
No Evidence of Bacterial Symbionts Influencing Host Specificity in Aphis gossypii Glover (Hemiptera: Aphididae)


Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Aphid and Plant Materials
2.2. Host Specificity Comparison between Winged and Wingless Morph
2.3. Bacterial Symbionts Analysis of Host-Specialized Biotypes
2.4. Buchnera Quantification of Winged and Wingless Aphids
2.5. Evaluating the Effects of Buchnera Abundance on Host Specificity
2.6. Data Analysis
3. Results
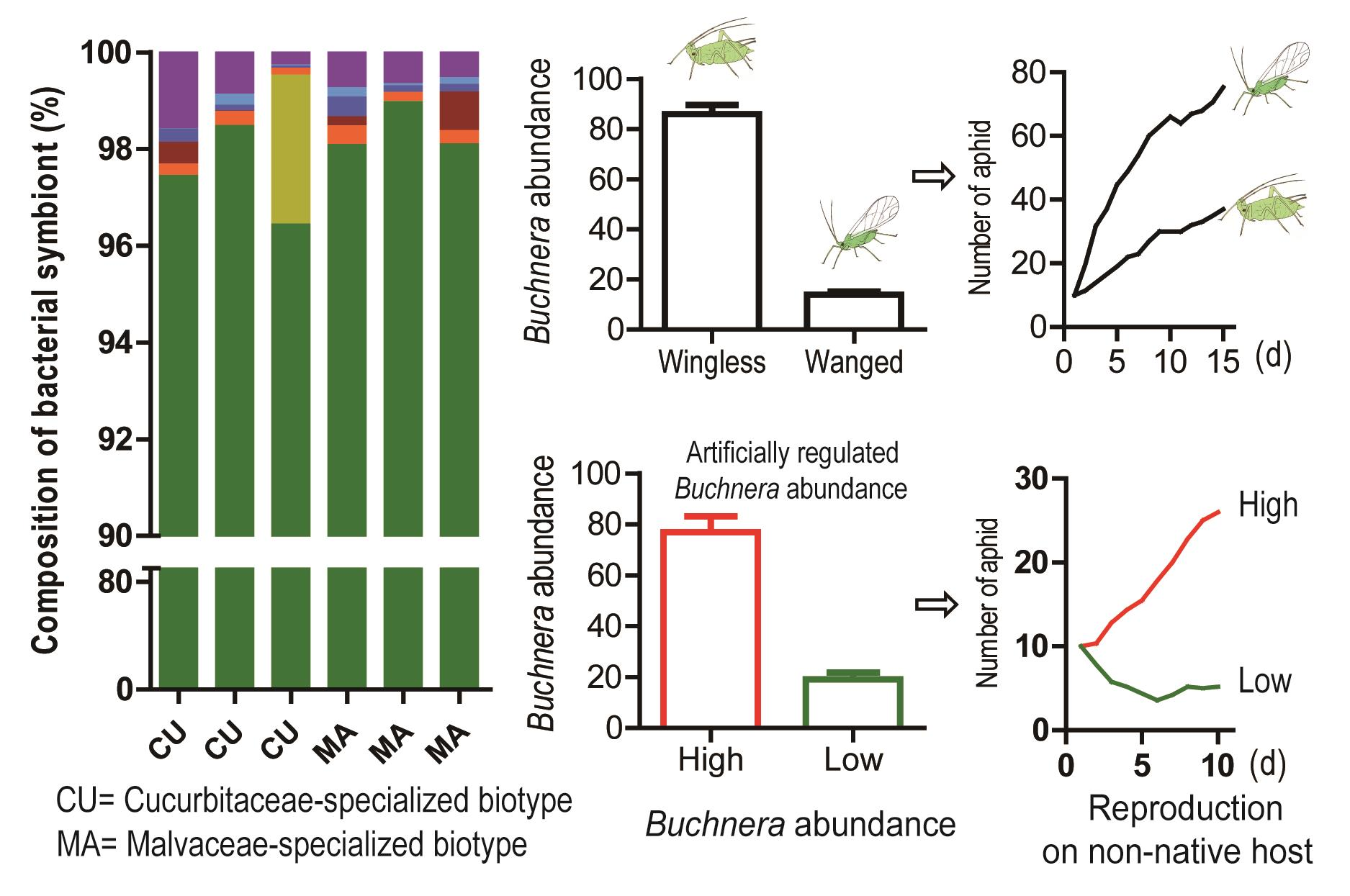
3.1. Host Specificity of Winged and Wingless Host-Specialized Biotypes
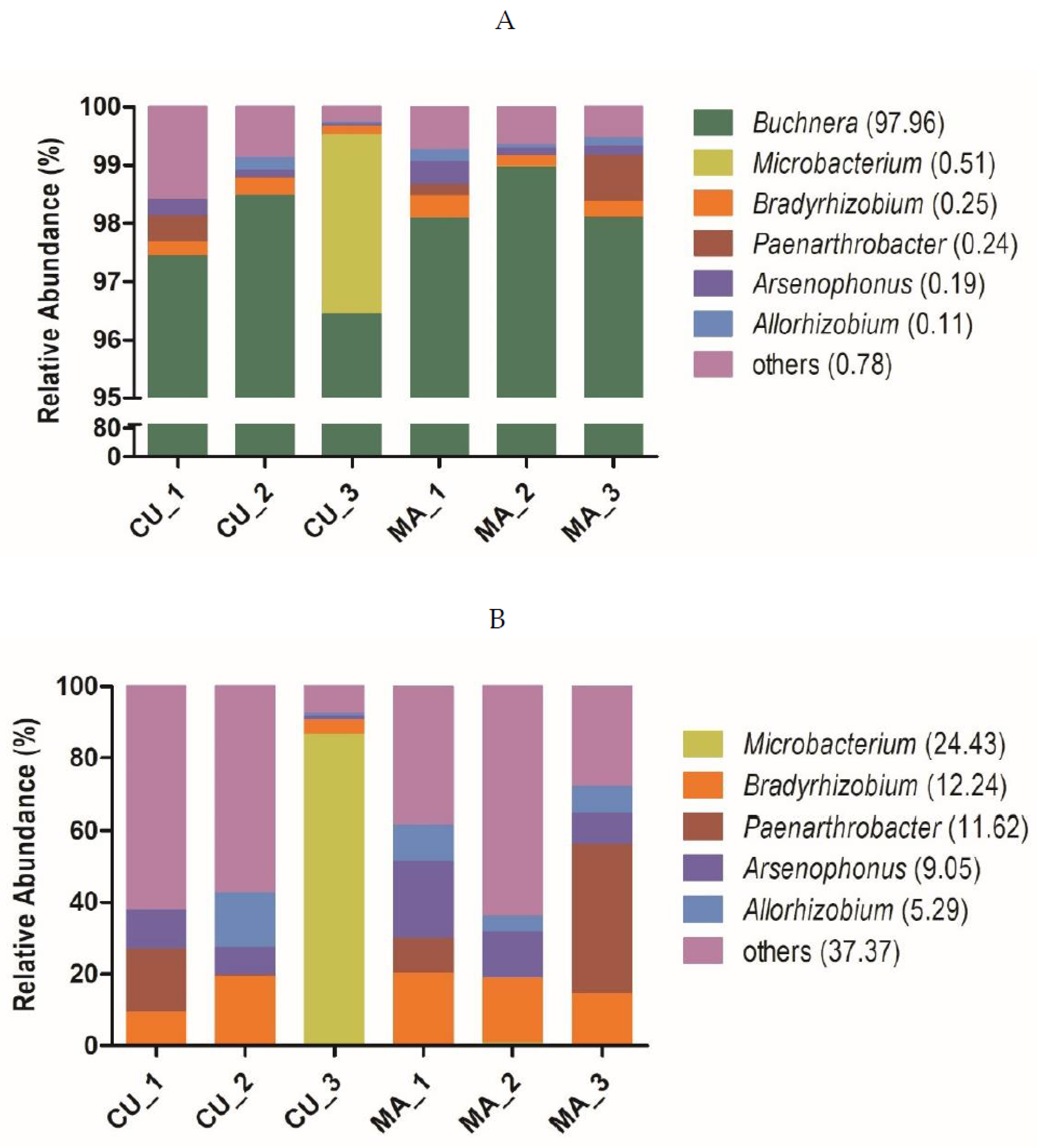
3.2. Symbiont Composition in CU and MA Biotype
3.3. Buchnera Abundance on Winged and Wingless Morphs
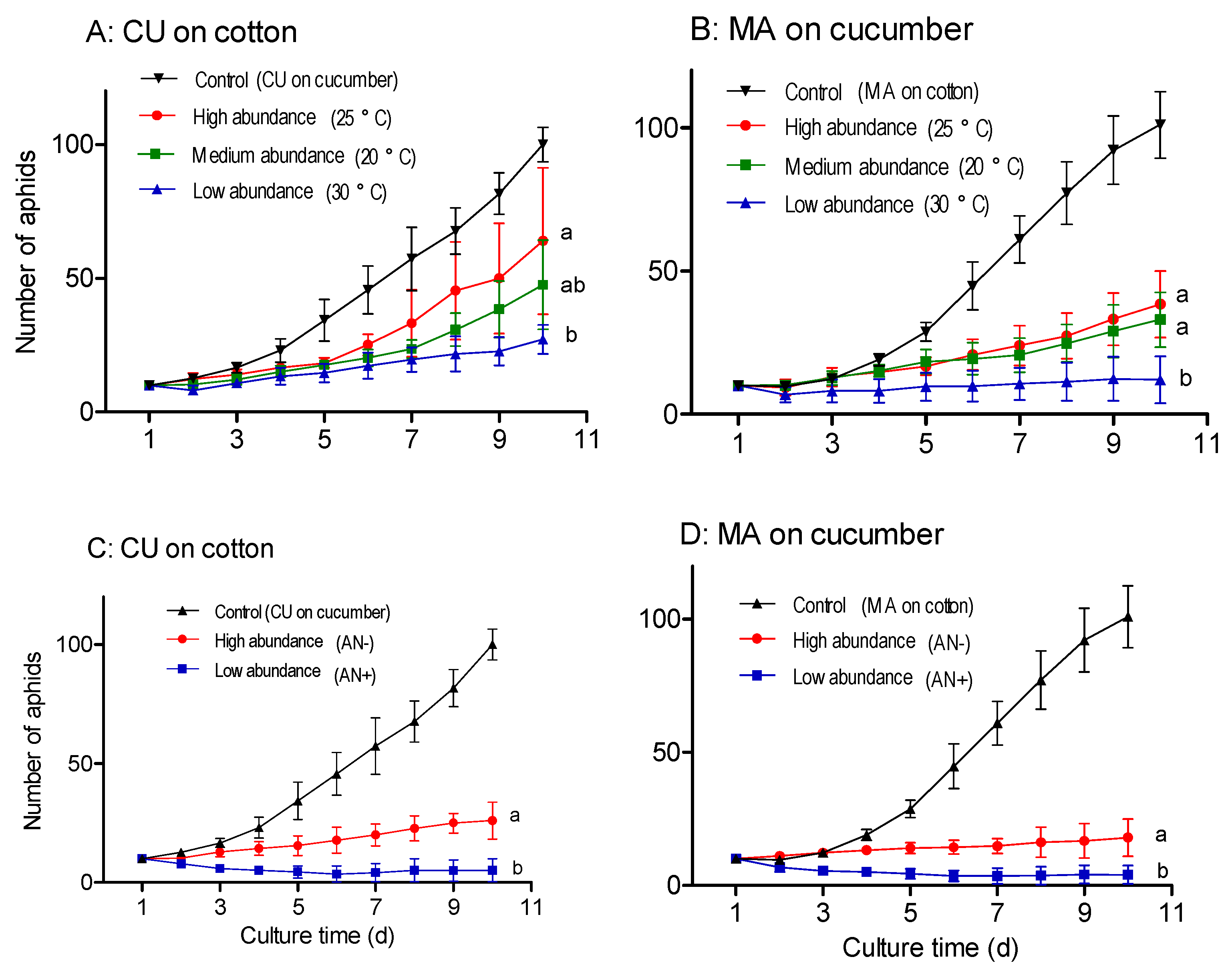
3.4. Response of Buchnera Abundance to Temperature and Antibiotics
3.5. Reproduction of Aphids with Different Abundance of Buchnera on Non-Native Hosts
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blackman, R.L.; Eastop, V.F. Aphids on The World’s Crops: An Identification and Information Guide; John Wiley and Sons, Ltd.: Chichester, UK, 2000; 466p. [Google Scholar]
- Moritz, D.R.; Da Silva Santos, N.; Zanini, F.; Pacini, B.A.; Petry, H.B.; Bertolini, E. Effect of vegetable oil on the efficiency of transmission of cowpea aphid-borne mosaic virus by Aphis gossypii Glover in passion fruit plants. Trop. Plant Pathol. 2021, 47, 298–302. [Google Scholar] [CrossRef]
- Müller, C.B.; Williams, I.S.; Hardie, J. The role of nutrition, crowding and interspecific interactions in the development of winged aphids. Ecol. Entomol. 2001, 26, 330–340. [Google Scholar] [CrossRef]
- Peccoud, J.; Simon, J.C.; Von Dohlen, C.; Coeur d’acier, A.; Plantegenest, M.; Vanlerberghe-Masutti, F.; Jousselin, E. Evolutionary history of aphid-plant associations and their role in aphid diversification. Comptes Rendus Biol. 2010, 333, 474–487. [Google Scholar] [CrossRef] [PubMed]
- Guldemond, J.A.; Tigges, W.T.; De Vrijer, P.W.F. Host Races of Aphis gossypii (Homoptera: Aphididae) on Cucumber and Chrysanthemum. Environ. Entomol. 1994, 23, 1235–1240. [Google Scholar] [CrossRef]
- Carletto, J.; Lombaert, E.; Chavigny, P.; Brévault, T.; Lapchin, L.; Vanlerberghe-Masutti, F. Ecological specialization of the aphid Aphis gossypii Glover on cultivated host plants. Mol. Ecol. 2009, 18, 2198–2212. [Google Scholar] [CrossRef]
- Najar-Rodríguez, A.J.; McGraw, E.A.; Hull, C.D.; Mensah, R.K.; Walter, G.H. The ecological differentiation of asexual lineages of cotton aphids: Alate behaviour, sensory physiology, and differential host associations. Biol. J. Linn. Soc. 2009, 97, 503–519. [Google Scholar] [CrossRef] [Green Version]
- Meng, L.; Li, B.P. Researches on biotypes of cotton aphid in xinjiang. Acta Geogr. Sin. 2001, 13, 30–35. [Google Scholar]
- Liu, X.D.; Zhai, B.P.; Zhang, X.X.; Lu, Y. Differentiation in morphometrics and ecological adaptability of cotton and cucumber biotypes of the cotton aphid, Aphis gossypii (Homoptera: Aphididae). Acta Entomol. Sin. 2004, 47, 768–773. [Google Scholar]
- Liu, X.D.; Zhai, B.P.; Zhang, X.X. Studies on the host biotypes and its cause of cotton aphid in Nanjing, China. China Sci. Agric. Sin. 2002, 1, 1211–1215. [Google Scholar]
- Wang, L.; Zhang, S.; Luo, J.-Y.; Wang, C.-Y.; Lv, L.-M.; Zhu, X.-Z.; Li, C.-H.; Cui, J.-J. Identification of Aphis gossypii Glover (Hemiptera: Aphididae) Biotypes from Different Host Plants in North China. PLoS ONE 2016, 11, e0146345. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Liang, X.L.; Zhao, H.Y.; Xu, T.T.; Liu, X.D. Special Plant Species Determines Diet Breadth of Phytophagous Insects: A Study on Host Plant Expansion of the Host-Specialized Aphis gossypii Glover. PLoS ONE 2013, 8, e60832. [Google Scholar] [CrossRef] [Green Version]
- Weinert, L.A.; Araujo-Jnr, E.V.; Ahmed, M.Z.; Welch, J.J. The incidence of bacterial endosymbionts in terrestrial arthropods. Proc. R. Soc. B Boil. Sci. 2015, 282, 20150249. [Google Scholar] [CrossRef]
- Moran, N.A.; Baumann, P. Bacterial endosymbionts in animals. Curr. Opin. Microbiol. 2000, 3, 270–275. [Google Scholar] [CrossRef]
- Tsuchida, T.; Koga, R.; Fukatsu, T. Host Plant Specialization Governed by Facultative Symbiont. Science 2004, 303, 1989. [Google Scholar] [CrossRef] [Green Version]
- McLean, A.H.C.; Van Asch, M.; Ferrari, J.; Godfray, H.C.J. Effects of bacterial secondary symbionts on host plant use in pea aphids. Proc. R. Soc. B Boil. Sci. 2011, 278, 760–766. [Google Scholar] [CrossRef]
- Hansen, A.K.; Moran, N.A. The impact of microbial symbionts on host plant utilization by herbivorous insects. Mol. Ecol. 2014, 23, 1473–1496. [Google Scholar] [CrossRef] [PubMed]
- Leonardo, T.E.; Muiru, G.T. Facultative symbionts are associated with host plant specialization in pea aphid populations. Proc. R. Soc. B Boil. Sci. 2003, 270 (Suppl. 2), S209–S212. [Google Scholar] [CrossRef] [Green Version]
- Brady, C.M.; White, J.A. Cowpea aphid (Aphis craccivora) associated with different host plants has different facultative endosymbionts: Aphid symbiont and host plant associations. Ecol. Entomol. 2013, 38, 433–437. [Google Scholar] [CrossRef]
- Moran, N.A.; McCutcheon, J.P.; Nakabachi, A. Genomics and Evolution of Heritable Bacterial Symbionts. Annu. Rev. Genet. 2008, 42, 165–190. [Google Scholar] [CrossRef] [Green Version]
- Montllor, C.B.; Maxmen, A.; Purcell, A.H. Facultative bacterial endosymbionts benefit pea aphids Acyrthosiphon pisum under heat stress: Aphid symbionts and heat stress. Ecol. Entomol. 2002, 27, 189–195. [Google Scholar] [CrossRef]
- Dunbar, H.E.; Wilson, A.C.C.; Ferguson, N.R.; Moran, N.A. Aphid Thermal Tolerance Is Governed by a Point Mutation in Bacterial Symbionts. PLoS Biol. 2007, 5, e96. [Google Scholar] [CrossRef] [PubMed]
- Frago, E.; Mala, M.; Weldegergis, B.T.; Yang, C.; McLean, A.; Godfray, H.C.J.; Gols, R.; Dicke, M. Symbionts protect aphids from parasitic wasps by attenuating herbivore-induced plant volatiles. Nat. Commun. 2017, 8, 1860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Łukasik, P.; van Asch, M.; Guo, H.; Ferrari, J.; Godfray, H.C.J. Unrelated facultative endosymbionts protect aphids against a fungal pathogen. Ecol. Lett. 2013, 16, 214–218. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Jiang, L.; Qiao, G.; Chen, J. The Bacterial Flora Associated with the Polyphagous Aphid Aphis gossypii Glover (Hemiptera: Aphididae) Is Strongly Affected by Host Plants. Microb. Ecol. 2020, 79, 971–984. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.-J.; He, H.-P.; Zhao, H.-M.; Xian, Y.-D.; Guo, H.; Liu, B.; Xue, K. Microbiome diversity of cotton aphids (Aphis gossypii) is associated with host alternation. Sci. Rep. 2021, 11, 5260. [Google Scholar] [CrossRef]
- Najar-Rodríguez, A.J.; McGraw, E.A.; Mensah, R.K.; Pittman, G.W.; Walter, G.H. The microbial flora of Aphis gossypii: Patterns across host plants and geographical space. J. Invertebr. Pathol. 2009, 100, 123–126. [Google Scholar] [CrossRef]
- Ali, F.; Hu, X.; Wang, D.; Yang, F.; Guo, H.; Wang, Y. Plant pathogen-mediated rapid acclimation of a host-specialized aphid to a non-host plant. Ecol. Evol. 2021, 11, 15261–15272. [Google Scholar] [CrossRef]
- Cloutier, C.; Douglas, A.E. Impact of a parasitoid on the bacterial symbiosis of its aphid host. Entomol. Exp. Appl. 2003, 109, 13–19. [Google Scholar] [CrossRef]
- Mech, A.M.; Harper, S.J.; Havill, N.P.; Von Dohlen, C.D.; Burke, G.R. Ecological factors influencing the beneficial endosymbionts of the hemlock woolly adelgid (Hemiptera: Adelgidae): Factors influencing A. tsugae endosymbionts. Insect Sci. 2019, 26, 97–107. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-Y.; Lai, C.-Y.; Kuo, M.-H. Temperature effect on the growth of Buchnera endosymbiont in Aphis craccivora (Hemiptera: Aphididae). Symbiosis 2009, 49, 53–59. [Google Scholar] [CrossRef]
- Wilkinson, T. The elimination of intracellular microorganisms from insects: An analysis of antibiotic-treatment in the pea aphid (Acyrthosiphon pisum). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 1998, 119, 871–881. [Google Scholar] [CrossRef]
- Lu, W.-N.; Chiu, M.-C.; Kuo, M.-H. Host life stage- and temperature-dependent density of the symbiont Buchnera aphidicola in a subtropical pea aphid (Acyrthosiphon pisum) population. J. Asia-Pac. Entomol. 2014, 17, 537–541. [Google Scholar] [CrossRef]
- Aldyhim, Y.N.; Khalil, A.F. Influence of temperature and daylength on population development of Aphis gossypii on Cucurbita pepo. Entomol. Exp. Appl. 1993, 67, 167–172. [Google Scholar] [CrossRef]
- Kersting, U.; Satar, S.; Uygun, N. Effect of temperature on development rate and fecundity of apterous Aphis gossypii Glover (Hom., Aphididae) reared on Gossypium hirsutum L. J. Appl. Entomol. 1999, 123, 23–27. [Google Scholar] [CrossRef]
- Kocourek, F.; Havelka, J.; Beránková, J.; Jarošík, V. Effect of temperature on development rate and intrinsic rate of increase of Aphis gossypii reared on greenhouse cucumbers. Entomol. Exp. Appl. 1994, 71, 59–64. [Google Scholar] [CrossRef]
- Wu, K.M.; Liu, Q.X. Study on the influence of temperature on life parameters of cotton aphid. Cotton Sci. 1992, 4, 61–68. [Google Scholar]
- Zhao, J.Y.; Zhang, X.X.; Zhang, G.X.; Chen, X.F. The ecological conditions related to seasonal population variation of cotton aphid, Aphis gossypii Glover. Acta Ecol. Sin. 2001, 21, 2045–2050. [Google Scholar]
- Zamani, A.A.; Talebi, A.A.; Fathipour, Y.; Baniameri, V. Effect of temperature on biology and population growth parameters of Aphis gossypii Glover (Hom., Aphididae) on greenhouse cucumber. J. Appl. Entomol. 2006, 130, 453–460. [Google Scholar] [CrossRef]
- Barman, A.K.; Gadhave, K.R.; Dutta, B.; Srinivasan, R. Plasticity in host utilization by two host-associated populations of Aphis gossypii Glover. Bull. Entomol. Res. 2018, 108, 360–369. [Google Scholar] [CrossRef]
- Nishikori, K.; Morioka, K.; Kubo, T.; Morioka, M. Age- and morph-dependent activation of the lysosomal system and Buchnera degradation in aphid endosymbiosis. J. Insect Physiol. 2009, 55, 351–357. [Google Scholar] [CrossRef]
- Li, X.H.; Li, B.P. Effect of temperature on the abundance of mycetocytes in Aphis fabae Scopoli (Aphididae, Homoptera) and body size of their hosts. Acta Entomol. Sin. 2006, 3, 428–432. [Google Scholar]
- Su, Q.; Pan, H.P.; Wang, S.L.; Wu, Q.J.; Xu, B.Y.; Zhang, Y.J. A comparative study of the removal of endosymbionts in Bemisia tabaci biotypes B and Q using three antibiotics. Chin. J. Appl. Entomol. 2012, 49, 190–196. [Google Scholar]
- Zhang, X.C. Sequencing and phylogenetic analysis of the wsp gene of Wolbachia in Echinothrips americanus Morgan. J. Northwest A&F Univ. Nat. Sci. Ed. 2018, 40, 131–139. [Google Scholar]
- Liu, F.-H.; Kang, Z.-W.; Tan, X.-L.; Fan, Y.-L.; Tian, H.-G.; Liu, T.-X. Physiology and defense responses of wheat to the infestation of different cereal aphids. J. Integr. Agric. 2020, 19, 1464–1474. [Google Scholar] [CrossRef]
- Wang, D.; Shi, X.; Dai, P.; Liu, D.; Dai, X.; Shang, Z.; Ge, Z.; Meng, X. Comparison of fitness traits and their plasticity on multiple plants for Sitobion avenae infected and cured of a secondary endosymbiont. Sci. Rep. 2016, 6, 23177. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Level | CU | MA |
|---|---|---|---|
| Temperature | |||
| 15 °C | 49.00 ± 3.29 b | 35.22 ± 2.24 c | |
| 20 °C | 61.22 ± 6.12 b | 55.70 ± 4.98 b | |
| 25 °C | 108.10 ± 5.02 a | 79.00 ± 2.61 a | |
| 30 °C | 25.70 ± 2.69 c | 22.70 ± 5.51 c | |
| 35 °C | 10.50 ± 2.58 d | 5.20 ± 1.25 d | |
| Antibiotics | |||
| −(0 μg/mL) | 102.08 ± 2.62 a | 74.00 ± 2.52 a | |
| +(200 μg/mL) | 17.58 ± 3.65 b | 14.24 ± 2.51 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, H.; Yang, F.; Meng, M.; Feng, J.; Yang, Q.; Wang, Y. No Evidence of Bacterial Symbionts Influencing Host Specificity in Aphis gossypii Glover (Hemiptera: Aphididae). Insects 2022, 13, 462. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050462
Guo H, Yang F, Meng M, Feng J, Yang Q, Wang Y. No Evidence of Bacterial Symbionts Influencing Host Specificity in Aphis gossypii Glover (Hemiptera: Aphididae). Insects. 2022; 13(5):462. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050462
Chicago/Turabian StyleGuo, Hao, Fengying Yang, Min Meng, Jingjing Feng, Qinglan Yang, and Yongmo Wang. 2022. "No Evidence of Bacterial Symbionts Influencing Host Specificity in Aphis gossypii Glover (Hemiptera: Aphididae)" Insects 13, no. 5: 462. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050462