adults of Diaphorina citri. Pest Manag. Sci. 2021, 77, 4040–4053. [Google Scholar] [CrossRef]
- Gallou, A.; Serna-Domínguez, M.G.; Berlanga-Padilla, A.M.; Ayala-Zermeño, M.A.; Mellín-Rosas, M.A.; Montesinos-Matías, R.; Arredondo-Bernal, H.C. Species clarification of Isaria isolates used as biocontrol agents against Diaphorina citri (Hemiptera: Liviidae) in Mexico. Fungal Biol. 2016, 120, 414–423. [Google Scholar] [CrossRef]
- Ibarra-Cortes, K.H.; Guzman-Franco, A.W.; Gonzalez-Hernandez, H.; Ortega-Arenas, L.D.; Villanueva-jimenez, J.A.; Robles-bermudez, A. Susceptibility of Diaphorina citri (Hemiptera: Liviidae) and its parasitoid Tamarixia radiata (Hymenoptera: Eulophidae) to entomopathogenic fungi under laboratory conditions. Neotrop. Entomol. 2018, 47, 131–138. [Google Scholar] [CrossRef]
- Humber, R.A. Fungi: Identification. In Manual of Techniques in Insect Pathology; Lacey, L.A., Ed.; Academic Press: London, UK, 1997. [Google Scholar]
- Humber, R.A. Identification of entomopathogenic fungi. In Manual of Techniques in Invertebrate Pathology, 2nd ed.; Lacey, L.A., Ed.; Academic Press: San Diego, CA, USA, 2012. [Google Scholar]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Finney, D.J. Probit Analysis: A Statistical Treatment of the Sigmoid Response Curve, 3rd ed.; Cambridge University Press: Cambridge, UK, 1972. [Google Scholar]
- Grafton-Cardwell, E.E.; Stelinski, L.L.; Stansly, P.A. Biology and management of Asian citrus psyllid, vector of the huanglongbing pathogens. Annu. Rev. Entomol. 2013, 58, 413–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoy, M.A.; Singh, R.; Rogers, M.E. Evaluations of a novel isolate of Isaria fumosorosea for control of the Asian citrus psyllid, Diaphorina citri (Hemiptera: Psyllidae). Fla. Entomol. 2010, 93, 24–32. [Google Scholar] [CrossRef]
- Pinto, A.P.F.; Filho, A.B.; Almeida, J.E.M.D.; Wenzel, I.M. Beauveria bassiana pathogenicity to Diaphorina citri and compatibility of the fungus with phytosanitary products. Pesqui. Agropecuária Bras. 2012, 47, 1673–1680. [Google Scholar] [CrossRef] [Green Version]
- Bugeme, D.M.; Maniania, N.K.; Knapp, M.; Boga, H.I. Effect of temperature on virulence of Beauveria bassiana and Metarhizium anisopliae isolates to Tetranychus evansi. Exp. Appl. Acarol. 2008, 46, 275–285. [Google Scholar] [CrossRef]
- Erler, F.; Ates, A.O. Potential of two entomopathogenic fungi, Beauveria bassiana and Metarhizium anisopliae (Coleoptera: Scarabaeidae), as biological control agents against the June beetle. J. Insect Sci. 2015, 15, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández, M.; Miranda, I. Comportamiento de Diaphorina citri Kuwayama (Hemiptera: Psyllidae). Parte I: Características morfológicas, incidencia y enemigos naturales asociados. Catheter. Cardiovasc. Interv. 2005, 2, 27–31. [Google Scholar]
- Avery, P.B.; Wekesa, V.W.; Hunter, W.B.; Hall, D.G.; McKensie, C.L.; Osborne, L.S.; Powell, C.A.; Rogers, M.E. Effects of the fungus Isaria fumosorosea (Hypocreales: Cordycipitaceae) on reduced feeding and mortality of the Asian citrus psyllid, Diaphorina citri (Hemiptera: Psyllidae). Biocontrol Sci. Technol. 2011, 21, 1065–1078. [Google Scholar] [CrossRef]
- Cortez-Madrigal, H.J.; Sánchez-Saavedra, M.; Díaz-Godínez, G.; MoraAguilera, G. Enzymatic activity and pathogenicity of entomopathogenic fungi from central and southeastern Mexico to Diaphorina citri (Hemiptera: Psyllidae). Southwest. Entomol. 2014, 39, 491–502. [Google Scholar] [CrossRef]
- Bamisile, B.S.; Dash, C.K.; Akutse, K.S.; Qasim, M.; Ramos Aguila, L.C.; Wang, F.; Keppanan, R.; Wang, L. Endophytic Beauveria bassiana in Foliar-treated citrus limon plants acting as a growth suppressor to three successive generations of Diaphorina citri Kuwayama (Hemiptera: Liviidae). Insects 2019, 10, 176. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Wu, Q.; Fan, J.; Huang, J.; Wu, Z.; Lin, J.; Bin, S.; Shu, B. Effects of the entomopathogenic fungus Clonostachys rosea on mortality rates and gene expression profiles in Diaphorina citri adults. J. Invertebr. Pathol. 2021, 179, 107539. [Google Scholar] [CrossRef] [PubMed]
- Gandarilla-Pacheco, F.L.; Galán-Wong, L.J.; López-Arroyo, J.I.; Rodríguez-Guerra, R.; Quintero-Zapata, I. Optimization of pathogenicity tests for selection of native isolates of entomopathogenic fungi isolated from citrus-growing areas of México on adults of Diaphorina ciri Kuwayama (Hemiptera: Liviidae). Fla. Entomol. 2013, 96, 187–195. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | GenBank Accession Number | Isolate Number | Location | Number of Nucleotides Analyzed (bp) |
|---|---|---|---|---|
| C. fumosorosea | OL872288.1 | SCAU-CFDC01 | Guangzhou, China | 564 |
| C. fumosorosea | MG837716 | FAFU-1 | Fuzhou, China | 603 |
| C. fumosorosea | HM209050.1 | ARSEF:3302 | Mexico | 570 |
| C. fumosorosea | HM209049.1 | CNRCB1 | Mexico | 572 |
| C. fumosorosea | FJ765017.1 | SKCH-1 | Guangzhou, China | 635 |
| C. fumosorosea | KX057373.1 | IfTS02 | Guangzhou, China | 564 |
| M. anisopliae | KX380792.1 | YD2-1-8 | Nanchang, China | 559 |
| M. anisopliae | JN377427.1 | zj | Wenchang, China | 685 |
| B. brongniartii | JX110381.1 | SASR HHB32B | South Africa | 595 |
| B. brongniartii | JF947191.1 | ARSEF8153 | Canada | 593 |
| B. bassiana | MH854860.1 | CBS 119.26 | Indonesia | 637 |
| B. bassiana | KF772861.1 | YD-1 | Wuhan, Hubei | 491 |
| B. bassiana | AY531961.1 | 813 | USA | 797 |
| Beauveria pseudobassiana | LT220765.1 | TM1527B | Portugal | 958 |
| I. amoenerosea | HM161732.1 | CBS729.73 | Thailand | 865 |
| Developmental Stage | Days | Slop ± SE | Regression Virulence Model | LC50 (Conidia mL−1) (95% CI) | p * |
|---|---|---|---|---|---|
| 3rd instar nymphs | 3 | 0.66 ± 0.09 | Y = 0.66X − 2.51 | 6.0 × 103 (2.37 × 103–1.25 × 104) | 0.65 |
| 4 | 0.72 ± 0.11 | Y = 0.72X − 2.49 | 3.00 × 103 (7.73–1.86 × 104) | 0.06 | |
| 5 | 1.03 ± 0.16 | Y = 1.03X − 3.46 | 2.32 × 103 (1.10 × 103–4.08 × 103) | 0.72 | |
| 6 | 1.16 ± 0.19 | Y = 1.16X − 3.77 | 1.82 × 103 (8.94 × 102–3.09 × 103) | 0.25 | |
| 7 | 0.99 ± 0.18 | Y = 0.99X − 3.03 | 1.14 × 103 (1.61–4.63 × 103) | 0.11 | |
| 5th instar nymphs | 3 | 0.56 ± 0.28 | Y = 0.56X − 2.84 | 1.14 × 105 (2.15 × 104–8.04 × 105) | 0.17 |
| 4 | 0.86 ± 0.26 | Y = 0.86X − 4.13 | 6.55 × 104 (1.97 × 103–2.15 × 105) | 0.17 | |
| 5 | 1.63 ± 0.42 | Y = 1.63X − 7.28 | 2.86 × 104 (6.31 × 103–5.84 × 104) | 0.51 | |
| 6 | 1.74 ± 0.53 | Y = 1.74X − 7.92 | 3.46 × 104 (4.76 × 103–7.23 × 104) | 0.54 | |
| 7 | 1.57 ± 0.44 | Y = 1.57X − 6.98 | 2.88 × 104 (4.08 × 103–6.42 × 104) | 0.44 | |
| Adult | 3 | 0.32 ± 0.08 | Y = 0.32X − 2.36 | 2.68 × 107 (3.25 × 106–6.73 × 109) | 0.96 |
| 4 | 0.30 ± 0.08 | Y = 0.30X − 2.18 | 1.85 × 107 (2.26 × 106–5.01 × 109) | 0.98 | |
| 5 | 0.27 ± 0.08 | Y = 0.27X − 1.84 | 5.43 × 106 (7.73 × 105–7.51 × 108) | 0.87 | |
| 6 | 0.27 ± 0.08 | Y = 0.27X − 1.82 | 4.76 × 106 (6.89 × 105–6.02 × 108) | 0.94 | |
| 7 | 0.26 ± 0.08 | Y = 0.26X − 1.70 | 4.69 × 106 (6.38 × 105–8.82 × 108) | 0.95 |
| Developmental Stage | Conidia Concentration | Slop ± SE | Regression Model | LT50 (days) (95% CI) | p |
|---|---|---|---|---|---|
| 3rd instar nymphs | 1 × 107 | 5.71 ± 1.10 | Y = 5.71X − 0.36 | 1.18 (0.94–1.35) | 0.09 |
| 1 × 106 | 4.55 ± 0.65 | Y = 4.55X − 0.85 | 1.53 (1.26–1.78) | 0.95 | |
| 1 × 105 | 4.20 ± 0.38 | Y = 4.23X − 1.11 | 1.75 (1.54–1.94) | 0.91 | |
| 1 × 104 | 2.01 ± 0.32 | Y = 2.06X − 1.06 | 3.28 (2.69–3.94) | 0.64 | |
| 1 × 103 | 1.58 ± 0.30 | Y = 1.58X − 1.33 | 6.93 (5.40–10.85) | 0.10 | |
| 5th instar nymphs | 1 × 107 | 6.35 ± 0.58 | Y = 6.35X − 1.14 | 1.51 (1.39–1.63) | 0.31 |
| 1 × 106 | 4.78 ± 0.39 | Y = 4.78X − 1.58 | 2.14 (1.75–2.51) | 0.06 | |
| 1 × 105 | 3.32 ± 0.32 | Y = 3.32X − 1.42 | 2.67 (2.34–2.97) | 0.40 | |
| 1 × 104 | 2.10 ± 0.29 | Y = 2.10X − 1.48 | 5.09 (4.39–6.17) | 0.77 | |
| 1 × 103 | 1.75 ± 0.31 | Y = 1.75X − 1.54 | 7.60 (6.03–11.43) | 0.17 | |
| Adults | 1 × 108 | 1.61 ± 0.58 | Y = 1.61X − 0.13 | 1.20 (0.00–3.67) | 0.74 |
| 5 × 107 | 1.57 ± 0.64 | Y = 1.57X − 0.56 | 2.28 (0.00–5.89) | 0.97 | |
| 1 × 107 | 1.63 ± 0.62 | Y = 1.63X − 0.96 | 3.88 (0.14–6.99) | 0.88 | |
| 5 × 106 | 1.24 ± 0.54 | Y = 1.24X − 1.01 | 6.51 (0.04–13.78) | 0.83 | |
| 1 × 106 | 1.28 ± 0.83 | Y = 1.28X − 1.26 | 9.71 (0.21–15.78) | 0.98 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, Y.; Wu, S.; He, X.; Wang, D.; He, Y.; Nian, X. Identification of a Cordyceps fumosorosea Fungus Isolate and Its Pathogenicity against Asian Citrus Psyllid, Diaphorina citri (Hemiptera: Liviidae). Insects 2022, 13, 374. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13040374
Luo Y, Wu S, He X, Wang D, He Y, Nian X. Identification of a Cordyceps fumosorosea Fungus Isolate and Its Pathogenicity against Asian Citrus Psyllid, Diaphorina citri (Hemiptera: Liviidae). Insects. 2022; 13(4):374. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13040374
Chicago/Turabian StyleLuo, Yaru, Shujie Wu, Xinyu He, Desen Wang, Yurong He, and Xiaoge Nian. 2022. "Identification of a Cordyceps fumosorosea Fungus Isolate and Its Pathogenicity against Asian Citrus Psyllid, Diaphorina citri (Hemiptera: Liviidae)" Insects 13, no. 4: 374. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13040374