
Bacterial Symbionts in Ceratitis capitata
by
 , , , ,
, , , ,
Alessia Cappelli
1 ,
,
Dezemona Petrelli
2,
Giuliano Gasperi
3,
Aurelio Giuseppe Maria Serrao
1,
Irene Ricci
1,
Claudia Damiani
1,* and
Guido Favia
1 1
School of Biosciences & Veterinary Medicine, University of Camerino, CIRM Italian Malaria Network, Via Gentile III da Varano, 62032 Camerino, Italy
2
School of Biosciences & Veterinary Medicine, University of Camerino, Via Gentile III da Varano, 62032 Camerino, Italy
3
Department of Biology and Biotechnology, University of Pavia, 27100 Pavia, Italy
*
Author to whom correspondence should be addressed.
Insects 2022, 13(5), 474; https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050474
Submission received: 31 March 2022
/
Revised: 6 May 2022
/
Accepted: 17 May 2022
/
Published: 19 May 2022
(This article belongs to the Special Issue Selected Papers from the 1st International Electronic Conference on Entomology)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
The Mediterranean fly (Medfly), Ceratitis capitata, is considered one of the world’s most destructive fruit pests, as it can attack commercially important fruit, thus causing considerable economic damages, estimated to be more than 2 billion dollars annually. The yield reductions are mainly due to the damage incurred when larvae feed directly on the pulp, inducing the premature fruit drop. Additionally, oviposition holes facilitate secondary fungal and bacterial infections, further reducing the yields. Integrated pest management (IPM) strategies for medfly control are highly dependent on the use of insecticides, which, however, pose environmental concerns. Alternative strategies include the Sterile Insect Technique (SIT), which aims to eliminate or suppress pest insects without using pesticides. Lately, the medfly microbiota has been explored to develop new control strategies for insect pests and insect vectors. Here, we report the characterization of the microbial communities associated with selected organs of three different populations of C. capitata to identify possible candidates for a Symbiotic Control approach. Our findings provide new knowledge about the microbiota associated with C. capitata and stress the characterization of microbial symbionts as possible tools for a Symbiotic Control approach to implementing the pest management programs.
Abstract
Ceratitis capitata (Diptera: Tephritidae) is responsible for extensive damage in agriculture with important economic losses. Several strategies have been proposed to control this insect pest including insecticides and the Sterile Insect Technique. Traditional control methods should be implemented by innovative tools, among which those based on insect symbionts seem very promising. Our study aimed to investigate, through the 16S Miseq analysis, the microbial communities associated with selected organs in three different medfly populations to identify possible candidates to develop symbiont-based control approaches. Our results confirm that Klebsiella and Providencia are the dominant bacteria in guts, while a more diversified microbial community has been detected in reproductive organs. Concertedly, we revealed for the first time the presence of Chroococcidiopsis and Propionibacterium as stable components of the medfly’s microbiota. Additionally, in the reproductive organs, we detected Asaia, a bacterium already proposed as a tool in the Symbiotic Control of Vector-Borne Diseases. A strain of Asaia, genetically modified to produce a green fluorescent protein, was used to ascertain the ability of Asaia to colonize specific organs of C. capitata. Our study lays the foundation for the development of control methods for C. capitata based on the use of symbiont bacteria.
1. Introduction
Ceratitis capitata (Diptera: Tephritidae) is one of the main destructive pests of fruit production worldwide [1] because of its significant physical damage to fruits and vegetables and its economic impact on agriculture and forestry. Due to its ability to tolerate and adapt to a wide range of climates and its capability to attack a broad spectrum of plant species, C. capitata has a large distribution. This is the case in Mediterranean countries as well, considering that medfly damage (linked to the population density) is more consistent in intensive cultures such as in Latin America than in Mediterranean agriculture [2]. Several strategies have been proposed to control the medfly distribution. Among them, methods such as the insecticide bait spray and the Sterile Insect Technique (SIT) have been demonstrated to be effective [3,4]. Although the chemical method works efficiently in medfly control, it has some disadvantages related to its toxicity to humans and animals and makes the fruits or plants polluted by leaving residues on them. Despite the fact that SIT has been successful in several countries, releasing X-rays sterile medflies aimed to reduce the wild insect population and helping to reduce the broad spectrum of chemical treatments, new control methods that are easily applicable are urgently needed.
Indeed, the study of microbiota could open new perspectives in medfly control to integrate other control methods. In fact, in several entomological systems (both insect pests and insect vectors), the use of symbiotic microbes to control harmful insects is already common practice in some cases; in others, it may become so in the near future [5,6,7]. A striking case in this context is represented by mosquitos’ vectors of pathogens that are dangerous for humans and animals. In particular, some selected bacteria have been enrolled in mosquito control. One very effective method is the biocontrol of mosquito borne diseases through mosquitoes transinfected with the endosymbiotic bacteria Wolbachia. Others refer to the bacterium Asaia, which has been isolated from many different mosquito species. Asaia is a natural effector for mosquito immune priming and is often detected at a very high prevalence, and this is attributed to several aspects of its biology. It has also been proposed for paratransgenic control applications [8,9].
Nevertheless, up to now, studies on the microbiota composition of C. capitata are already limited [10,11,12,13,14] and mostly related to the SIT and Incompatible Insect Technique (IIT). It has been noted that these procedures affect the medfly gut bacterial community, and sterile males are less competent in attracting and mating with wild females and are affected by the genetic background of the medfly population [15,16,17]. Post-irradiation dietary supplementation with Klebsiella oxytoca improved the mating latency of infertile males, suggesting the potential use of this bacterium to improve the SIT success [15]. Additionally, selected bacteria can act as probiotics in the diet of larval and adult stages, improving C. capitata mass rearing and enhancing the SIT application [18]. Similar to C. capitata, the intestinal probiotic K. oxytoca restored the ecological fitness of Bactrocera dorsalis, another destructive, polyphagous and invasive insect pest of fruits and vegetables, following a decline post-irradiation [19]. In B. dorsalis bacteria belonging to Enterobacteriaceae, Enterobacter cloacae, K. oxytoca, Morganella sp., Providencia rettgerii and C. freundii were identified as predominant in the gut, suggesting their involvement in promoting host fitness under stressful conditions and host immune system response [20,21,22]. K. michiganensis was implicated in promoting insect resistance to long-term, low-temperature stress [23]. Moreover, the gut symbiont Citrobacter sp. seemed to be involved in increasing the host resistance against an organophosphate insecticide [20].
Here, we present the results of a study aimed to characterize the microbial communities in different anatomical districts (guts and reproductive organs) of three different populations of C. capitata to determine whether selected symbionts could be translated into potential tools for the Symbiotic Control (SC) of medfly, which could be integrated with more traditional approaches. Considering the presence of Asaia in all three medfly populations, we investigate the possibility of using Asaia in the SC of C. capitata.
2. Materials and Methods
2.1. Ceratitis capitata Rearing
The strains of C. capitata used in this work were: (i) the Guatemala strain, established in 1989 from wild pupae collected in Antigua (Guatemala); (ii) the La Réunion strain, established in 1994 from wild pupae collected near St. Denis (La Reunion, France); (iii) the ISPRA strain, established in 1968 at the European Community Joint Research Centre (Ispra, Italy) with wild flies from Sicily and Greece and maintained in Pavia since 1979. These strains originated from the Department of Biology & Biotechnology, University of Pavia, where they are maintained under standard rearing conditions [24], and, since 2018, they have been maintained in the insectary at the School of Biosciences and Veterinary Medicine, University of Camerino. The insects were kept in cages (25 × 25 × 25 cm) made of a steel frame covered with nettings and maintained at the standard lighting conditions of 12 h light and 12 h dark, 26 ± 1 °C temperature and 75 ± 5% humidity and in aseptic conditions during both the developmental and adult stages.
2.2. DNA Extraction
Before DNA extraction, the insect surface was sterilized in 70% ethanol and rinsed three times in sterile PBS. The samples were homogenized with sterile 0.5-mm wide glass beads (Bertin Instrument, Montigny Le-Bretonneux, France) for 30 s at 6800 rpm by an automatic tissue homogenizer (Precellys 24 Bertin Instrument, Montigny Le-Bretonneux, France). Genomic DNA was extracted using a Jet Flex Genomic DNA Purification kit (Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA), according to the manufacturer’s instructions.
2.3. 16S rRNA Profiling
The 16S Miseq analysis was conducted on the male and female organs (gut and reproductive organs) of three different C. capitata populations. Single pools of 20 organs for each group were obtained from cohorts of 10-day-old insects dissecting in sterile conditions. DNA extraction was performed as described above. A negative control consisting of a blank sample was included for each batch of extraction to control for the contamination of bacteria possibly introduced during the DNA extraction. They were not further processed since no quantifiable extract was produced from each negative control.
16S rRNA profiling was conducted by LGC Genomics (Berlin, Germany). Libraries preparation was performed by covering the hypervariable region V3–V4 of 16S ribosomal RNA using 341F and 785R oligonucleotides [25]. The data were pre-processed using the Illumina bcl2fastq 2.17.1.14 software, and the reads were sorted by amplicon inline barcodes. Sequencing adapter remnants were clipped from all reads. 16S pre-processing and OTU picking from the amplicons were analyzed using Mothur 1.35.1 [26]. The sequence alignments were performed against the 16S Mothur-Silva SEED r119 reference alignment. OTU diversity was analyzed with QIIME 1.9.0 [27], and annotations of the putative species level of OTUs were obtained with NCBI BLAST+ 2.2.29 [28]. The raw data were submitted as BioProject accession number PRJNA682004 to the NCBI database.
2.4. Molecular Detection of Asaia
The presence of Asaia was investigated in 20 female and 20 male adults from the three C. capitata populations. Asaia detection was performed by PCR using specific oligonucleotides targeting the 270-bps fragment of the 16S rRNA gene described in Favia et al. [8]. A total of 50 ng of genomic DNA were used in PCR reaction containing 1X Buffer, 0.25 mM dNTPs, 0.9 U DreamTaq Polymerase (Thermo Scientific, Waltham, MA, USA) and 200 nM of Asafor (5′-GCGCGTAGGCGGTTTACAC-3′) and Asarev (5′-AGCGTCAGTAATGAGCCAGGTT-3′) oligonucleotides. The amplification protocol included: initial denaturation at 95 °C for 3 min, followed by 30 cycles consisting of denaturation at 95 °C for 30 s, annealing at 60 °C for 30 s and extension at 72 °C for 30 s and concluding with a final extension step of 10 min at 72 °C. The PCR products were electrophoresed on a 1% agarose gel to determine the presence and size of the amplified DNA. The amplicons were purified and sequenced by the mean of the Sanger method (Eurofins Genomics, Ebersberg, Germany). The 16S RNA sequences were analyzed by BLASTN (http://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 1 May 2022).
2.5. Asaia Isolation from C. capitata Adults
Bacteria isolation was performed from C. capitata adults. Before the isolation, the insect surface was sterilized in 70% ethanol and rinsed three times in sterile PBS. Asaia isolation was performed as described in Favia et al. [8] using a liquid enrichment medium at pH 3.5 followed by plating in a carbonate-containing solid medium. The enrichment medium allowed for the elimination of those microorganisms not tolerating low pH, while the following plating on CaCO3-containing medium permitted the isolation of acid-producing bacteria capable of creating CaCO3 dissolution haloes around the colonies. To confirm the isolation, pink-pigmented, shiny and smooth colonies were analyzed though Asaia-specific PCR.
2.6. Asaia sp. Transformation
Asaia strains isolated from the three populations of C. capitata were transformed with plasmids pHM2 containing a GFP gene cassette to express a green phenotype, as described in Favia et al. [8]. Asaia cells were grown overnight on GLY agar (yeast extract 1%, glycerol 2.5%, agar 2%, pH 5) for 48 h at 30 °C. The culture was diluted 1:20 into 25 mL of GLY and incubated with aeration until the cells reached the early log phase (optical density at 600 nm, 0.5–0.8). After an incubation of 15 min on ice, the cells were harvested (2700× g, 10 min, 4 °C) and washed twice with 10 mL of cold 1 mM HEPES (pH 7.0). The pellet was resuspended in 5 mL of cold 10% (v/v) glycerol, centrifuged again and finally resuspended in 0.5 mL of cold 10% (v/v) glycerol. Then, 65 microliters of competent cells (≈9.2 × 109 CFU/mL) were mixed with 0.2 μg of plasmid DNA, transferred to a cold 0.1 cm-diameter cuvette and pulsed at 1800 V in the Electroporator apparatus (Biorad, Hercules, CA, USA). The cells were immediately added with 1 mL of GLY medium, transferred to a tube and incubated at 30 °C for 3 h. Transformant cells were selected by plating on GLY agar medium added with 100 μg/mL kanamycin, 40 μg/mL 5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside (XGal) 24 and 0.5 mM isopropyl-β-D-thiogalactopyranoside (IPTG) for testing the Lac+ phenotype. Successful transformants were confirmed by fluorescence microscopy analysis and specific Asaia-PCR. A Gfp-tagged transformant from the ISPRA population was hence selected for mosquito recolonization experiments.
2.7. Medfly Recolonization by Asaia Expressing Green Fluorescent Protein (Gfp)
The ISPRA population was fed with modified Asaia to evaluate the ability of the bacterium to colonize the insect host. The AsaiaGfp strain from the ISPRA medfly was grown for 24 h at 30 °C in GLY medium enriched with 100 µg/mL kanamycin. The cells were harvested by centrifugation, washed three times in 0.9% NaCl, adjusted to a concentration of 108 cells/mL and resuspended in sugar solution 25%. For monitoring the long-term colonization of C. capitata, the suspension was supplemented with 100 μg/mL of kanamycin to avoid the loss of plasmid from the bacterial cells. After 3, 5, 10 and 15 days of exposure to the feeding solutions containing AsaiaGfp, the insects were dissected to monitor the presence of green bacteria in the organs. At each timepoint, the guts and reproductive organs of 7 males and 7 females were dissected in sterile PBS, fixed with 4% paraformaldehyde for 10 min at 4 °C and mounted in glycerol–PBS for analysis with a Nikon C2+ Laser Scanning Confocal Microscope (Nikon, Minato, Tokyo, Japan).
2.8. Isolation of Bacteria from the ISPRA Population
Bacteria from the ISPRA population medfly were isolated, processing a single pool of 10 gut and 10 reproductive organs from males and females. Each pool was homogenized as previously described in 100 µL of sterile 1X PBS, and serial dilution in 1X PBS was prepared from 1:10 to 1:1000. One hundred microliters of each dilution were spread on the surface of Blood Agar plates (ThermoFisher Scientific, Waltham, MA, USA) and incubated at 35 °C for 48 h. The isolated colonies were picked and transferred on Luria Bertani (LB) agar plates (1% NaCl, 1% Tryptone, 0.5% Yeast extract, 2% Agar, pH7). To isolate gram-negative bacteria, clones were plated on MacConkey agar (ThermoFisher Scientific, Waltham, MA, USA) and incubated at 35 °C for 48 h. The identification of gram-negative oxidase-negative isolates was performed using the API20E system (bioMérieux, Inc, Marcy-l’Étoile, France).
The other isolates were molecularly characterized using universal oligonucleotides targeting the 16S RNA ribosomal bacterial (805 Rev 5′-TCGACATCGTTTACGGCGTG-3′ and 27 for 5′-AGAGTTTGATCCTGGCTCAG-3′). A total of 50 ng of genomic DNA were used in PCR reaction containing 1× Buffer, 0.25 mM dNTPs, 0.9 U DreamTaq Polymerase (Thermo Scientific, Waltham, MA, USA), 200 nM of 27F and 805R oligonucleotides. The amplification protocol included: initial denaturation at 95 °C for 3 min, followed by 30 cycles consisting of denaturation at 95 °C for 30 s, annealing at 56 °C for 30 s and extension at 72 °C for 30 s and concluding with a final extension step of 10 min at 72 °C. The PCR products were electrophoresed on a 1% agarose gel to determine the presence and size of the amplified DNA. The amplicons were purified and sequenced by the mean of the Sanger method (Eurofins Genomics, Germany). The 16S RNA sequences were analyzed by BLASTN (http://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 1 May 2022).
2.9. Antagonistic Assay
Antagonistic activity of the Klebsiella oxytoca strain was evaluated by an Agar diffusion assay against the Asaia strain isolated from the ISPRA population medfly as described in Ciolfi and Marri [29], with some modifications. Briefly, overnight cultures of K. oxitoca and Asaia in a GLY medium were used for the test. A lawn was prepared by mixing 5 mL of soft-agar (0.7% agar) with 59 μL of Asaia culture (OD600, 0.1) and poured over the GLY agar medium in plates. Three microliters of K. oxytoca (OD600, 0.3) were then spotted on the lawn. The plates were incubated for 4 days at 26 °C and examined daily for zones of inhibition.
3. Results
3.1. 16S rRNA Profiling
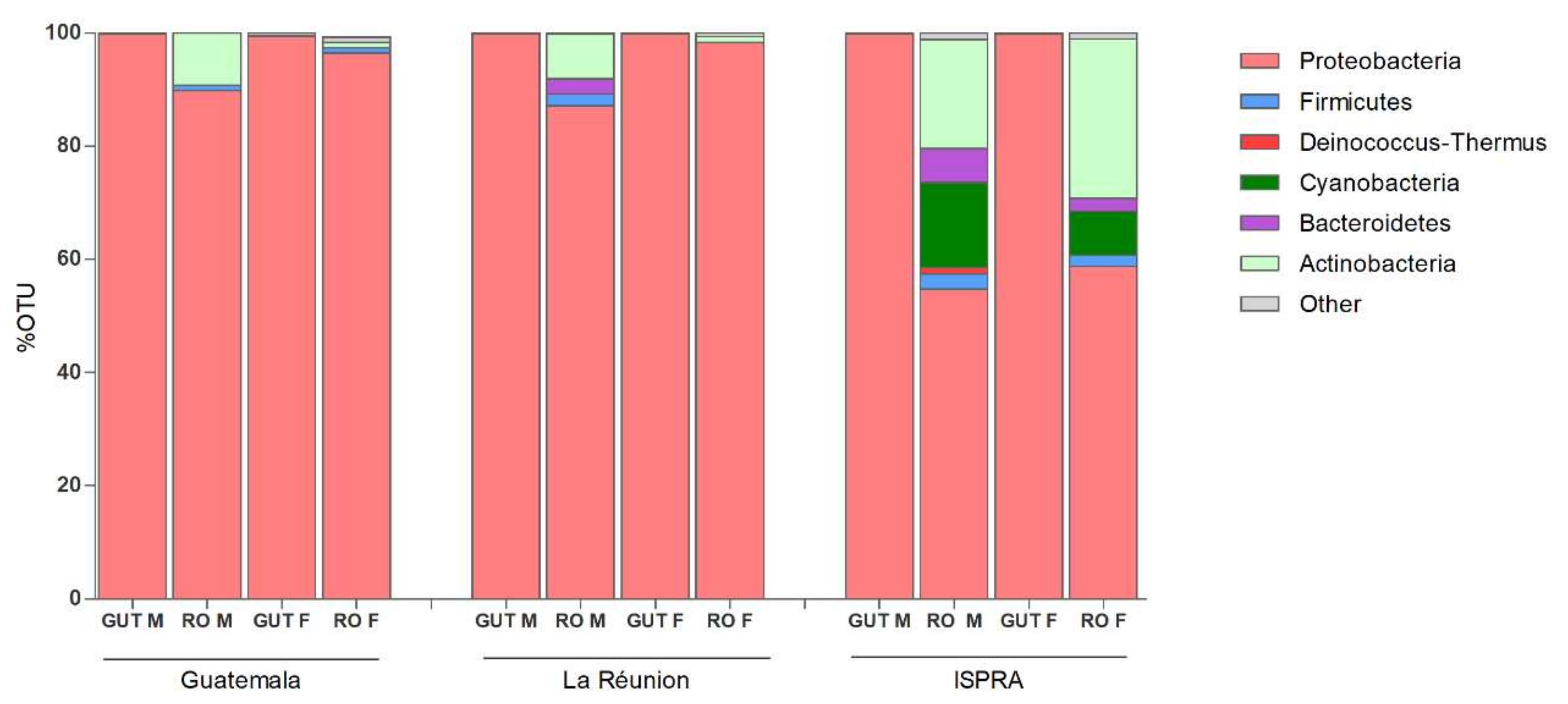
The microbiome sequencing of male and female organs of three different populations of C. capitata generated a total of 4.2 M reads, varying among samples (minimum = 64.760, maximum = 701.290), with an average of 352.944 reads. Analysis of the rarefaction curves indicated an adequate sampling quality, suggesting a coherent number of sequence reads per sample. The Principal Coordinates Analysis (PCoA) plots show the high similarity of microbial composition among all of the guts analyzed, while a more specific microbial community in the ISPRA reproductive organs was represented. No substantial difference in microbial composition is observed between males and females in any strain (Figure S1). At the phylum level, the Proteobacteria results are the most prevalent in all groups, particularly in guts (males and females 99%). The phyla Actinobacteria and Firmicutes were detected in reproductive organs. Bacteroides phylum was revealed in the reproductive organs of La Réunion (males: 2.7%) and ISPRA strains (male: 6% and females: 2.3%). Additionally, Cyanobacteria phylum was present in ISPRA population reproductive organs (males: 15% and females: 7.8%) (Figure 1 and Table S1).
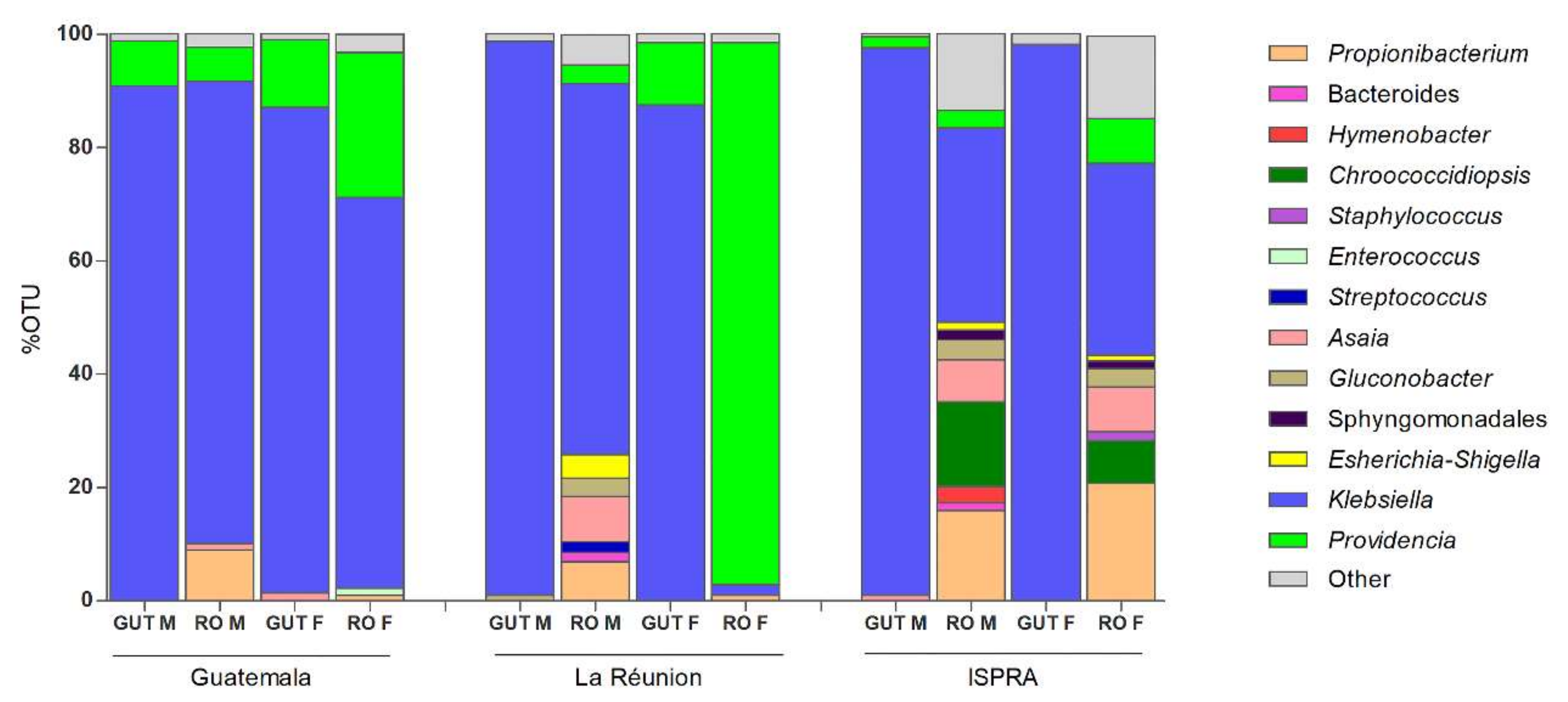
At the genus level, among the Proteobacteria phylum, Klebsiella, belonging to the class of Gammaproteobacteria, was the most abundant bacteria, with a range around 85–98% in the guts and 33–80% in the male and female reproductive organs of all three populations, except in the La Rèunion females, where Providencia is highly represented (95.7%), while Klebsiella is at 1.8%. Providencia was detected with a different percentage range (6–95%) in all samples—although in the male guts of the La Réunion strain and the female guts of the ISPRA strain, the percentage was lower than 1% (0.3% and 0.5%, respectively) (Figure 2 and Figure S2).
Other interesting bacteria belonging to the phylum Proteobacteria (class: Alphaproteobacteria) such as Asaia and Gluconobacter were also detected, which were present in all samples, albeit with variable percentages (see Supplementary Materials Table S2). Considering the 1% cut-off for sample analysis, Asaia was present in the male reproductive organs (1.1%) and female guts (1.4%) in the Guatemala strain, the male reproductive organs (8%) of the La Réunion strain and the male guts (1%) and male (7.4%) and female (8%) reproductive organs of the ISPRA strain.
Gluconobacter was mostly detected in the males (guts: 1% and reproductive organs: 3.2%) of the La Réunion population and in the male (3.6%) and female (3.2%) reproductive organs of the ISPRA strain.
Just in the ISPRA population, the bacterium Chroococcidiopsis, belonging to the phylum Cyanobacteria, reached a much higher density, being detected in male (14.9%) and female (7.6%) reproductive organs.
In all three populations, Propionibacterium was detected only in male and female reproductive organs, with a range of 1–20%.
As already reported in several studies, no Wolbachia was detected in any samples [30].
3.2. Detection, Isolation and Transformation of Asaia from C. capitata Populations
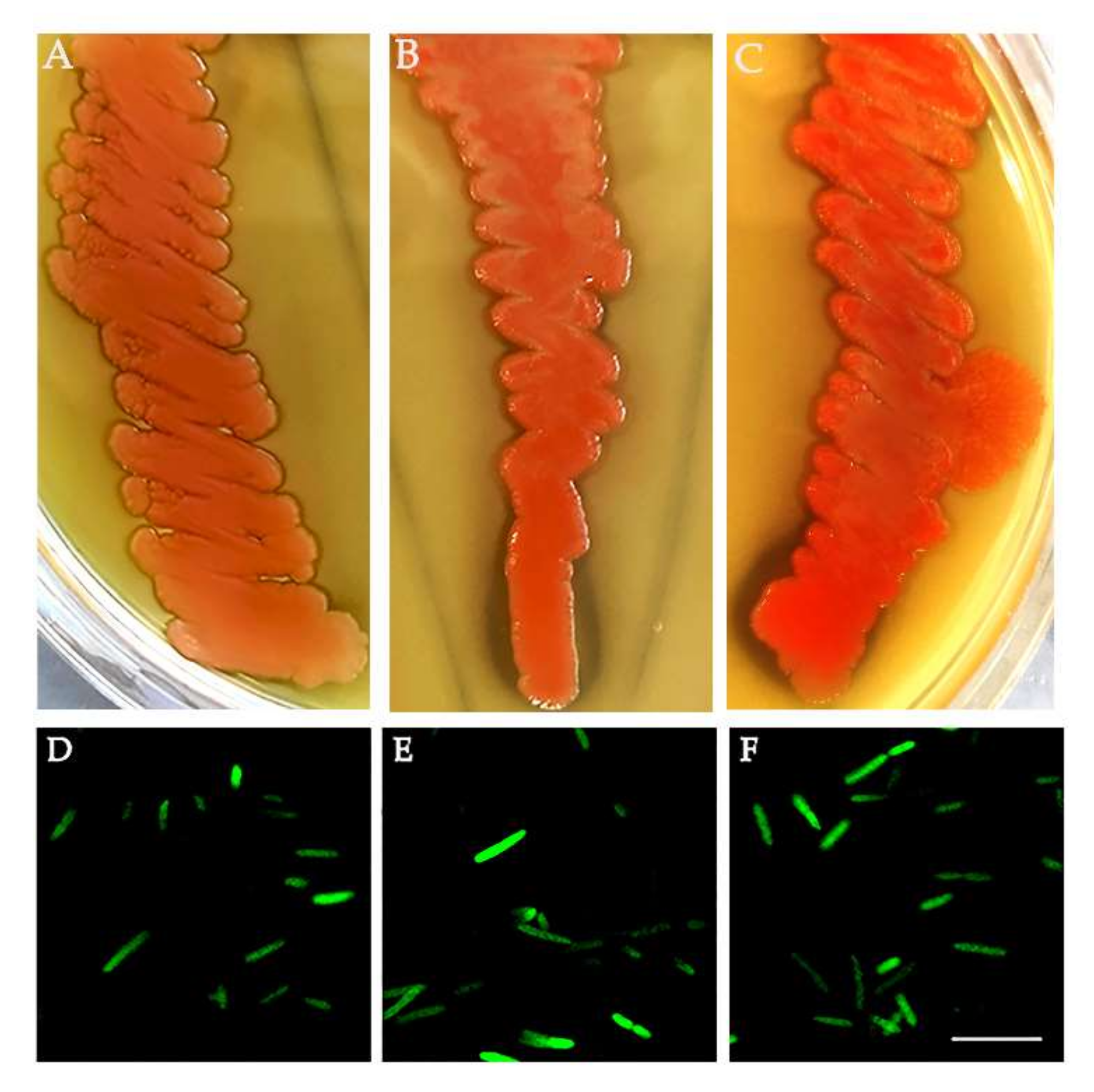
Considering the presence of Asaia shown in the NGS analysis, we investigated its presence in the new generation of medfly in depth. We tested for Asaia presence in 40 adults (20 males and 20 females) of each population by specific PCR. Asaia was detected in 100% of samples (a total of 120 individuals). Moreover, Asaia was isolated from C. capitata adults using a selected medium followed by plating in a carbonate-rich medium. The resulting single, pink colonies capable of dissolving carbonate in the medium and generating dissolution haloes were confirmed to be Asaia by 16S rRNA gene sequencing, with 99.8% nucleotide identity with A. bogorensis (Figure 3A–C). Additionally, Asaia was genetically modified to express a GFP protein to confirm its versability (Figure 3D–F).
3.3. Medfly Recolonization Using Asaia Strain Expressing Green Fluorescent Protein (Gfp)
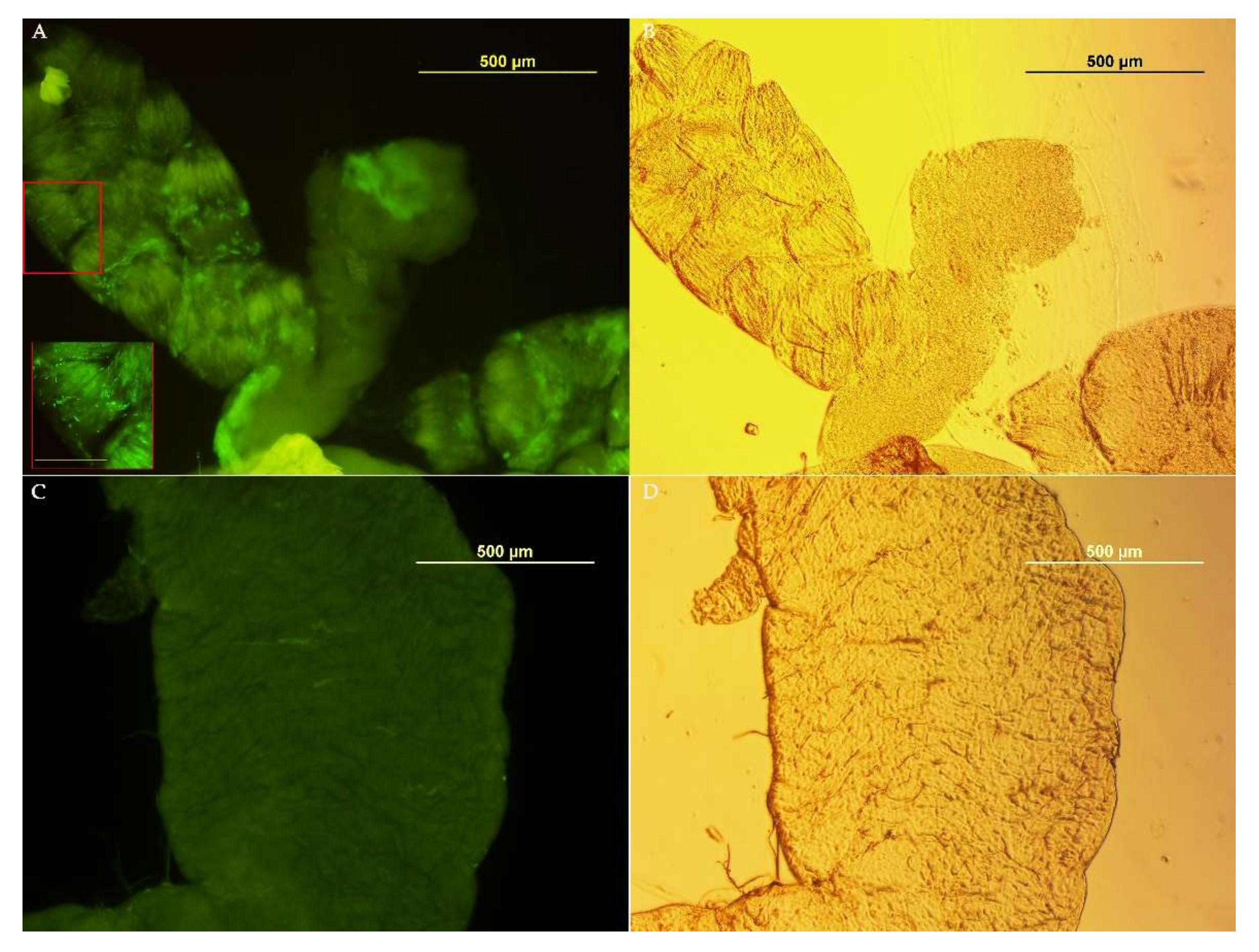
To investigate the possibility of using Asaia as a Symbiotic Control tool for medfly, we tested its ability to colonize the insect’s organs in the ISPRA population, where Asaia was most abundant in the reproductive organs with respect to the other populations. Asaia-GFP was observed in the crop, an initial portion of the alimentary tract, of all samples (males: 7/7 and females: 7/7) only at the first time point (3 days after Asaia administration) (Figure 4). At the following time points, no individual tested positive for fluorescence. Likely, the presence of strong dominant bacteria, such as Klebsiella and Providencia, and the unfavorable sugary diet may have prevented Asaia from colonizing medfly organs in the long term.
3.4. Bacteria Isolation from the ISPRA Population and Activity Test
Considering the competition phenomena of symbionts to colonize specific niches in insects [31,32], we proceeded with the isolation of other bacteria from male and female organs (guts and reproductive organs) to select microorganisms showing potential features to interfere with the presence of Asaia. From each pool of guts plated on MacConkey Agar, Klebsiella oxytoca (Enterobacteriaceae family) and Serratia marcescens (Yersiniaceae) were isolated. The same species were found in the reproductive organs of both males and females while, Aeromonas spp. (Aeromonadaceae) was detected only in female reproductive organs. Moreover, the sequencing of a longer fragment of the16S rRNA gene (780 bps) of other isolates allowed for the identification of the bacterium Enterococcus sp. (Enterococcaceae), with an identity of 99% (MK764705.1) in the same organs.
Since K. oxytoca has been described to have a killer phenotype [29], we hypothesized that the strong presence of this bacterium in the gut could interfere with Asaia stabilization in this anatomical district. We tested the killer activity of K. oxytoca against Asaia, plating the tester bacterium in soft agar and adding a drop of K. oxytoca in the center. After 4 days at 26 °C, we observed a homogeneous layer of Asaia grown around the spot of Klebsiella, suggesting that Asaia was not sensible to the K. oxytoca activity (Figure S2).
4. Discussion
The microbiota composition of three different populations of C. capitata, which were reared under the same conditions, showed a homogeneous microbial community in the guts, which is dominated by Klebsiella and Providencia, as reported in previous studies [29,33,34,35], while highly different microbial communities were detected in the reproductive organs. These data support the need to investigate bacterial communities associated with different anatomical districts to identify potentially useful microorganisms in the development of medfly Symbiotic Control approaches. In this context, the first detection of Chroococcidiopsis and Propionibacterium as stable components of the medfly’s microbiota may open interesting perspectives. Propionibacterium has been already described as part of the microbiota of other insects, such as mosquitos [36], likely accomplishing a possible nutritional role, while Chroococcidiopsis has been never detected in insects, and its role in the reproductive organs is still unknown.
Among other interesting bacteria detected in our studies, we focused on Klebsiella oxytoca and Enterococcus sp, which were isolated from guts and reproductive organs. Enterococcus was previously described as part of the microbiota of Bactrocera dorsalis, where it seemed involved, together with K. oxytoca, in the reproduction and survival of the insect pest [37]. Klebsiella sp. has been previously described as a major component of wild medfly’s intestine [38], likely involved in nitrogen fixation [38,39] and associated with male mating success [40]. Additionally, K. oxytoca strains isolated from the alimentary tract of wild medflies showed an antagonistic activity in vitro against other bacteria such as Escherichia coli, Enterobacter cloacae and Salmonella typhimurium [29].
This is particularly interesting in the light of the fact that we detected Asaia in all of the samples but located only in the reproductive organs. In fact, Asaia, an insect symbiont proposed as a tool for the control of mosquito-borne diseases by direct paratransgenic applications and indirectly through the upregulation of the host immune response [9], has been described as strictly associated with several insects, such as mosquitoes, in association with salivary glands, guts and reproductive organs [8]. In some mosquito strains, Asaia competes with Wolbachia in the reproductive organs [41,42].
Thus, a possible competition between Klebsiella and Asaia could have been hypothesized. However, the strain of Asaia that we have isolated from C. capitata did not show susceptibility to K. oxytoca in the antagonistic assay. Although Asaia is resistant to antagonistic activity, we can hypothesize that interspecies competition among other bacterial populations for nutritional sources could explain the inability of Asaia to colonize the gut of medfly, but further investigations are needed to better understand these competitive phenomena.
5. Conclusions
Despite the growing interest in the role of the microbiota in the biology of many insects and in the possible use of microbiota components for the development of innovative control methods, only a few studies have been published to date on the microbiota of Ceratitis capitata. Our study, although still intended as a preliminary, has identified some bacteria that could potentially be used for the development of control methods based on symbionts and which, at the same time, offer the possibility of better understanding some aspects of microbial competition between the symbionts of insects.
Supplementary Materials
The following supporting information can be downloaded at: https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/insects13050474/s1, Figure S1: Principal Coordinates Analysis (PCoA) plots of samples colored according to different organs; Figure S2: Antagonistic test of Klebsiella oxytoca against Asaia using the Agar diffusion assay; Table S1: % OTU Phyla in C. capitata strains; Table S2: % OTU Genera in C. capitata strains.
Author Contributions
Conceptualization, C.D. and G.F.; methodology, A.C.; D.P.; C.D. and A.G.M.S.; validation, C.D.; I.R.; G.G. and G.F.; formal analysis, A.C.; D.P. and C.D.; resources, G.G.; data curation, A.C.; C.D.; I.R. and A.G.M.S.; writing—original draft preparation, A.C.; writing—review and editing, C.D. and G.F.; supervision, C.D.; project administration, G.F.; funding acquisition, G.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the University of Camerino (FAR2019) to G.F.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All the reads related to the 16S Miseq analysis (Bioproject PRJNA682004) have been deposited in The EMBL Nucleotide Sequence Database (NCBI).
Conflicts of Interest
The authors declare no conflict of interest.
References
- White, I.M.; Elson-Harris, M.M. Fruit Flies of Economic Significance: Their Identification and Bionomics; CABI Publishing: Wallingford, UK, 1992. [Google Scholar]
- Malacrida, A.R.; Gomulski, L.M.; Bonizzoni, M.; Bertin, S.; Gasperi, G.; Guglielmino, C.R. Globalization and fruitfly invasion and expansion: The medfly paradigm. Genetica 2007, 131, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Plá, I.; García de Oteyza, J.; Tur, C.; Martínez, M.; Laurín, M.C.; Alonso, E.; Martínez, M.; Martín, Á.; Sanchis, R.; Navarro, M.C.; et al. Sterile Insect Technique Programme against Mediterranean Fruit Fly in the Valencian Community (Spain). Insects 2021, 12, 415. [Google Scholar] [CrossRef] [PubMed]
- Vreysen, M.J.B.; Abd-Alla, A.M.M.; Bourtzis, K.; Bouyer, J.; Caceres, C.; de Beer, C.; Oliveira Carvalho, D.; Maiga, H.; Mamai, W.; Nikolouli, K.; et al. The Insect Pest Control Laboratory of the Joint FAO/IAEA Programme: Ten Years (2010–2020) of Research and Development, Achievements and Challenges in Support of the Sterile Insect Technique. Insects 2021, 12, 346. [Google Scholar] [CrossRef] [PubMed]
- Utarini, A.; Indriani, C.; Ahmad, R.A.; Tantowijoyo, W.; Arguni, E.; Ansari, M.R.; Supriyati, E.; Wardana, D.S.; Meitika, Y.; Ernesia, I.; et al. Efficacy of Wolbachia-Infected Mosquito Deployments for the Control of Dengue. N. Engl. J. Med. 2021, 384, 2177–2186. [Google Scholar] [CrossRef]
- Li, H.; Wei, Y.; Yuan, G.; Guan, R. Insight into maize gene expression profiles responses to symbiotic bacteria derived from Helicoverpa armigera and Ostrinia furnacalis. Arch. Microbiol. 2021, 204, 56. [Google Scholar] [CrossRef]
- Polenogova, O.V.; Noskov, Y.A.; Artemchenko, A.S.; Zhangissina, S.; Klementeva, T.N.; Yaroslavtseva, O.N.; Khodyrev, V.P.; Kruykova, N.A.; Glupov, V.V. Citrobacter freundii, a natural associate of the Colorado potato beetle, increases larval susceptibility to Bacillus thuringiensis. Pest. Manag. Sci. 2022. [Google Scholar] [CrossRef]
- Favia, G.; Ricci, I.; Damiani, C.; Raddadi, N.; Crotti, E.; Marzorati, M.; Rizzi, A.; Urso, R.; Brusetti, L.; Borin, S.; et al. Bacteria of the genus Asaia stably associate with Anopheles stephensi, an Asian malarial mosquito vector. Proc. Natl. Acad. Sci. USA 2007, 104, 9047–9051. [Google Scholar] [CrossRef] [Green Version]
- Cappelli, A.; Damiani, C.; Mancini, M.V.; Valzano, M.; Rossi, P.; Serrao, A.; Ricci, I.; Favia, G. Asaia Activates Immune Genes in Mosquito Eliciting an Anti-Plasmodium Response: Implications in Malaria Control. Front. Genet. 2019, 10, 836. [Google Scholar] [CrossRef] [Green Version]
- Behar, A.; Yuval, B.; Jurkevitch, E. Gut bacterial communities in the Mediterranean fruit fly (Ceratitis capitata) and their impact on host longevity. J. Insect Physiol. 2008, 54, 1377–1383. [Google Scholar] [CrossRef]
- Aharon, Y.; Pasternak, Z.; Ben Yosef, M.; Behar, A.; Lauzon, C.; Yuval, B.; Jurkevitch, E. Phylogenetic, metabolic, and taxonomic diversities shape mediterranean fruit fly microbiotas during ontogeny. Appl. Environ. Microbiol. 2013, 79, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Malacrinò, A.; Campolo, O.; Medina, R.F.; Palmeri, V. Instar- and host-associated differentiation of bacterial communities in the Mediterranean fruit fly Ceratitis capitata. PLoS ONE 2018, 13, e0194131. [Google Scholar] [CrossRef] [PubMed]
- Zaada, D.S.Y.; Ben-Yosef, M.; Yuval, B.; Jurkevitch, E. The host fruit amplifies mutualistic interaction between Ceratitis capitata larvae and associated bacteria. BMC Biotechnol. 2019, 19, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikolouli, K.; Augustinos, A.A.; Stathopoulou, P.; Asimakis, E.; Mintzas, A.; Bourtzis, K.; Tsiamis, G. Genetic structure and symbiotic profile of worldwide natural populations of the Mediterranean fruit fly, Ceratitis capitata. BMC Genet. 2020, 21, 128. [Google Scholar] [CrossRef] [PubMed]
- Ben Ami, E.; Yuval, B.; Jurkevitch, E. Manipulation of the microbiota of mass-reared Mediterranean fruit flies Ceratitis capitata (Diptera: Tephritidae) improves sterile male sexual performance. ISME J. 2010, 4, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Zabalou, S.; Apostolaki, A.; Livadaras, I.; Franz, G.; Robinson, A.S.; Savakis, C.; Bourtzis, K. Incompatible insect technique: Incompatible males from a Ceratitis capitata genetic sexing strain. Entomol. Exp. Appl. 2009, 132, 232–240. [Google Scholar] [CrossRef]
- Kyritsis, G.A.; Augustinos, A.A.; Livadaras, I.; Cáceres, C.; Bourtzis, K.; Papadopoulos, N.T. Medfly-Wolbachia symbiosis: Genotype x genotype interactions determine host’s life history traits under mass rearing conditions. BMC Biotechnol. 2019, 19, 96. [Google Scholar] [CrossRef] [Green Version]
- Kyritsis, G.A.; Augustinos, A.A.; Cáceres, C.; Bourtzis, K. Medfly Gut Microbiota and Enhancement of the Sterile Insect Technique: Similarities and Differences of Klebsiella oxytoca and Enterobacter sp. AA26 Probiotics during the Larval and Adult Stages of the VIENNA 8 D53+ Genetic Sexing Strain. Front. Microbiol. 2017, 8, 2064. [Google Scholar] [CrossRef]
- Cai, Z.; Yao, Z.; Li, Y.; Xi, Z.; Bourtzis, K.; Zhao, Z.; Bai, S.; Zhang, H. Intestinal probiotics restore the ecological fitness decline of. Evol. Appl. 2018, 11, 1946–1963. [Google Scholar] [CrossRef]
- Cheng, D.; Guo, Z.; Riegler, M.; Xi, Z.; Liang, G.; Xu, Y. Gut symbiont enhances insecticide resistance in a significant pest, the oriental fruit fly Bactrocera dorsalis (Hendel). Microbiome 2017, 5, 13. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Jin, L.; Zhang, H. Comparison of the diversity of the bacterial communities in the intestinal tract of adult Bactrocera dorsalis from three different populations. J. Appl. Microbiol. 2011, 110, 1390–1401. [Google Scholar] [CrossRef]
- Muniz, L.R.; Knosp, C.; Yeretssian, G. Intestinal antimicrobial peptides during homeostasis, infection, and disease. Front. Immunol. 2012, 3, 310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raza, M.F.; Wang, Y.; Cai, Z.; Bai, S.; Yao, Z.; Awan, U.A.; Zhang, Z.; Zheng, W.; Zhang, H. Gut microbiota promotes host resistance to low-temperature stress by stimulating its arginine and proline metabolism pathway in adult Bactrocera dorsalis. PLoS Pathog. 2020, 16, e1008441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saul, S. Rearing methods for the medfly, Ceratitis capitata. Ann. Entomol. Soc. Am. 1982, 75, 480–483. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Ciolfi, S.; Marri, L. Dominant symbiotic bacteria associated with wild medfly populations reveal a bacteriocin-like killing phenotype: A ‘cold-case’ study. Bull. Entomol. Res. 2020, 110, 457–462. [Google Scholar] [CrossRef]
- Zabalou, S.; Riegler, M.; Theodorakopoulou, M.; Stauffer, C.; Savakis, C.; Bourtzis, K. Wolbachia-induced cytoplasmic incompatibility as a means for insect pest population control. Proc. Natl. Acad. Sci. USA 2004, 101, 15042–15045. [Google Scholar] [CrossRef] [Green Version]
- Itoh, H.; Jang, S.; Takeshita, K.; Ohbayashi, T.; Ohnishi, N.; Meng, X.Y.; Mitani, Y.; Kikuchi, Y. Host-symbiont specificity determined by microbe-microbe competition in an insect gut. Proc. Natl. Acad. Sci. USA 2019, 116, 22673–22682. [Google Scholar] [CrossRef]
- Shukla, S.P.; Vogel, H.; Heckel, D.G.; Vilcinskas, A.; Kaltenpoth, M. Burying beetles regulate the microbiome of carcasses and use it to transmit a core microbiota to their offspring. Mol. Ecol. 2018, 27, 1980–1991. [Google Scholar] [CrossRef] [PubMed]
- Jurkevitch, E. Riding the Trojan horse: Combating pest insects with their own symbionts. Microb. Biotechnol. 2011, 4, 620–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Cock, M.; Virgilio, M.; Vandamme, P.; Augustinos, A.; Bourtzis, K.; Willems, A.; De Meyer, M. Impact of Sample Preservation and Manipulation on Insect Gut Microbiome Profiling. A Test Case with Fruit Flies (Diptera, Tephritidae). Front. Microbiol. 2019, 10, 2833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Cock, M.; Virgilio, M.; Vandamme, P.; Bourtzis, K.; De Meyer, M.; Willems, A. Comparative Microbiomics of Tephritid Frugivorous Pests (Diptera: Tephritidae) From the Field: A Tale of High Variability Across and Within Species. Front. Microbiol. 2020, 11, 1890. [Google Scholar] [CrossRef] [PubMed]
- Muturi, E.J.; Ramirez, J.L.; Rooney, A.P.; Kim, C.H. Comparative analysis of gut microbiota of mosquito communities in central Illinois. PLoS Negl. Trop. Dis. 2017, 11, e0005377. [Google Scholar] [CrossRef]
- Akami, M.; Ren, X.M.; Qi, X.; Mansour, A.; Gao, B.; Cao, S.; Niu, C.Y. Symbiotic bacteria motivate the foraging decision and promote fecundity and survival of Bactrocera dorsalis (Diptera: Tephritidae). BMC Microbiol. 2019, 19, 229. [Google Scholar] [CrossRef] [Green Version]
- Behar, A.; Yuval, B.; Jurkevitch, E. Enterobacteria-mediated nitrogen fixation in natural populations of the fruit fly Ceratitis capitata. Mol. Ecol. 2005, 14, 2637–2643. [Google Scholar] [CrossRef]
- Behar, A.; Yuval, B.; Jurkevitch, E. Community Structure of the Mediterranean Fruit Fly Microbiota: Seasonal and Spatial Sources of Variation. Isr. J. Ecol. Evol. 2008, 54, 181–191. [Google Scholar] [CrossRef]
- Ben-Yosef, M.; Jurkevitch, E.; Yuval, B. Effect of bacteria on nutritional status and reproductive success of the Mediterranean fruit fly Ceratitis capitata. Physiol. Entomol. 2008, 33, 145–154. [Google Scholar] [CrossRef]
- Hughes, G.L.; Dodson, B.L.; Johnson, R.M.; Murdock, C.C.; Tsujimoto, H.; Suzuki, Y.; Patt, A.A.; Cui, L.; Nossa, C.W.; Barry, R.M.; et al. Native microbiome impedes vertical transmission of Wolbachia in Anopheles mosquitoes. Proc. Natl. Acad. Sci. USA 2014, 111, 12498–12503. [Google Scholar] [CrossRef] [Green Version]
- Rossi, P.; Ricci, I.; Cappelli, A.; Damiani, C.; Ulissi, U.; Mancini, M.V.; Valzano, M.; Capone, A.; Epis, S.; Crotti, E.; et al. Mutual exclusion of Asaia and Wolbachia in the reproductive organs of mosquito vectors. Parasites Vectors 2015, 8, 278. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Phylum level composition (% of OTUs) in different organs of three different populations of C. capitata. Only OTUs representing >1% of the total reads are represented. RO: reproductive organs; F: females; M: males.
Figure 1.
Phylum level composition (% of OTUs) in different organs of three different populations of C. capitata. Only OTUs representing >1% of the total reads are represented. RO: reproductive organs; F: females; M: males.

Figure 2.
Genus level composition (% of OTUs) in different organs of C. capitata. Only OTUs representing >1% of the total reads are represented. RO: reproductive organs; M: male; F: female.
Figure 2.
Genus level composition (% of OTUs) in different organs of C. capitata. Only OTUs representing >1% of the total reads are represented. RO: reproductive organs; M: male; F: female.

Figure 3.
Asaia strains from C. capitata. Asaia isolates from the (A) Guatemala strain, (B) La réunion strain and (C) ISPRA strain. Asaia strains genetically modified to express a green fluorescent protein (GFP): (D) Guatemala strain, (E) La Réunion strain and (F) ISPRA strain. Bar = 10 µm.
Figure 3.
Asaia strains from C. capitata. Asaia isolates from the (A) Guatemala strain, (B) La réunion strain and (C) ISPRA strain. Asaia strains genetically modified to express a green fluorescent protein (GFP): (D) Guatemala strain, (E) La Réunion strain and (F) ISPRA strain. Bar = 10 µm.

Figure 4.
Asaia-GFP detection in C. capitata organs. (A) Gut colonized with Asaia-GFP; in the red box, a magnification of a gut portion with green bacteria (bar 50µm). (B) Phase-contrast image, (C) Gut from C. capitata fed on a standard diet (without Asaia). (D) Phase-contrast image.
Figure 4.
Asaia-GFP detection in C. capitata organs. (A) Gut colonized with Asaia-GFP; in the red box, a magnification of a gut portion with green bacteria (bar 50µm). (B) Phase-contrast image, (C) Gut from C. capitata fed on a standard diet (without Asaia). (D) Phase-contrast image.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cappelli, A.; Petrelli, D.; Gasperi, G.; Serrao, A.G.M.; Ricci, I.; Damiani, C.; Favia, G. Bacterial Symbionts in Ceratitis capitata. Insects 2022, 13, 474. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050474
AMA Style
Cappelli A, Petrelli D, Gasperi G, Serrao AGM, Ricci I, Damiani C, Favia G. Bacterial Symbionts in Ceratitis capitata. Insects. 2022; 13(5):474. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050474
Chicago/Turabian StyleCappelli, Alessia, Dezemona Petrelli, Giuliano Gasperi, Aurelio Giuseppe Maria Serrao, Irene Ricci, Claudia Damiani, and Guido Favia. 2022. "Bacterial Symbionts in Ceratitis capitata" Insects 13, no. 5: 474. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050474
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.