
Optimization of a Mass Trapping Method against the Striped Cucumber Beetle Acalymma vittatum in Organic Cucurbit Fields

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites and Experimental Plots
2.2. Pest Control Potential of the Mass Trapping Method
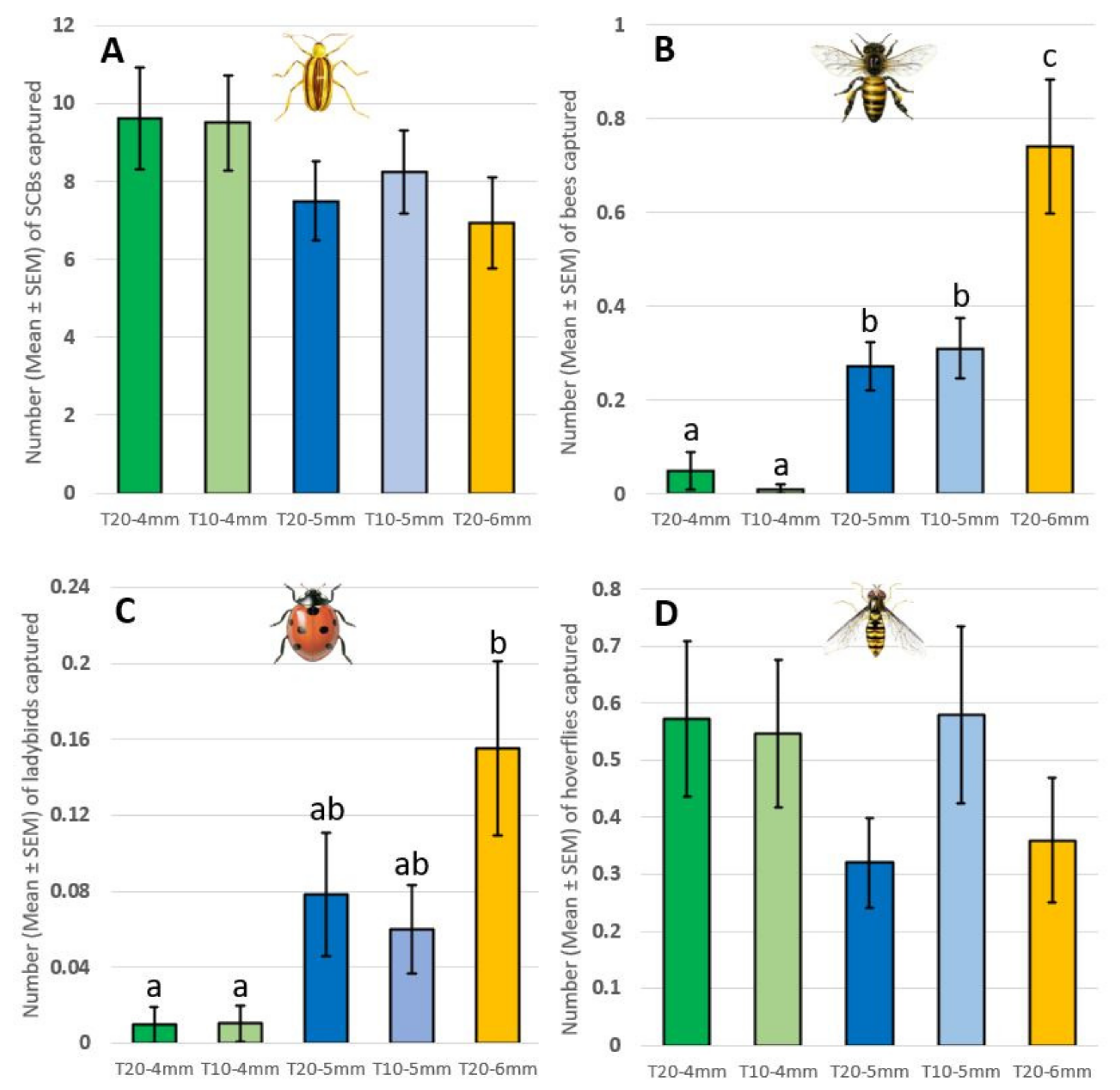
2.3. Selection of the Optimal Trap (Number and Diameter of Trap Entrances)
- T20-4mm: traps with two series of ten holes of 4 mm in Ø (diameter) per side;
- T10-4mm: traps with two series of five holes of 4 mm in Ø per side;
- T20-5mm: traps with two series of ten holes of 5 mm in Ø per side;
- T10-5mm: traps with two series of five holes of 5 mm in Ø per side;
- T20-6mm: traps with two series of ten holes of 6 mm in Ø per side.
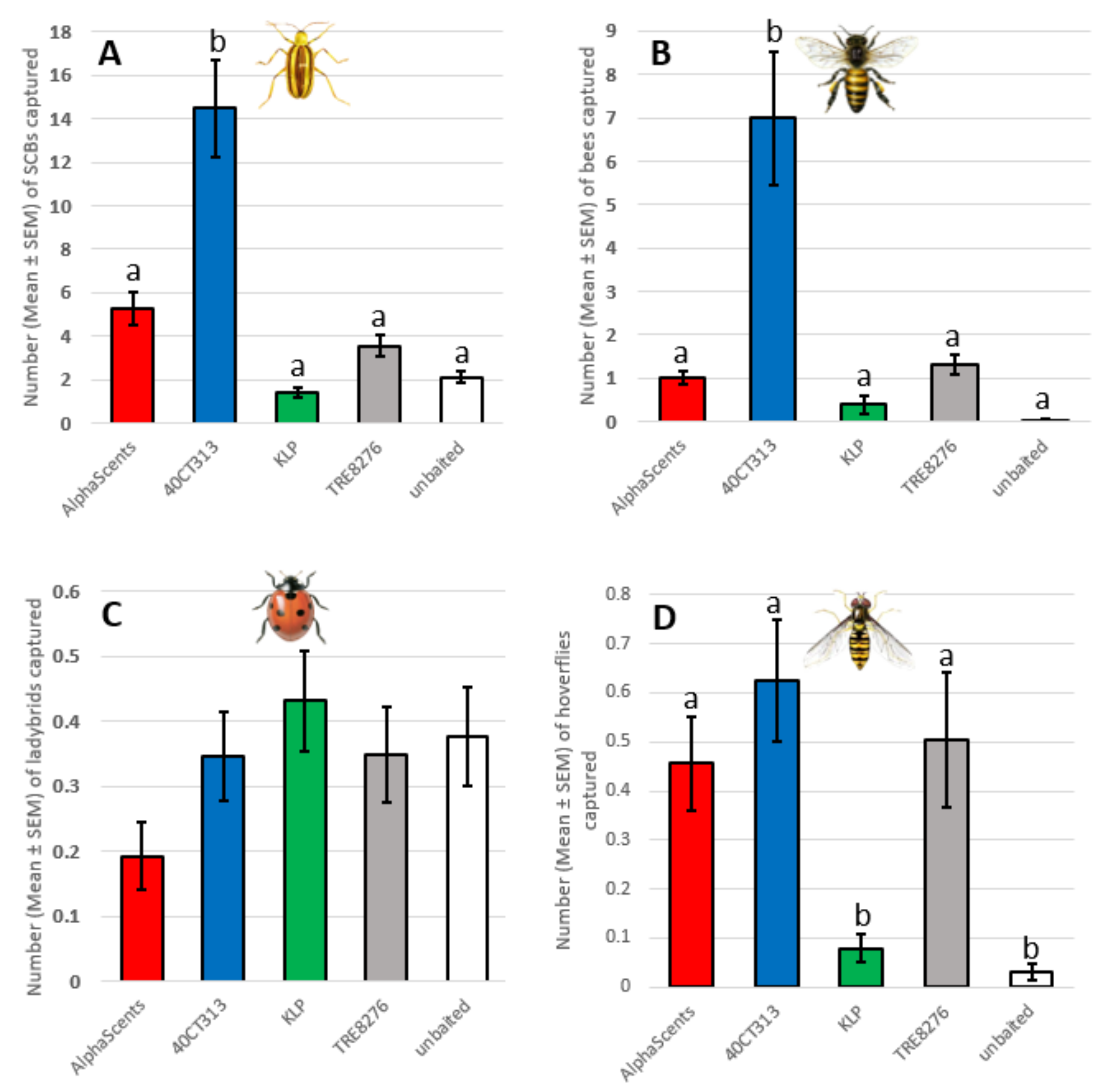
2.4. Selection of the Optimal Lure
- Alpha Scents SCB lure (Alpha Scents Inc., Canby, OR, USA) composed of Indole, (E)-cinnamaldehyde and 1,2,4-trimethoxybenzene, i.e., TIC mixture [26];
- 40CT313 5-compound lure (Distribution Solida Inc., Saint-Ferréol-les-Neiges, QC, Canada), the identities of the five compounds of the 40CT313 lure were not disclosed. This attractant is the equivalent of the AgBio 5-compound lure P313-B5 (AgBio Inc., Westminster, CO, USA), which Piñero had found to be most attractive to SCB in his research;
- KLP lure composed of 4-methoxy-cinnamaldehyde and indole (Csalomon®, Budapest, Hungary);
- Trécé TRE8276 (1,2,4-trimethoxybenzene [500 mg], indole [500 mg], and (E)-cinnamaldehyde [500 mg], i.e., TIC mixture (Trécé®, Adair, OK, USA);
- No lure.
3. Results
3.1. Pest Control Potential of the Mass Trapping Method
3.2. Selection of the Optimal Trap (Number and Ø of Trap Entrances)
3.3. Selection of the Optimal Lure
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haber, A.I.; Wallingford, A.K.; Grettenberger, I.M.; Ramirez Bonilla, J.P.; Vinchesi-Vahl, A.C.; Weber, D.C. Striped cucumber beetle and western striped cucumber beetle (Coleoptera: Chrysomelidae). J. Int. Pest Manag. 2021, 12. [Google Scholar] [CrossRef]
- Isley, D. The striped cucumber beetle. Ark. Agric. Exp. Stat. Bull. 1927, 216, 2–35. [Google Scholar]
- Capinera, J. Family Chrysomelidae, Several Subfamilies—Leaf Beetles. In Handbook of Vegetable Pests, 2nd ed.; Fernandez, B.J., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 120–122. [Google Scholar]
- Iriis Phytoprotection: Fiche Technique de la Chrysomèle Rayée du Concombre. Available online: https://www.iriisphytoprotection.qc.ca/Fiche/Insecte?imageId=4237#:~:text=Pupe%20%3A%204%2C0%20%C3%A0%206,pattes%20sont%20jaunes%20et%20noires (accessed on 22 March 2022).
- Chittenden, F.H. The Striped Cucumber Beetle and How to Control it; US Government Print Office: Washington, DC, USA, 1923; pp. 2–6. [Google Scholar]
- Cornell University’s Insect Diagnostic Laboratory: Factsheet of the Striped Cucumber Beetle. Available online: https://ecommons.cornell.edu/bitstream/handle/1813/14356/Striped%20Cucumber%20Beetle.pdf?sequence=2 (accessed on 27 April 2022).
- Gould, G.E. The Biology and Control of the Striped Cucumber Beetle; Purdue University: West Lafayette, IN, USA, 1942; pp. 1–11. [Google Scholar]
- Le réseau d’avertissement Phytosanitaire: Factsheet on the Striped Cucumber Beetle. Available online: https://www.agrireseau.net/documents/Document_97620.pdf (accessed on 27 April 2022).
- Rojas, E.S.; Gleason, M.L.; Batzer, J.C.; Duffy, M. Feasibility of Delaying Removal of Row Covers to Suppress Bacterial Wilt of Muskmelon (Cucumis melo). Plant Dis. 2011, 95, 729–734. [Google Scholar] [CrossRef] [Green Version]
- Ellers-Kirk, C.; Fleischer, S.J. Development and Life Table of Acalymma vittatum (Coleoptera: Chrysomelidae), a Vector of Erwinia tracheiphila in Cucurbits. Environ. Entomol. 2006, 35, 875–880. [Google Scholar] [CrossRef] [Green Version]
- Couture, I. Principaux insecticides et fongicides homologués pour les cucurbitacées en 2020. Agriréseau 2020, 1, 1–25. [Google Scholar]
- Health Canada: Costumer Product Safety. Available online: https://pr-rp.hc-sc.gc.ca/ls-re/index-eng.php (accessed on 22 March 2022).
- SAgE Pesticides: Factcheet of the Toxicology of Pyrethrin. Available online: https://www.sagepesticides.qc.ca/Recherche/RechercheMatiere/DisplayMatiere?MatiereActiveID=166&searchText=pyr%C3%A9thrines&isProduct=False (accessed on 3 March 2022).
- Porcel, M.; Cotes, B.; Campos, M. Biological and behavioral effects of kaolin particle film on larvae and adults of Chrysoperla carnea (Neuroptera: Chrysopidae). Biol. Control. 2011, 59, 98–105. [Google Scholar] [CrossRef]
- Brandt, K. Organic Options for Striped Cucumber Beetle Management in Cucumbers. Master’s Thesis, Grand Valley State University, Allendale, MI, USA, June 2012. [Google Scholar]
- Duval, J. Stratégies gagnantes dans la lutte contre certains insectes ravageurs des cultures maraîchères biologiques. In Proceedings of the Centre D’expertise et de Transfert en Agriculture Biologique et de Proximité, Québec, QC, Canada, 8 February 2013. [Google Scholar]
- Ingwell, L.L.; Thompson, S.L.; Kaplan, I.; Foster, R.E. High tunnels: Protection for rather than from insect pests? Pest Manag. Sci. 2017, 73, 2439–2446. [Google Scholar] [CrossRef]
- eOrganic: Managing Cucumber Beetles in Organic Farming Systems. Available online: https://eorganic.org/node/5307 (accessed on 22 March 2022).
- University of Missouri’s Integrated Pest Management Faculty: A Novel Mass Trapping System to Control Cucumber Beetles in Cucurbit Crops. Available online: https://ipm.missouri.edu/cropPest/2016/6/A-novel-mass-trapping-system-to-control-cucumber-beetles-in-cucurbit-crops/ (accessed on 27 April 2022).
- Ternest, J.J.; Ingwell, L.L.; Foster, R.E.; Kaplan, I. Comparing Prophylactic Versus Threshold-Based Insecticide Programs for Striped Cucumber Beetle (Coleoptera: Chrysomelidae) Management in Watermelon. J. Econ. Entomol. 2020, 113, 872–881. [Google Scholar] [CrossRef]
- Bomfim, I.; Freitas, B.; Aragão, F.; Walters, F. Pollination in Cucurbit Crops. In Handbook of Cucurbits: Growth, Cultural Practices and Physiology, 1st ed.; Pessarakli, M., Ed.; CRC Press: Boca Raton, FL, USA, 2016; pp. 181–200. [Google Scholar]
- Delaplane, K.; Mayer, D.F. Squash, pumpkin and gourd. In Crop Pollination by Bees, 1st ed.; CABI Publishing: New York, NY, USA, 2000; pp. 223–226, 257–259. [Google Scholar]
- McGregor, S.E. Cucumber and gherkin, Pumpkin and squash. In Insect Pollination of Cultivated Crop Plants, 1st ed.; McGregor, S.E., Ed.; Agricultural Research Service, US Department of Agriculture: Washington, DC, USA, 1976; Volume 496, pp. 203–208, 306–310. [Google Scholar]
- Loreto, F.; Dicke, M.; Schnitzler, J.-P.; Turlings, T.C.J. Plant volatiles and the environment. Plant Cell Environ. 2014, 37, 1905–1908. [Google Scholar] [CrossRef]
- Robert, C.A.M.; Erb, M.; Hibbard, B.E.; Wade French, B.; Zwahlen, C.; Turlings, T.J.C. A specialist root herbivore reduces plant resistance and uses an induced plant volatile to aggregate in a density-dependent manner. Funct. Ecol. 2012, 26, 1429–1440. [Google Scholar] [CrossRef]
- Nunes, C.E.P.; Peñaflor, M.F.G.V.; Bento, J.M.S.; Salvador, M.J.; Sazima, M. The dilemma of being a fragrant flower: The major floral volatile attracts pollinators and florivores in the euglossine-pollinated orchid Dichaea pendula. Oecologia 2016, 182, 933–946. [Google Scholar] [CrossRef]
- Szendrei, Z.; Rodriguez-Saona, C. A meta-analysis of insect pest behavioral manipulation with plant volatiles. Entomol. Exp. Appli. 2010, 134, 201–210. [Google Scholar] [CrossRef]
- Lewis, P.A.; Lampman, R.L.; Metcalf, R.L. Kairomonal attractants for Acalymma vittatum (Coleopter: Chrysomelidae). Environ. Entomol. 1990, 19, 8–14. [Google Scholar] [CrossRef]
- Andersen, J.F.; Metcalf, R.L. Identification of a volatile attractant for Diabrotica and Acalymma spp. from blossoms of Cucurbita maxima Duchesne. J. Chem. Ecol. 1986, 12, 687–699. [Google Scholar]
- Andrews, E.S.; Theis, N.; Adler, L.S. Pollinator and Herbivore Attraction to Cucurbita Floral Volatiles. J. Chem. Ecol. 2007, 33, 1682–1691. [Google Scholar] [CrossRef]
- Metcalf, R.L.; Metcalf, E.R. Plant Kairomones in Insect Ecology and Control, 1st ed.; Chapman and Hall Ltd.: London, UK, 1992. [Google Scholar]
- Dicke, M.; Takken, W. Chemical Ecology: From Gene to Ecosystem; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2006; Volume 16, pp. 175–189. [Google Scholar]
- Light, D.M.; Knight, A.L.; Henrick, C.A.; Rajapaska, D.; Lingren, B.; Dickens, J.C.; Roitman, J. A pear-derived kairomone with pheromonal potency that attracts male and female codling moth, Cydia pomonella (L.). Naturwissenschaften 2001, 88, 333–338. [Google Scholar] [CrossRef]
- Martel, J.; Alfored, A.; Dickens, J. Synthetic host volatiles increase efficacy of trap cropping for management of Colorado potato beetle, Leptinotarsa decemlineata (Say). Agric. For. Entomol. 2005, 7, 79–86. [Google Scholar] [CrossRef]
- Piñero, J.C. A Comparative Assessment of the Response of Two Species of Cucumber Beetles (Coleoptera: Chrysomelidae) to Visual and Olfactory Cues and Prospects for Mass Trapping. J. Econ. Entomol. 2018, 111, 1439–1445. [Google Scholar] [CrossRef]
- Milosavljević, I.; Hoddle, C.D.; Mafra-Neto, A.; Gómez-Marco, F.; Hoddle, M.S. Effects of Food Bait and Trap Type on Captures of Rhynchophorus palmarum (Coleoptera: Curculionidae) and Trap Bycatch in Southern California. J. Econ. Entomol. 2020, 113, 2407–2417. [Google Scholar] [CrossRef]
- Cha, D.H.; Gill, M.A.; Epsky, N.D.; Werle, C.T.; Adamczyk, J.J., Jr.; Landolt, P.J. From a non-target to a target: Identification of a fermentation volatile blend attractive to Zaprionus indianus. J. Appl. Entomol. 2015, 139, 114–122. [Google Scholar] [CrossRef]
- Spears, L.R.; Looney, C.; Ikerd, H.; Koch, J.B.; Griswold, T.; Strange, J.P.; Ramirez, R.A. Pheromone Lure and Trap Color Affects Bycatch in Agricultural Landscapes of Utah. Environ. Entomol. 2016, 45, 1009–1016. [Google Scholar] [CrossRef] [Green Version]
- Cavanagh, A.; Hazzard, R.; Adler, L.; Boucher, J. Using trap crops for control of Acalymma vittatum (Coleoptera: Chrysomelidae) reduces insecticide use in butternut squash. J. Econ. Entomol. 2009, 102, 1101–1107. [Google Scholar] [CrossRef]
- Dobson, H.E.M.; Bernays, E. Floral volatiles in insect biology. In Insect-Plant Interactions, 1st ed.; Bernays, E.A., Ed.; CRC Press: Boca Raton, FL, USA, 1994; Volume 5, pp. 47–81. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tinslay, J.; Fournier, M.; Couture, I.; Lafontaine, P.J.; Lefebvre, M.; Lucas, E. Optimization of a Mass Trapping Method against the Striped Cucumber Beetle Acalymma vittatum in Organic Cucurbit Fields. Insects 2022, 13, 465. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050465
Tinslay J, Fournier M, Couture I, Lafontaine PJ, Lefebvre M, Lucas E. Optimization of a Mass Trapping Method against the Striped Cucumber Beetle Acalymma vittatum in Organic Cucurbit Fields. Insects. 2022; 13(5):465. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050465
Chicago/Turabian StyleTinslay, Jessee, Marc Fournier, Isabelle Couture, Pierre J. Lafontaine, Maxime Lefebvre, and Eric Lucas. 2022. "Optimization of a Mass Trapping Method against the Striped Cucumber Beetle Acalymma vittatum in Organic Cucurbit Fields" Insects 13, no. 5: 465. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050465