
Effects of Larval Diet on the Male Reproductive Traits in the West Indian Sweet Potato Weevils Euscepes postfasciatus (Coleoptera: Curculionidae)

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Insects
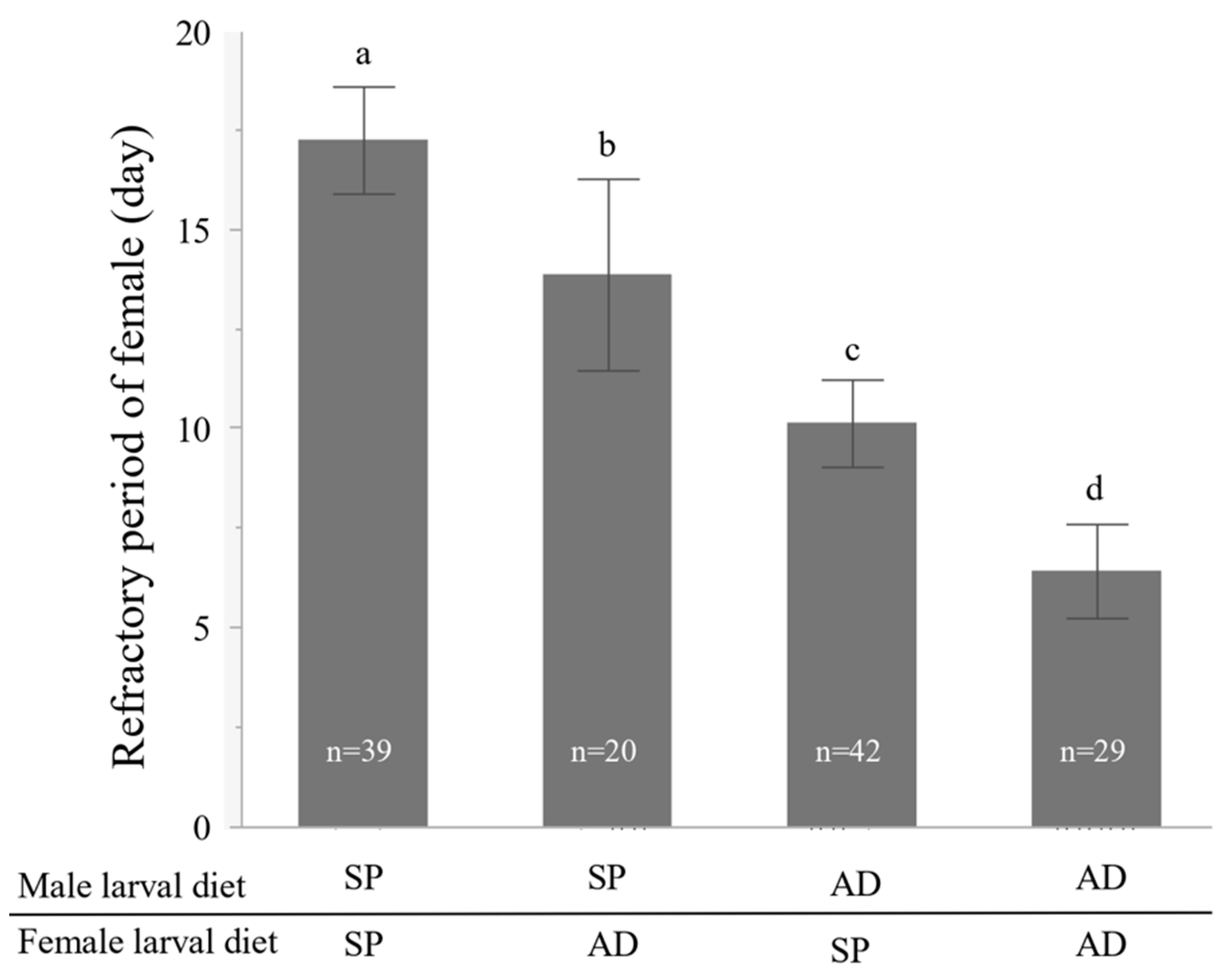
2.2. Experiment 1. Effects of the Larval Diet of Males and Females on the Refractory Period of Females
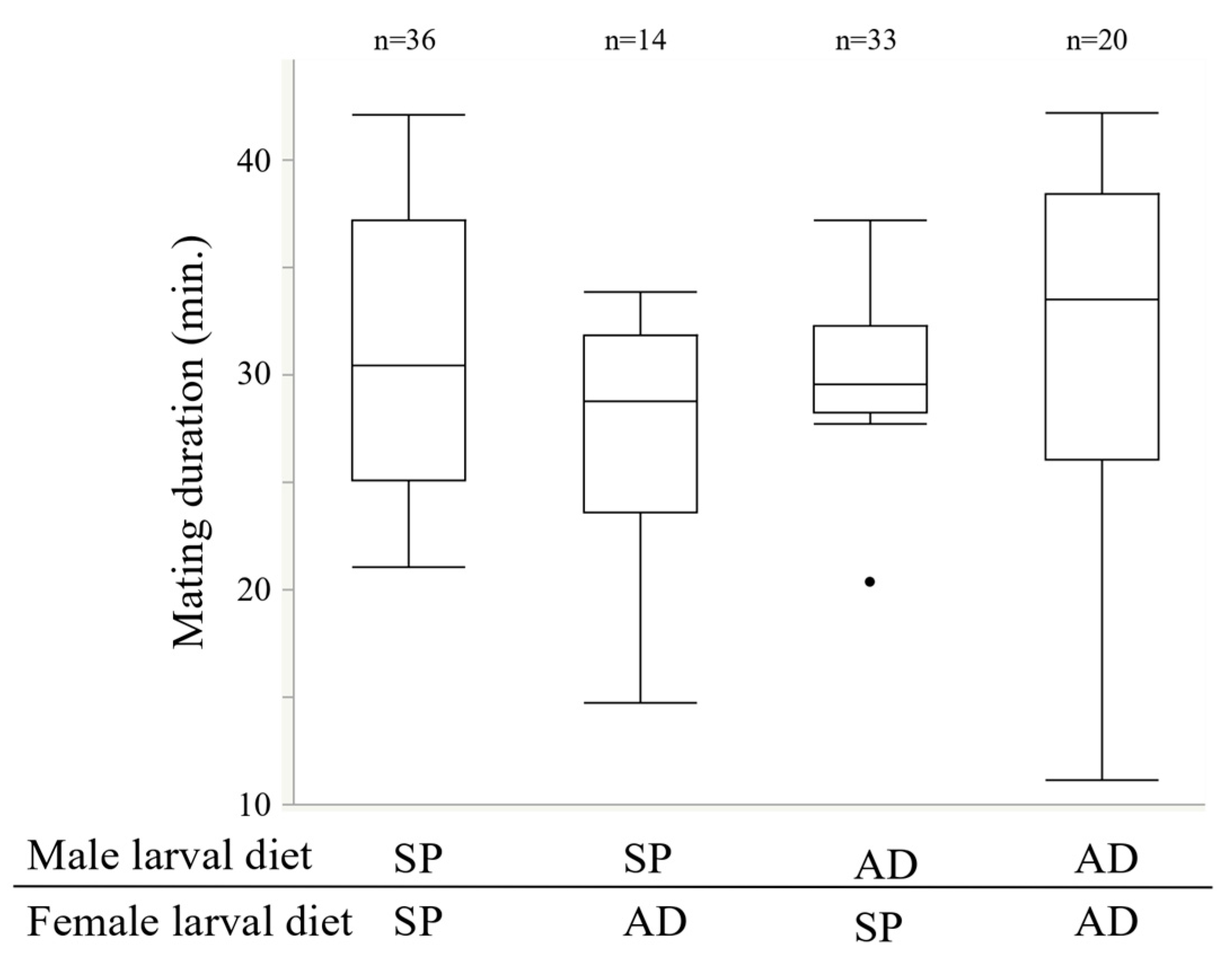
2.3. Experiment 2. Effects of the Larval Diet on Male Reproductive Traits
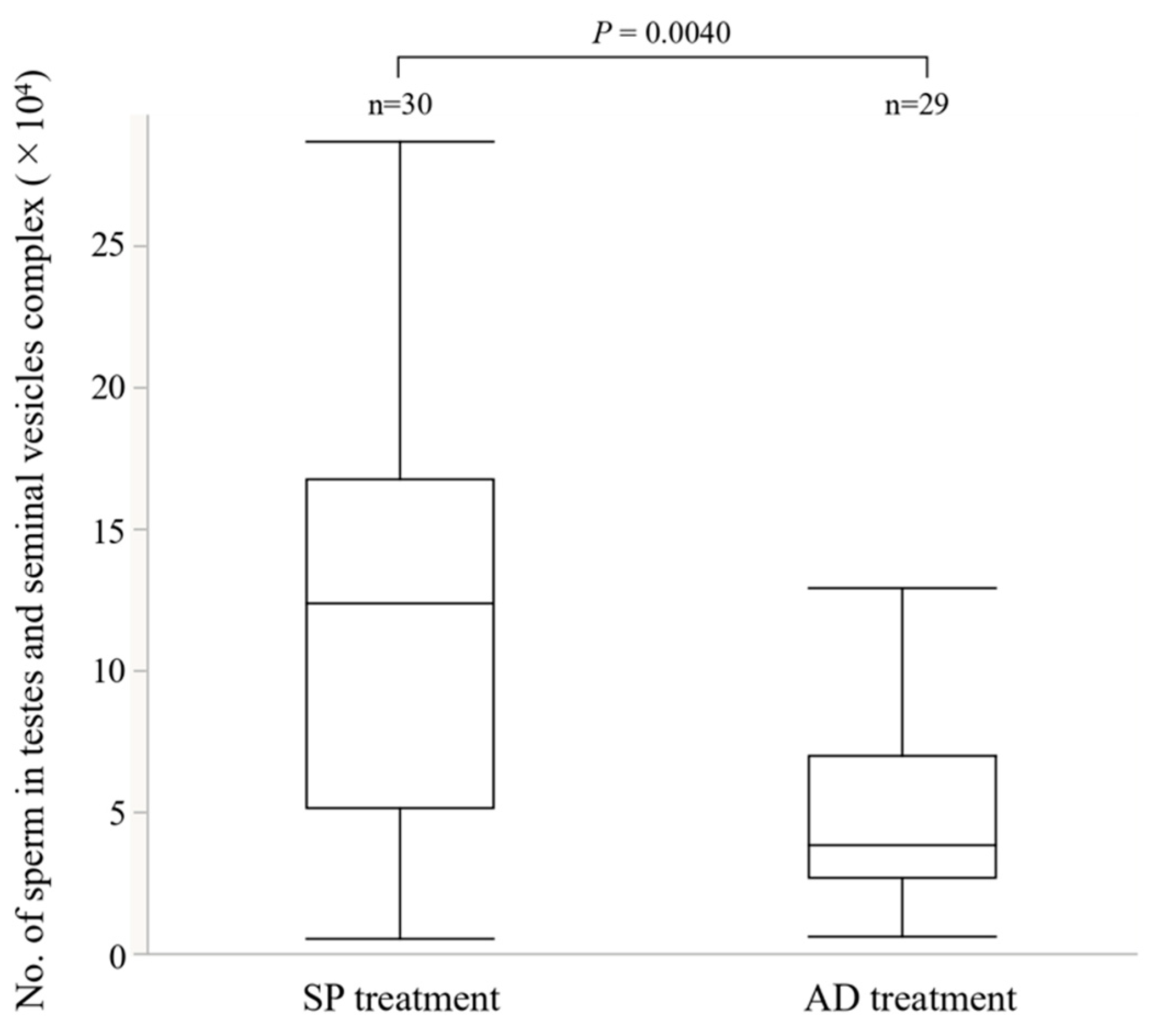
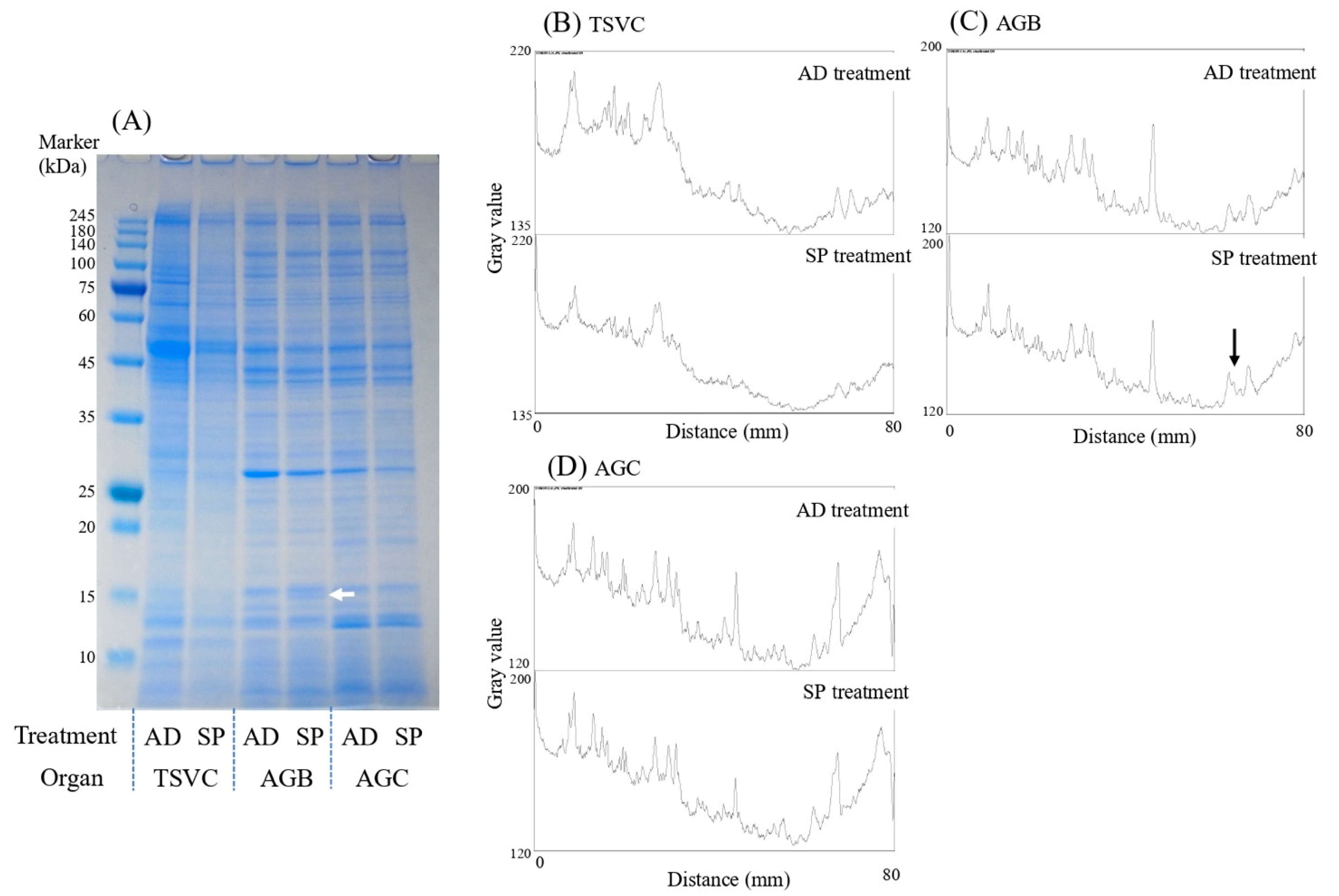
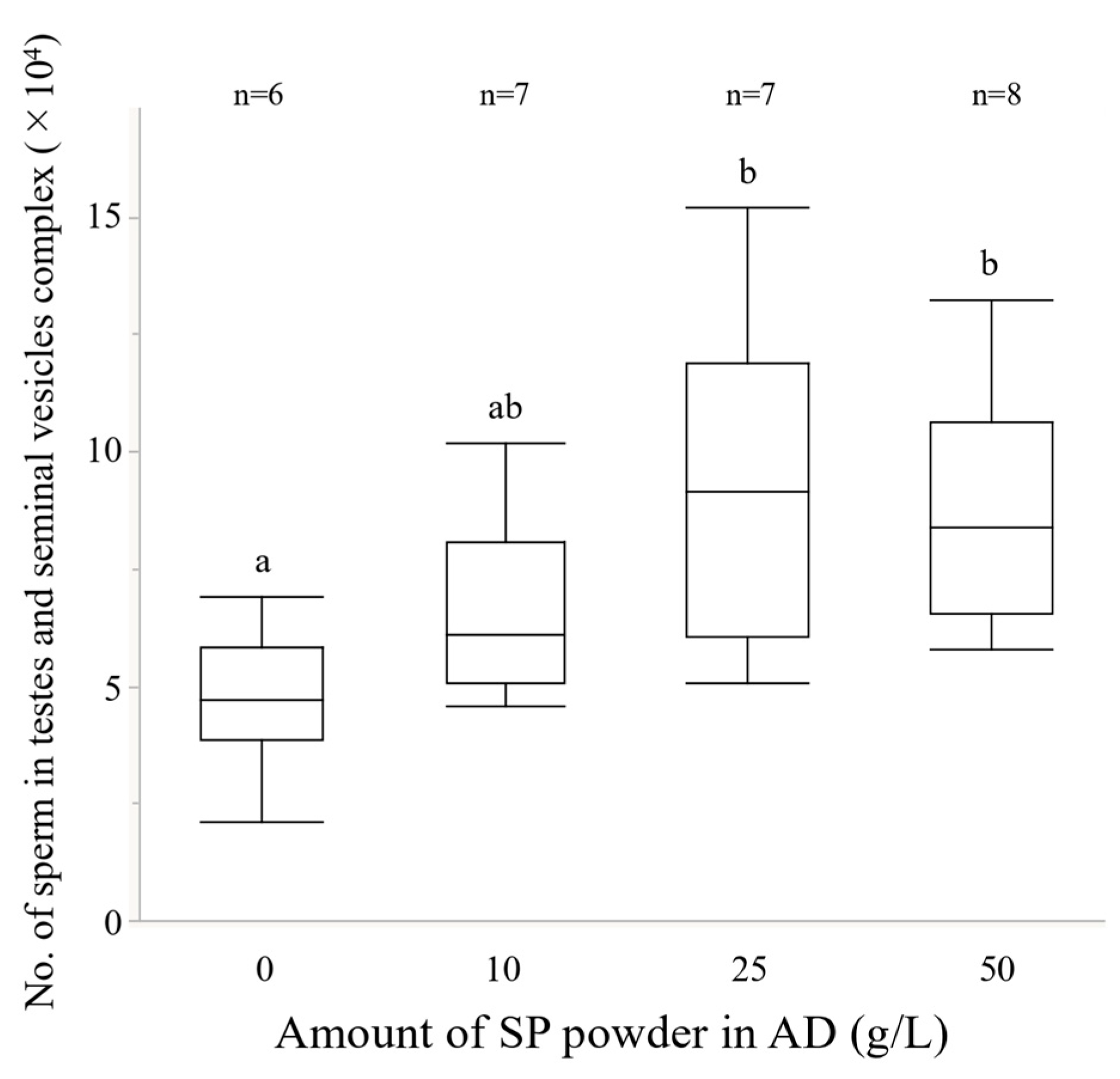
2.3.1. Number of Sperm in the Testes-Seminal Vesicles Complex
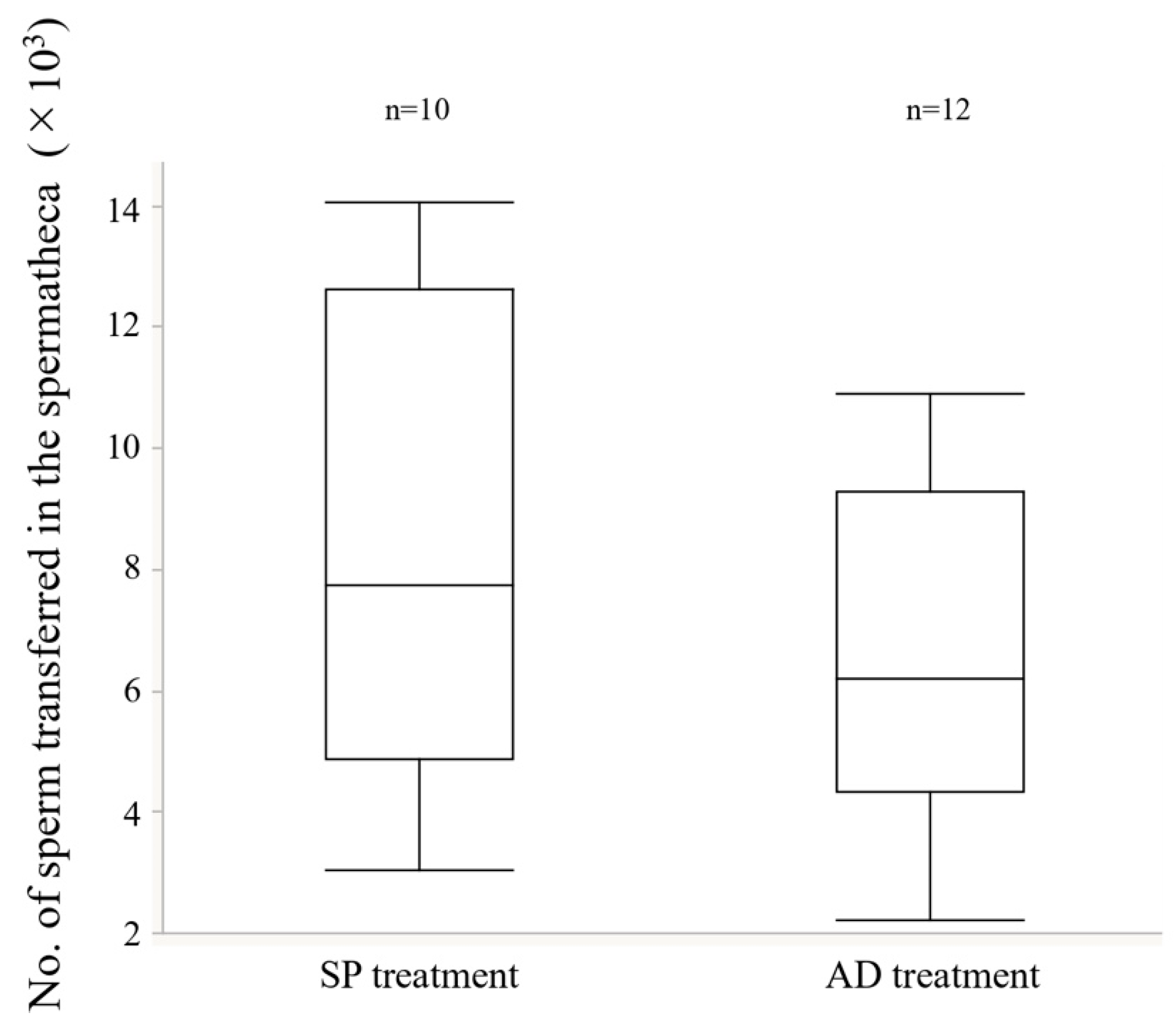
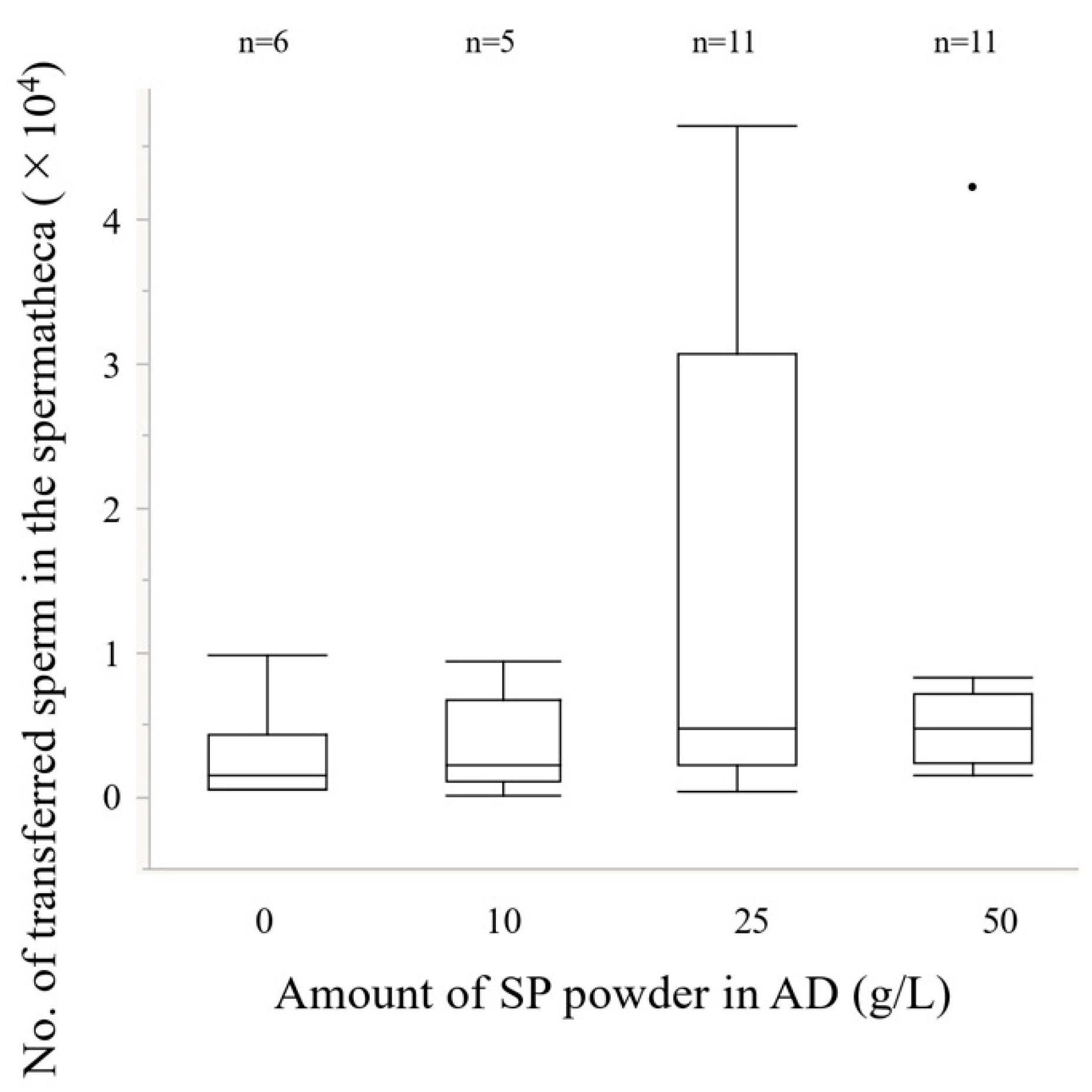
2.3.2. Number of Transferred Sperm into the Spermatheca
2.3.3. Seminal Fluid Substances
SDS-PAGE
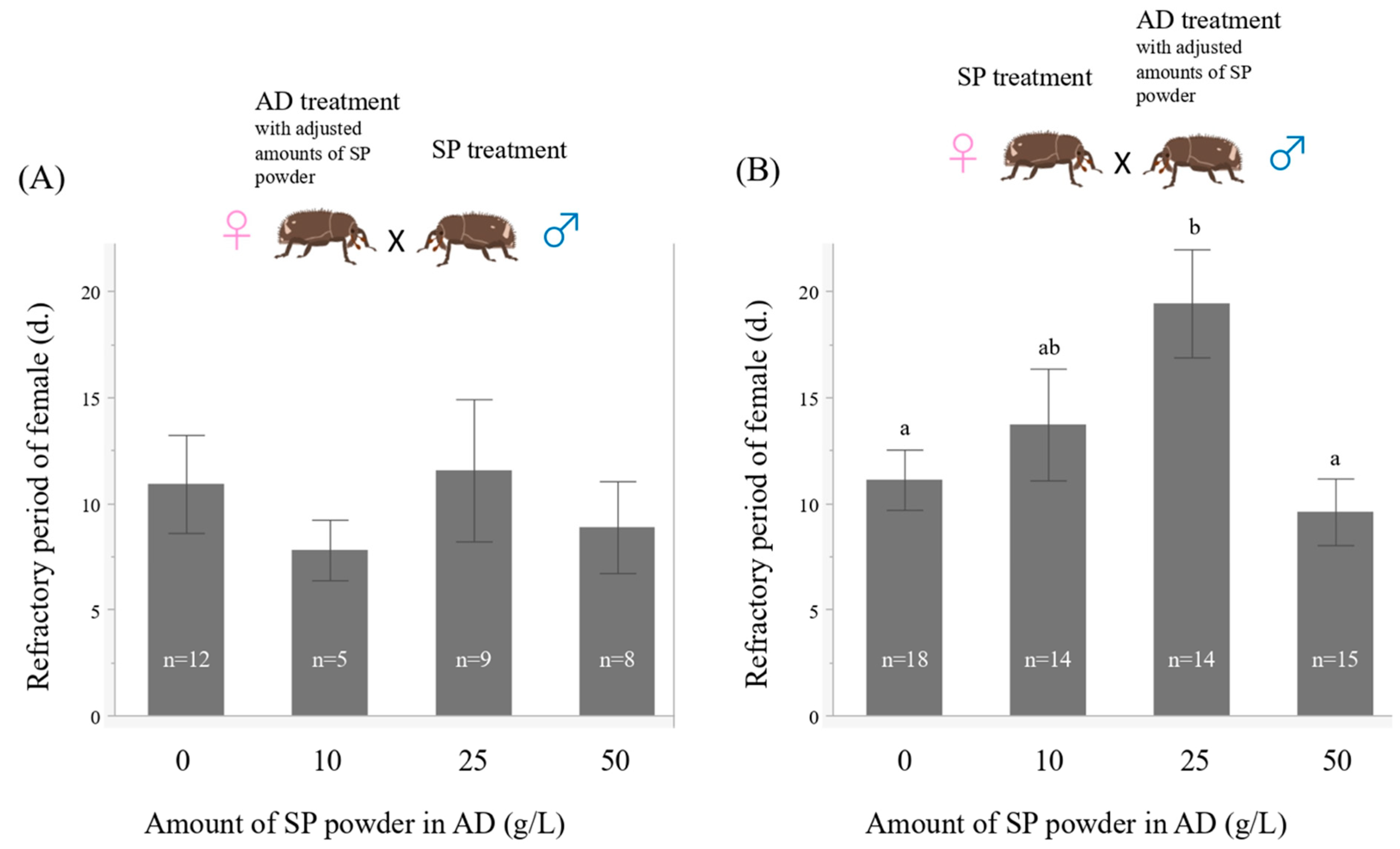
2.4. Experiment 3. Effects of the Amount of Sweet Potato Powder in the Artificial Diet on the Refractory Period of Females
2.5. Experiment 4. Effects of the Amount of Sweet Potato Powder in the Artificial Diet on Reproductive Traits
2.6. Statistics
3. Results
3.1. Experiment 1. Effects of the Larval Diet of Males and Females on the Refractory Period of Females
3.2. Experiment 2. Effects of the Larval Diet on Male Reproductive Traits
3.3. Experiment 3. Effects of the Amount of Sweet Potato Powder in the Artificial Diet on the Refractory Period of Females
3.4. Experiment 4. Effects of the Amount of Sweet Potato Powder in the Artificial Diet on Reproductive Traits
4. Discussion
4.1. Effects of Larval Diet on the Female Refractory Period
4.2. Effects of Larval Diet on the Reproductive Traits
4.3. Effects of the Amount of Sweet Potato Powder in the Artificial Diet on the Refractory Period of Females
4.4. Application Aspects
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yamasaki, A.; Shimizu, K.; Fujisaki, K. Effect of host plant part on larval body-color polymorphism in Helicoverpa armigera (Lepidoptera: Noctuidae). Ann. Entomol. Soc. Am. 2009, 102, 76–84. [Google Scholar] [CrossRef] [Green Version]
- Gobbi, P.; Martínez-Sánchez, A.; Rojo, S. The effects of larval diet on adult life-history traits of the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae). Eur. J. Entomol. 2013, 110, 461–468. [Google Scholar] [CrossRef]
- Swanson, W.J.; Clark, A.G.; Waldrip-Dail, H.M.; Wolfner, M.F.; Aquadro, C.F. Evolutionary EST analysis identifies rapidly evolving male reproductive proteins in Drosophila. Proc. Natl. Acad. Sci. USA 2001, 98, 7375–7379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravi Ram, K.; Wolfner, M.F. Seminal influences: Drosophila Acps and the molecular interplay between males and females during reproduction. Integr. Comp. Biol. 2007, 47, 427–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Findlay, G.D.; Yi, X.; MacCoss, M.J.; Swanson, W.J. Proteomics reveals novel Drosophila seminal fluid proteins transferred at mating. PLoS Biol. 2008, 6, e178. [Google Scholar] [CrossRef]
- Hopkins, B.R.; Sepil, I.; Thézénas, M.L.; Craig, J.F.; Miller, T.; Charles, P.D.; Fischer, R.; Kessler, B.M.; Bretman, A.; Pizzari, T.; et al. Divergent allocation of sperm and the seminal proteome along a competition gradient in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2019, 116, 17925–17933. [Google Scholar] [CrossRef] [Green Version]
- Sepil, I.; Hopkins, B.R.; Dean, R.; Thézénas, M.L.; Charles, P.D.; Konietzny, R.; Fischer, R.; Kessler, B.M.; Wigby, S. Quantitative proteomics identification of seminal fluid proteins in male Drosophila melanogaster. Mol. Cell. Proteom. 2019, 18, S46–S58. [Google Scholar] [CrossRef] [Green Version]
- Dean, M.D.; Clark, N.L.; Findlay, G.D.; Karn, R.C.; Yi, X.; Swanson, W.J.; MacCoss, M.J.; Nachman, M.W. Proteomics and comparative genomic investigations reveal heterogeneity in evolutionary rate of male reproductive proteins in mice (Mus domesticus). Mol. Biol. Evol. 2009, 26, 1733–1743. [Google Scholar] [CrossRef] [Green Version]
- Baer, B.; Heazlewood, J.L.; Taylor, N.L.; Eubel, H.; Millar, A.H. The seminal fluid proteome of the honeybee Apis mellifera. Proteomics 2009, 9, 2085–2097. [Google Scholar] [CrossRef]
- Pilch, B.; Mann, M. Large-scale and high-confidence proteomic analysis of human seminal plasma. Genome Biol. 2006, 7, R40. [Google Scholar] [CrossRef] [Green Version]
- Degner, E.C.; Ahmed-Braimah, Y.H.; Borziak, K.; Wolfner, M.F.; Harrington, L.C.; Dorus, S. Proteins, transcripts, and genetic architecture of seminal fluid and sperm in the mosquito Aedes aegypti. Mol. Cell. Proteom. 2019, 18, S6–S22. [Google Scholar] [CrossRef] [PubMed]
- Sirot, L.K.; Hardstone, M.C.; Helinski, M.E.; Ribeiro, J.M.; Kimura, M.; Deewatthanawong, P.; Wolfner, M.F.; Harrington, L.C. Towards a semen proteome of the dengue vector mosquito: Protein identification and potential functions. PLoS Negl. Trop. Dis. 2011, 5, e989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poiani, A. Complexity of seminal fluid: A review. Behav. Ecol. Sociobiol. 2006, 60, 289–310. [Google Scholar] [CrossRef]
- Avila, F.W.; Sirot, L.K.; LaFlamme, B.A.; Rubinstein, C.D.; Wolfner, M.F. Insect seminal fluid proteins: Identification and function. Annu. Rev. Entomol. 2011, 56, 21–40. [Google Scholar] [CrossRef] [Green Version]
- Sirot, L.K.; Wong, A.; Chapman, T.; Wolfner, M.F. Sexual conflict and seminal fluid proteins: A dynamic landscape of sexual interactions. Cold Spring Harb. Perspect. Biol. 2014, 7, a017533. [Google Scholar] [CrossRef]
- Drony, D.C. The influence of the nutritional content of the adult male diet on testis mass, body condition and courtship vigour in a Hawaiian Drosophila. Funct. Ecol. 1996, 12, 920–928. [Google Scholar] [CrossRef]
- Taylor, P.W.; Yuval, B. Postcopulatory sexual selection in mediterranean fruit flies: Advantages for large and protein-fed males. Anim. Behav. 1999, 58, 247–254. [Google Scholar] [CrossRef] [Green Version]
- Blay, S.; Yuval, B. Nutritional correlates of reproductive success of male Mediterranean fruit flies (Diptera: Tephritidae). Anim. Behav. 1997, 54, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Yuval, B.; Kaspi, R.; Field, S.A.; Blay, S.; Taylor, P. Effects of post-teneral nutrition on reproductive success of male Mediterranean fruit flies (Diptera: Tephritidae). Fla. Entomol. 2002, 85, 165–170. [Google Scholar] [CrossRef]
- Gavriel, S.; Gazit, Y.; Yuval, B. Remating by female Mediterranean fruit flies (Ceratitis capitata, Diptera: Tephritidae): Temporal patterns and modulation by male condition. J. Insect Physiol. 2009, 55, 637–642. [Google Scholar] [CrossRef]
- Pérez-Staples, D.; Aluja, M.; Macías-Ordóñez, R.; Sivinski, J. Reproductive trade-offs from mating with a successful male: The case of the tephritid fly Anastrepha obliqua. Behav. Ecol. Sociobiol. 2008, 62, 1333–1340. [Google Scholar] [CrossRef]
- Pérez-Staples, D.; Harmer, A.M.T.; Collins, S.R.; Taylor, P.W. Potential for pre-release diet supplements to increase the sexual performance and longevity of male Queensland fruit flies. Agric. For. Entomol. 2008, 10, 255–262. [Google Scholar] [CrossRef]
- Aluja, M.; Rull, J.; Sivinski, J.; Trujillo, G.; Pérez-Staples, D. Male and female condition influence mating performance and sexual receptivity in two tropical fruit flies (Diptera: Tephritidae) with contrasting life histories. J. Insect Physiol. 2009, 55, 1091–1098. [Google Scholar] [CrossRef] [PubMed]
- Gage, M.J.G.; Cook, P.A. Sperm size or numbers? Effects of nutritional stress upon eupyrene and apyrene sperm production strategies in the moth Plodia interpunctella (Lepidoptera: Pyralidea). Funct. Ecol. 1994, 8, 594–599. [Google Scholar] [CrossRef]
- Amitin, E.G.; Pitnick, S. Influence of developmental environment on male- and female-mediated sperm precedence in Drosophila melanogaster. J. Evol. Biol. 2007, 20, 381–391. [Google Scholar] [CrossRef]
- Hawley, W.A. The biology of Aedes albopictus. J. Am. Mosq. Control Assoc. Suppl. 1988, 1, 1–39. [Google Scholar] [PubMed]
- Clements, A.N. The Biology of Mosquitoes. Development, Nutrition, and Reproduction; Chapman & Hall: London, UK, 1992; p. 509. [Google Scholar]
- Delisle, J.; Bouchard, A. Male larval nutrition in Choristoneura rosaceana (Lepidoptera: Tortricidae): An important factor in reproductive success. Oecologia 1995, 104, 508–517. [Google Scholar] [CrossRef]
- Engels, S.; Sauer, K.P. Energy beyond the pupal stage: Larval nutrition and its long-time consequences for male mating performance in a scorpionfly. J. Insect Physiol. 2007, 53, 633–638. [Google Scholar] [CrossRef]
- Abraham, S.; Goane, L.; Cladera, J.; Vera, M.T. Effects of male nutrition on sperm storage and remating behavior in wild and laboratory Anastrepha fraterculus (Diptera: Tephritidae) females. J. Insect Physiol. 2011, 57, 1501–1509. [Google Scholar] [CrossRef]
- Abraham, S.; Cladera, J.; Goane, L.; Teresa Vera, M.T. Factors affecting Anastrepha fraterculus female receptivity modulation by accessory gland products. J. Insect Physiol. 2012, 58, 1–6. [Google Scholar] [CrossRef]
- Kaspi, R.; Mossinson, S.; Drezner, T.; Kamensky, B.; Yuval, B. Effects of larval diet on development rates and reproductive maturation of male and female Mediterranean fruit flies. Physiol. Entomol. 2002, 27, 29–38. [Google Scholar] [CrossRef]
- Alleyne, E.H. Studies on the biology and behavior of the West Indian sweet potato weevil, Euscepes postfasciatus (Fairmaire) (Coleoptera: Curculionidae). In Proceedings of the Caribbean Food Crops Society, Dover, Barbados, 22–28 August 1982; pp. 236–243. [Google Scholar] [CrossRef]
- Raman, K.V.; Alleyne, E.H. Biology and management of the West Indian sweet potato weevil, Euscepes postfasciatus. In Sweet Potato Pest Management: A Global Perspective; Jansson, R.K., Raman, K.V., Eds.; Westview Press: Boulder, CO, USA, 1991; pp. 263–281. [Google Scholar]
- Moriya, S. Is it possible to eradicate the two weevil pests of sweet potato [Ipomoea batatas], Cylas formicarius and Euscepes postfasciatus from Japan? Bull. Okinawa Agric. Exp. Stn. 1997, 18, 19–27. [Google Scholar]
- Yasuda, K.; Kohama, T. Distribution of the sweet-potato weevil, Cylas formicarius (Fabricius) and the West Indian sweet-potato weevil, Euscepes postfasciatus (Fairmire) in Okinawa prefecture. Proc. Assoc. Plant Prot. Kyushu 1990, 36, 123–125. [Google Scholar] [CrossRef] [Green Version]
- Kuba, H.; Kohama, T.; Haraguchi, D. Eradication projects of exotic sweet potato weevils using sit in Okinawa. In Proceedings of the NIAES-FFTC Joint International Seminar on Biological Invasions: Environmental Impacts and the Development of a Database for the Asian-Pacific Region, Tsukuba, Japan, 11–15 November 2003; Oka, M., Matsui, M., Shiomi, T., Ogawa, Y., Tsuchiya, K., Eds.; National Institute for Agro-Environmental Sciences and Food and Fertilizer Technology Center for the Asian and Pacific Region: Tsukuba, Japan, 2003; pp. 273–287. [Google Scholar]
- Knipling, E.F. Possibilities of insect control or eradication through the use of sexually sterile males. J. Econ. Entomol. 1955, 48, 459–462. [Google Scholar] [CrossRef]
- Dyck, V.A.; Hendrichs, J.; Robinson, A.S. Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2021. [Google Scholar]
- Ohishi, T.; Honma, A.; Himuro, C.; Teruya, K. Effect of sweet potato powder content in larval artificial diets on the yield and quality of mass-reared West Indian sweet potato weevil, Euscepes postfasciatus (Coleoptera: Curculionidae). Jpn. J. Appl. Entomol. Zool. 2018, 62, 123–126. [Google Scholar] [CrossRef] [Green Version]
- Shimoji, Y.; Yamagishi, M. Reducing rearing cost and increasing survival rate of West Indian sweetpotato weevil, Euscepes postfasciatus (Fairmaire) (Coleoptera: Curculionidae) on artificial larval diet. Appl. Entomol. Zool. 2004, 39, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Urasaki, K.; Ohno, S.; Haraguchi, D.; Kohama, T. Effect of simplifying the preparation of larval artificial diet on survival and development of the West Indian sweetpotato weevil Euscepes postfasciatus (Fairmaire) (Coleoptera: Curculionidae). Jpn. J. Appl. Entomol. Zool. 2009, 53, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Himuro, C.; Kumano, N.; Honma, A.; Ikegawa, Y.; Ohishi, T. Appropriate number of inoculated eggs for mass-rearing the West Indian sweet potato weevil, Euscepes postfasciatus (Coleoptera: Curculionidae). Appl. Entomol. Zool. 2018, 53, 157–164. [Google Scholar] [CrossRef]
- Misa, K.; Himuro, C.; Honma, A.; Ikegawa, Y.; Ohishi, T. Effects of storage periods of an artificial larval diet on the yield and quality of mass-reared West Indian sweet potato weevil (Coleoptera: Curculionidae). J. Econ. Entomol. 2020, 113, 2613–2618. [Google Scholar] [CrossRef]
- Kumano, N.; Kuriwada, T.; Shiromoto, K.; Haraguchi, D.; Kohama, T. Intensive resistance by females before copulation induces insemination failure in the West Indian sweet potato weevil Euscepes postfasciatus. Popul. Ecol. 2011, 53, 111–117. [Google Scholar] [CrossRef]
- Sato, Y.; Kohama, T. Post-copulatory mounting behavior of the West Indian sweetpotato weevil, Euscepes postfasciatus (Fairmaire) (Coleoptera: Curculionidae). Ethology 2007, 113, 183–189. [Google Scholar] [CrossRef]
- Himuro, C.; Ikegawa, Y.; Honma, A. Males use accessory gland substances to inhibit remating by females in West Indian sweet potato weevil (Coleoptera: Curculionidae). Ann. Entomol. Soc. Am. 2017, 110, 374–380. [Google Scholar] [CrossRef]
- Kumano, N.; Haraguchi, D.; Kohama, T. Female mating status does not affect male mating behavior in the West Indian sweetpotato weevil, Euscepes postfasciatus (Coleoptera: Curculionidae). Entomol. Exp. Appl. 2009, 131, 39–45. [Google Scholar] [CrossRef]
- Wigby, P.; Perry, J.C.; Kim, Y.-H.; Sirot, L.K. Developmental environment mediates male seminal protein investment in Drosophila melanogaster. Func. Ecol. 2016, 30, 410–419. [Google Scholar] [CrossRef] [PubMed]
- Okinawa Prefectural Plant Protection Center. Annual Report of Eradication Program for the Destructive Pest in Okinawa in 2005; Okinawa Prefectural Government, Department of Agriculture, Forestry and Fisheries: Okinawa, Japan, 2005. [Google Scholar]
- Ohishi, T.; Tsurui, K.; Teruya, K. Occurrence of Neogregarine-like protozoan Farinocystis sp. in an artificial diet mass-rearing system in the West Indian sweet potato weevil, Euscepes postfasciatus (Coleoptera: Curculionidae). Jpn. J. Appl. Entomol. Zool. 2016, 60, 131–137. [Google Scholar] [CrossRef] [Green Version]
- Okinawa Prefectural Plant Protection Center. Annual Report of Eradication Program for the Destructive Pest in Okinawa in 2018; Okinawa Prefectural Government, Department of Agriculture, Forestry and Fisheries: Okinawa, Japan, 2018. [Google Scholar]
- Kohama, T.; Sugiyama, M. External characteristics for sexing the West Indian sweet potato weevil, Euscepes postfasciatus (Fairmaire) (Coleoptera: Curculionidae). Entomol. Sci. 2000, 3, 43–46. [Google Scholar]
- Shimizu, T.; Moriya, S. Daily locomotor activity in the Indian sweet potato weevil, Euscepes postfasciatus (Fairmaire) (Coleoptera: Curculionidae) and sweet potato weevil, Cylas formicarius (Fabricius) (Coleoptera: Brentidae) monitored by an actograph system. Appl. Entomol. Zool. 1996, 31, 626–628. [Google Scholar] [CrossRef] [Green Version]
- Kumano, N.; Haraguchi, D.; Kohama, T. Effect of irradiation on mating performance and mating ability in the West Indian sweetpotato weevil, Euscepes postfasciatus. Entomol. Exp. Appl. 2008, 127, 229–236. [Google Scholar] [CrossRef]
- Carroll, K.J. On the use and utility of the Weibull model in the analysis of survival data. Control. Clin. Trials 2003, 24, 682–701. [Google Scholar] [CrossRef]
- Zhu, H.P.; Xia, X.; Yu, C.H.; Adnan, A.; Liu, S.F.; Du, Y.K. Application of Weibull Model for survival of patients with gastric cancer. BMC Gastroenterol. 2011, 11, 1. [Google Scholar] [CrossRef] [Green Version]
- SAS Institute Inc. JMP® 14 Statistics and Graphics Guide; SAS Institute Inc.: Cary, NC, USA, 2018. [Google Scholar]
- Klowden, M.J. Sexual receptivity in Anopheles gambiae mosquitoes: Absence of control by male accessory gland substances. J. Insect Physiol. 2001, 47, 661–666. [Google Scholar] [CrossRef]
- Nakagawa, S.; Farias, G.J.; Suda, D.; Cunningham, R.T.; Chambers, D.L. Reproduction of the Mediterranean fruit fly:1 frequency of mating in the laboratory. Ann. Entomol. Soc. Am. 1971, 64, 949–950. [Google Scholar] [CrossRef]
- Gromko, M.H.; Newport, M.E.A.; Kortier, M.G. Sperm dependence of female receptivity to remating in Drosophila melanogaster. Evolution 1984, 38, 1273–1282. [Google Scholar] [CrossRef] [PubMed]
- Gromko, M.H.; Markow, T.A. Courtship and remating in field populations of Drosophila. Anim. Behav. 1993, 45, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Sakurai, T. Receptivity of female remating and sperm number in the sperm storage organ in the bean bug, Riptortus clavatus (Heteroptera: Alydidae). Res. Popul. Ecol. 1998, 40, 167–172. [Google Scholar] [CrossRef]
- Whittier, T.S.; Shelly, T.E. Productivity of singly vs. multiply mated female Mediterranean fruit fly, Ceratitis capitata (Diptera: Tephritidae). J. Kans. Entomol. Soc. 1993, 66, 200–209. [Google Scholar]
- Sirot, L.K.; Wolfner, M.F.; Wigby, S. Protein-specific manipulation of ejaculate composition in response to female mating status in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2011, 108, 9922–9926. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.S. The functional morphology and biochemistry of insect male accessory glands and their secretions. Annu. Rev. Entomol. 1984, 29, 233–255. [Google Scholar] [CrossRef]
- Gillott, C. Male accessory gland secretions: Modulators of female reproductive physiology and behavior. Annu. Rev. Entomol. 2003, 48, 163–184. [Google Scholar] [CrossRef]
- Chapman, T.; Bangham, J.; Vinti, G.; Seifried, B.; Lung, O.; Wolfner, M.F.; Smith, H.K.; Partridge, L.; Partridge, L. The sex peptide of Drosophila melanogaster: Female post-mating responses analyzed by using RNA interference. Proc. Natl. Acad. Sci. USA 2003, 100, 9923–9928. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, N.; Takami, Y. Inhibition of female mating by male accessory gland substances in the ground beetle Leptocarabus procerulus. Physiol. Entomol. 2014, 39, 12–18. [Google Scholar] [CrossRef]
- Himuro, C.; Fujisaki, K. Males of the seed bug Togo hemipterus (Heteroptera: Lygaeidae) use accessory gland substances to inhibit remating by females. J. Insect Physiol. 2008, 54, 1538–1542. [Google Scholar] [CrossRef] [PubMed]
- Miyatake, T.; Chapman, T.; Partridge, L. Mating-induced inhibition of remating in female Mediterranean fruit flies Ceratitis capitata. J. Insect Physiol. 1999, 45, 1021–1028. [Google Scholar] [CrossRef]
- Takami, Y.; Sasabe, M.; Nagata, N.; Sota, T. Dual function of seminal substances for mate guarding in a ground beetle. Behav. Ecol. 2008, 19, 1173–1178. [Google Scholar] [CrossRef] [Green Version]
- Wedell, N. Female receptivity in butterflies and moths. J. Exp. Biol. 2005, 208, 3433–3440. [Google Scholar] [CrossRef] [Green Version]
- Yamane, T.; Kimura, Y.; Katsuhara, M.; Miyatake, T. Female mating receptivity inhibited by injection of male-derived extracts in Callosobruchus chinensis. J. Insect Physiol. 2008, 54, 501–507. [Google Scholar] [CrossRef]
- Ram, K.R.; Wolfner, M.F. Sustained post-mating response in Drosophila melanogaster requires multiple seminal fluid proteins. PLoS Genet. 2007, 3, e238. [Google Scholar] [CrossRef] [Green Version]
- Goenaga, J.; Yamane, T.; Rönn, J.; Arnqvist, G. Within-species divergence in the seminal fluid proteome and its effect on male and female reproduction in a beetle. BMC Evol. Biol. 2015, 15, 266. [Google Scholar] [CrossRef] [Green Version]
- Himuro, C.; Honma, A.; Ikegawa, Y.; Ohishi, T.; Miyatake, T. Male body size does not affect the refractory period of females in the West Indian sweet potato weevil Euscepes postfasciatus (Fairmaire) (Coleoptera: Curculionidae) and the seed bug Togohemipterus (Scott) (Heteroptera: Lygaeidae). J. Ethol. 2021, 39, 39–46. [Google Scholar] [CrossRef]
- Sirot, L.K.; Findlay, G.D.; Sitnik, J.L.; Frasheri, D.; Avila, F.W.; Wolfner, M.F. Molecular characterization and evolution of a gene family encoding both female- and male-specific reproductive proteins in Drosophila. Mol. Biol. Evol. 2014, 31, 1554–1567. [Google Scholar] [CrossRef] [Green Version]
- Sirot, L.K. Modulation of seminal fluid molecules by males and females. Curr. Opin. Insect Sci. 2019, 35, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Teruya, K.; Kumano, N. Effect of long-term mass-rearing on mating performance in the West Indian sweet potato weevil, Euscepes postfasciatus. Jpn. J. Appl. Entomol. Zool. 2015, 59, 17–22. [Google Scholar] [CrossRef] [Green Version]
- Abraham, S.; Liendo, M.C.; Devescovi, F.; Peralta, P.A.; Yusef, V.; Ruiz, J.; Cladera, J.L.; Vera, M.T.; Segura, D.F. Remating behavior in Anastrepha fraterculus (Diptera: Tephritidae) females is affected by male juvenile hormone analog treatment but not by male sterilization. Bull. Entomol. Res. 2013, 103, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Haq, I.U.; Vreysen, M.J.B.; Teal, P.E.A.; Hendrichs, J. Methoprene application and diet protein supplementation to male melon fly, Bactrocera cucurbitae, modifies female remating behavior. Insect Sci. 2014, 21, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Olsson, M.; Madsen, T.; Shine, R. Is sperm really so cheap? Costs of reproduction in male adders, Vipera berus. Proc. R. Soc. Lond. B 1997, 264, 455–459. [Google Scholar] [CrossRef] [Green Version]
- Lüpold, S.; Tomkins, J.L.; Simmons, L.W.; Fitzpatrick, J.L. Female monopolization mediates the relationship between pre- and postcopulatory sexual traits. Nat. Commun. 2014, 5, 3184. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, F.; Rebar, D.; Greenfield, M.D. When do trade-offs occur? The roles of energy constraints and trait flexibility in bushcricket populations. J. Evol. Biol. 2018, 31, 287–301. [Google Scholar] [CrossRef]
- Somjee, U.; Miller, C.W.; Tatarnic, N.J.; Simmons, L.W. Experimental manipulation reveals a trade-off between weapons and testes. J. Evol. Biol. 2018, 31, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Tuni, C.; Han, C.S.; Dingemanse, N.J. Multiple biological mechanisms result in correlations between pre- and post-mating traits that differ among versus within individuals and genotypes. Proc. Biol. Sci. 2018, 285, 20180951. [Google Scholar] [CrossRef] [Green Version]
- Chung, M.J.; Jennions, M.D.; Fox, R.J. Quantifying the costs of pre- and postcopulatory traits for males: Evidence that costs of ejaculation are minor relative to mating effort. Evol. Lett. 2021, 5, 315–327. [Google Scholar] [CrossRef]
- Clark, N.L.; Aagaard, J.E.; Swanson, W.J. Evolution of reproductive proteins from animals and plants. Reproduction 2006, 131, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, J.L.; Ripoll, D.R.; Aquadro, C.F.; Wolfner, M.F. Comparative structural modeling and inference of conserved protein classes in Drosophila seminal fluid. Proc. Natl. Acad. Sci. USA 2004, 101, 13542–13547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicens, A.; Borziak, K.; Karr, T.L.; Roldan, E.R.S.; Dorus, S. Comparative sperm proteomics in mouse species with divergent mating systems. Mol. Biol. Evol. 2017, 34, 1403–1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilburn, D.B.; Swanson, W.J. From molecules to mating: Rapid evolution and biochemical studies of reproductive proteins. J. Proteom. 2016, 135, 12–25. [Google Scholar] [CrossRef] [Green Version]
- Sherman, M.; Tamashiro, M. The Sweetpotato Weevils in Hawaii: Their Biology and Control; Hawaii Agricultural Experiment Station, University of Hawaii: Honolulu, HI, USA, 1954; pp. 1–36. [Google Scholar]
- Ohno, S.; Haraguchi, D.; Urasaki, K.; Kohama, T. The West Indian sweet-potato weevil, Euscepes postfasciatus: Its occurrence and current problems of its control in Kume Island, Okinawa. Kontyu Shizen 2006, 41, 25–30. (In Japanese) [Google Scholar]
- Sato, Y.; Miyai, S.; Haraguchi, D.; Ohno, S.; Kohama, T.; Kawamura, K.; Yamagishi, M. Population dynamics of the West Indian sweetpotato weevil Euscepes postfasciatus (Fairmaire): A simulation analysis. J. Appl. Entomol. 2010, 134, 303–312. [Google Scholar] [CrossRef]
- Itô, Y.; Yamamura, K.; Manoukis, N.C. Role of population and behavioural ecology in the sterile insect technique. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 245–282. [Google Scholar]
- Pereira, R.; Yuval, B.; Liedo, P.; Teal, P.E.A.; Shelly, T.E.; McInnis, D.O.; Haq, I.; Taylor, P.W.; Hendrichs, J. Improving post-factory performance of sterile male fruit flies in support of the sterile insect technique. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 631–657. [Google Scholar]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Himuro, C.; Misa, K.; Honma, A.; Ikegawa, Y.; Ohishi, T.; Kumano, N. Effects of Larval Diet on the Male Reproductive Traits in the West Indian Sweet Potato Weevils Euscepes postfasciatus (Coleoptera: Curculionidae). Insects 2022, 13, 389. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13040389
Himuro C, Misa K, Honma A, Ikegawa Y, Ohishi T, Kumano N. Effects of Larval Diet on the Male Reproductive Traits in the West Indian Sweet Potato Weevils Euscepes postfasciatus (Coleoptera: Curculionidae). Insects. 2022; 13(4):389. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13040389
Chicago/Turabian StyleHimuro, Chihiro, Kinjo Misa, Atsushi Honma, Yusuke Ikegawa, Tsuyoshi Ohishi, and Norikuni Kumano. 2022. "Effects of Larval Diet on the Male Reproductive Traits in the West Indian Sweet Potato Weevils Euscepes postfasciatus (Coleoptera: Curculionidae)" Insects 13, no. 4: 389. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13040389