
Culex Flavivirus Isolation from Naturally Infected Mosquitoes Trapped at Rio de Janeiro City, Brazil

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
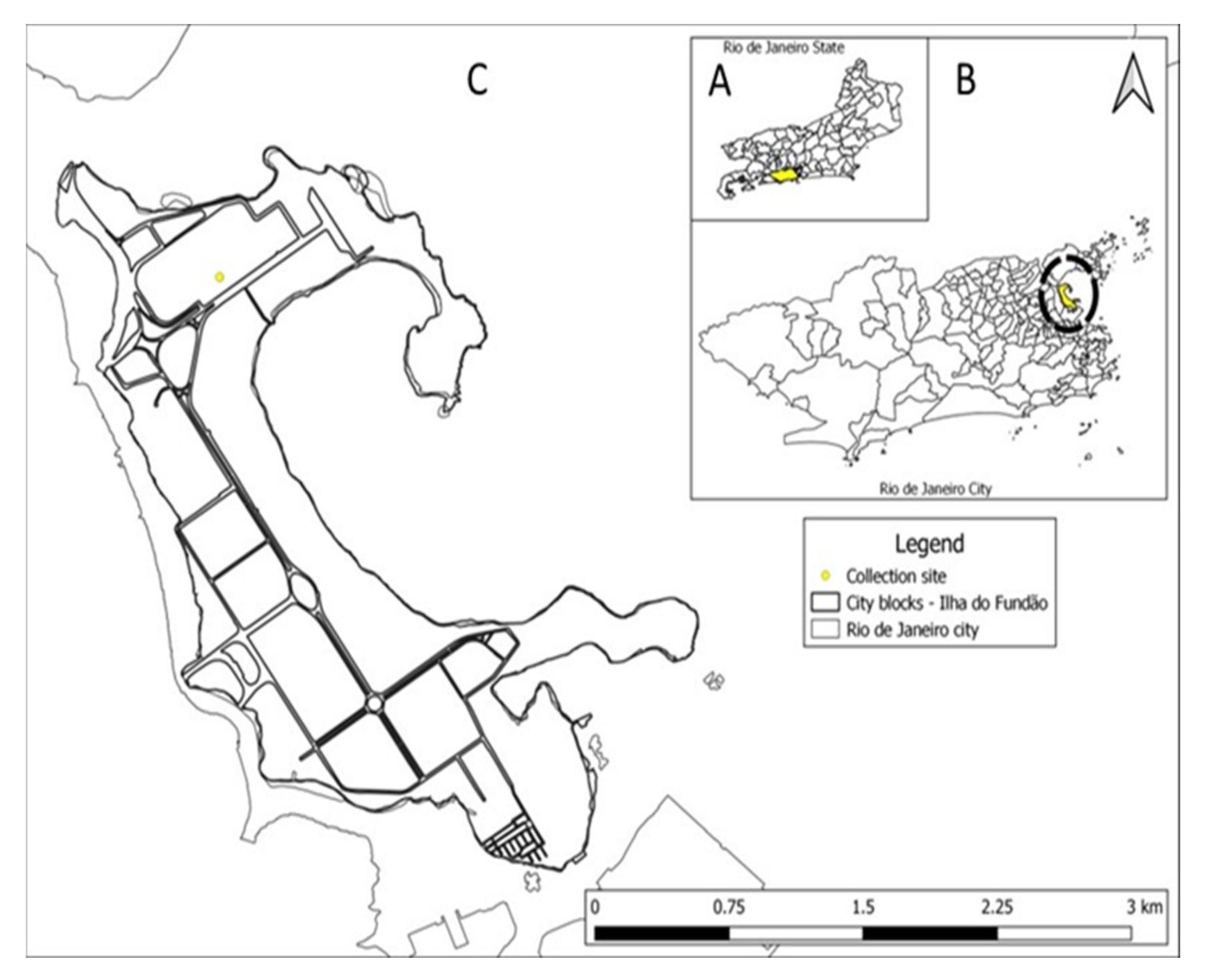
2.1. Sample Collection
2.2. CxFV Detection by qRT-PCR
2.3. Cell Culture
2.4. Virus Isolation
2.5. Plaque-Forming Units
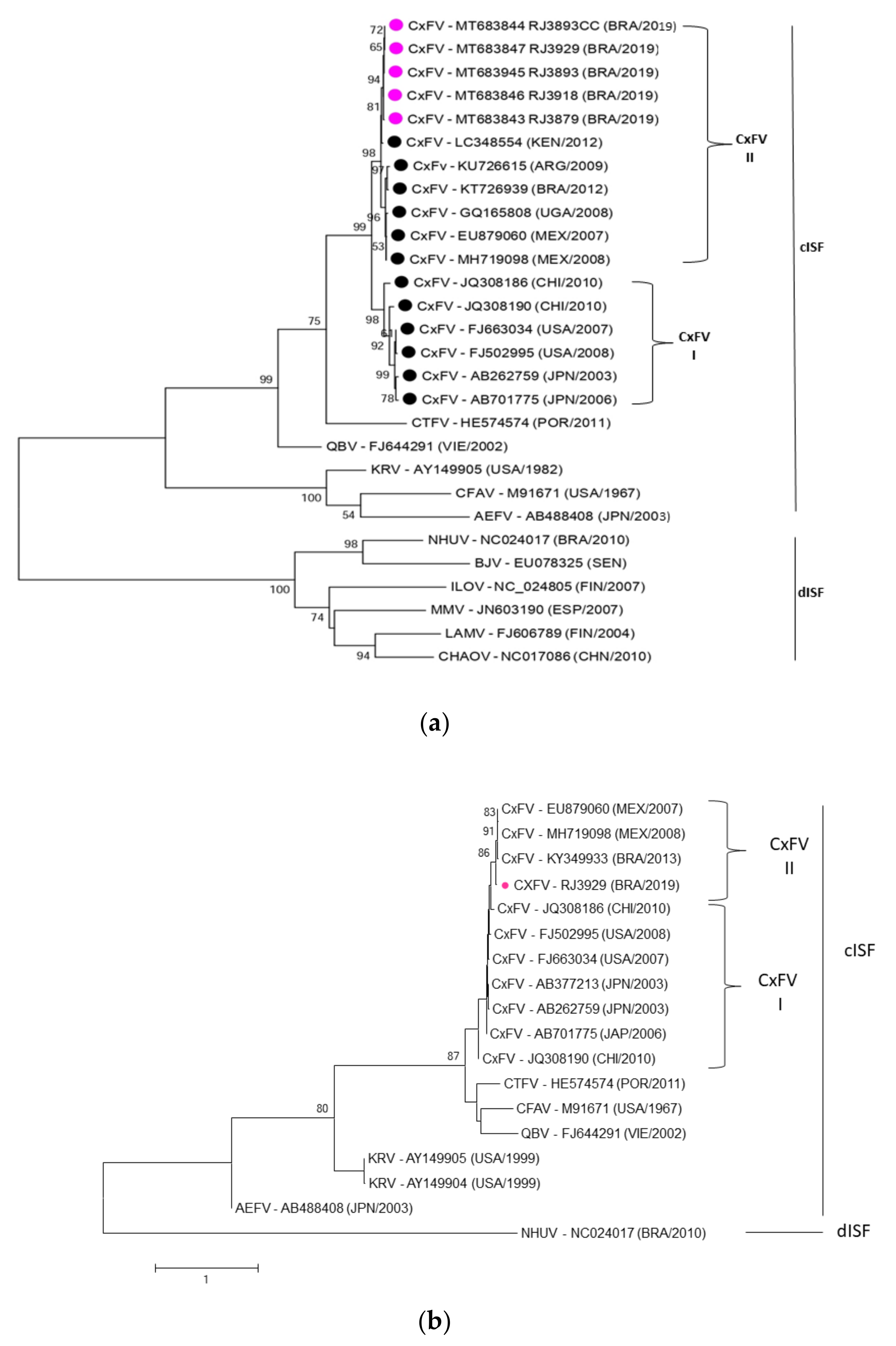
2.6. Sequencing and Phylogenetic Analysis
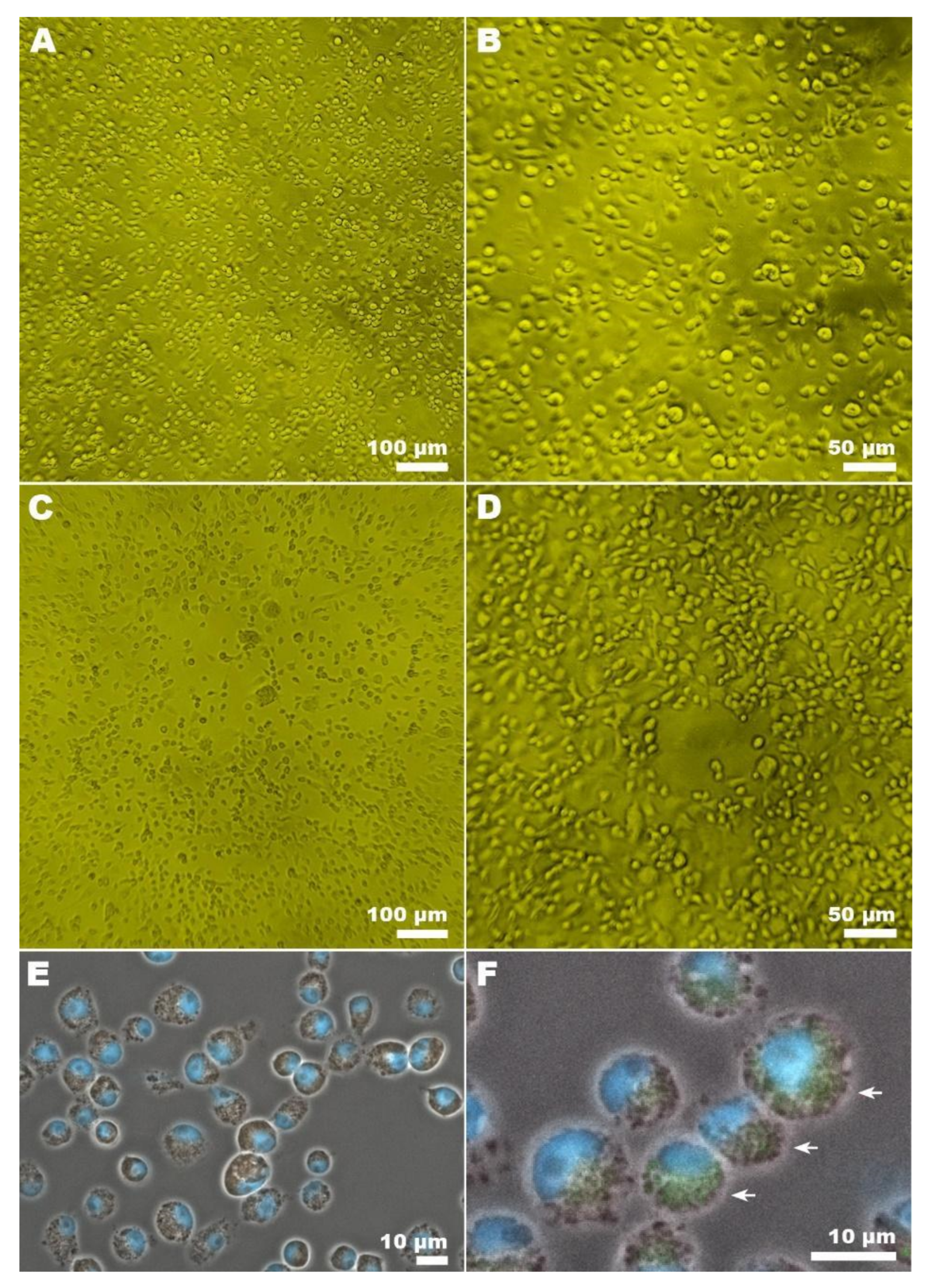
2.7. Light Microscopy of CxFV Infected Cells
2.8. Confocal Microscopy of CxFV Infected Cells
2.9. Statistical Analysis
3. Results
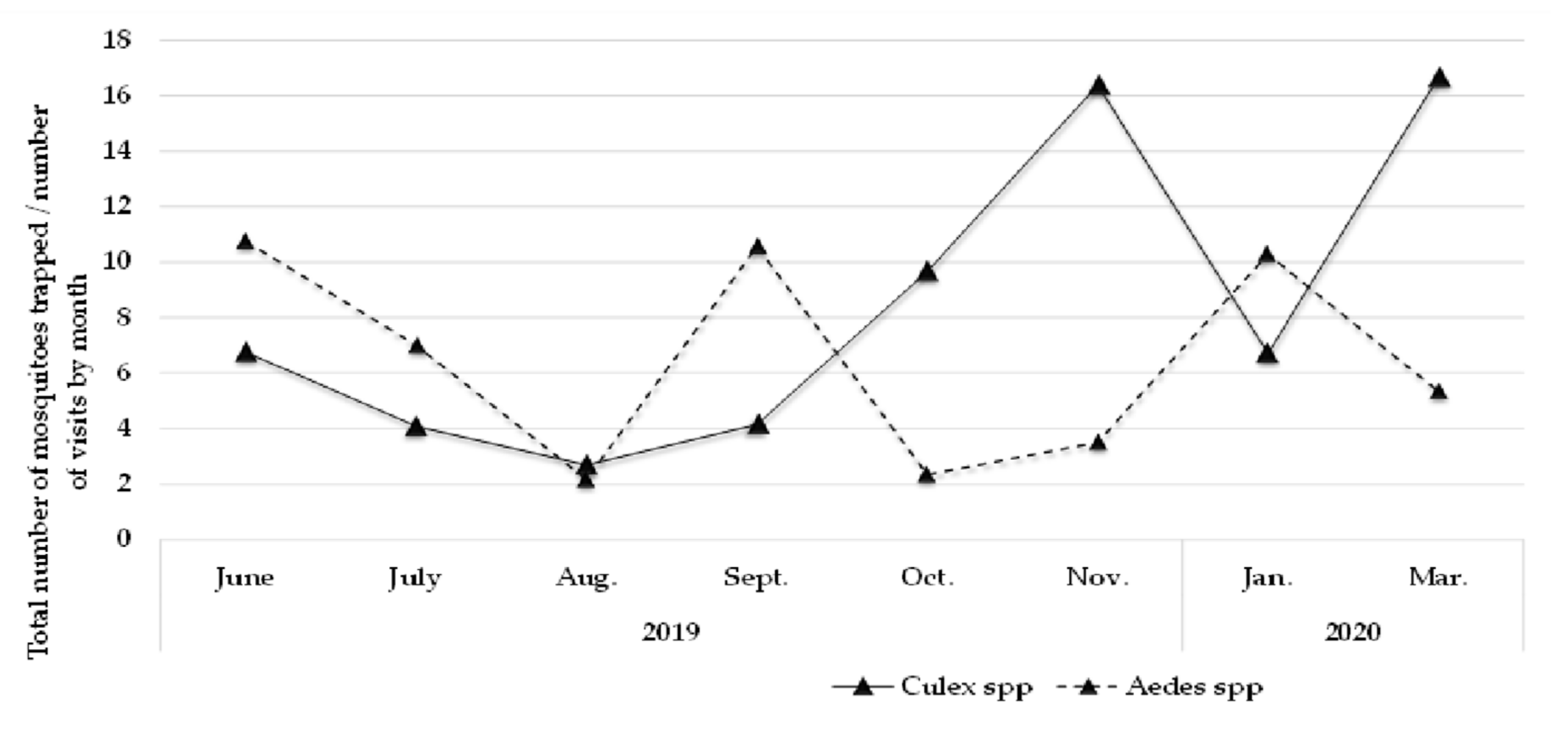
3.1. Mosquitoes Study Population
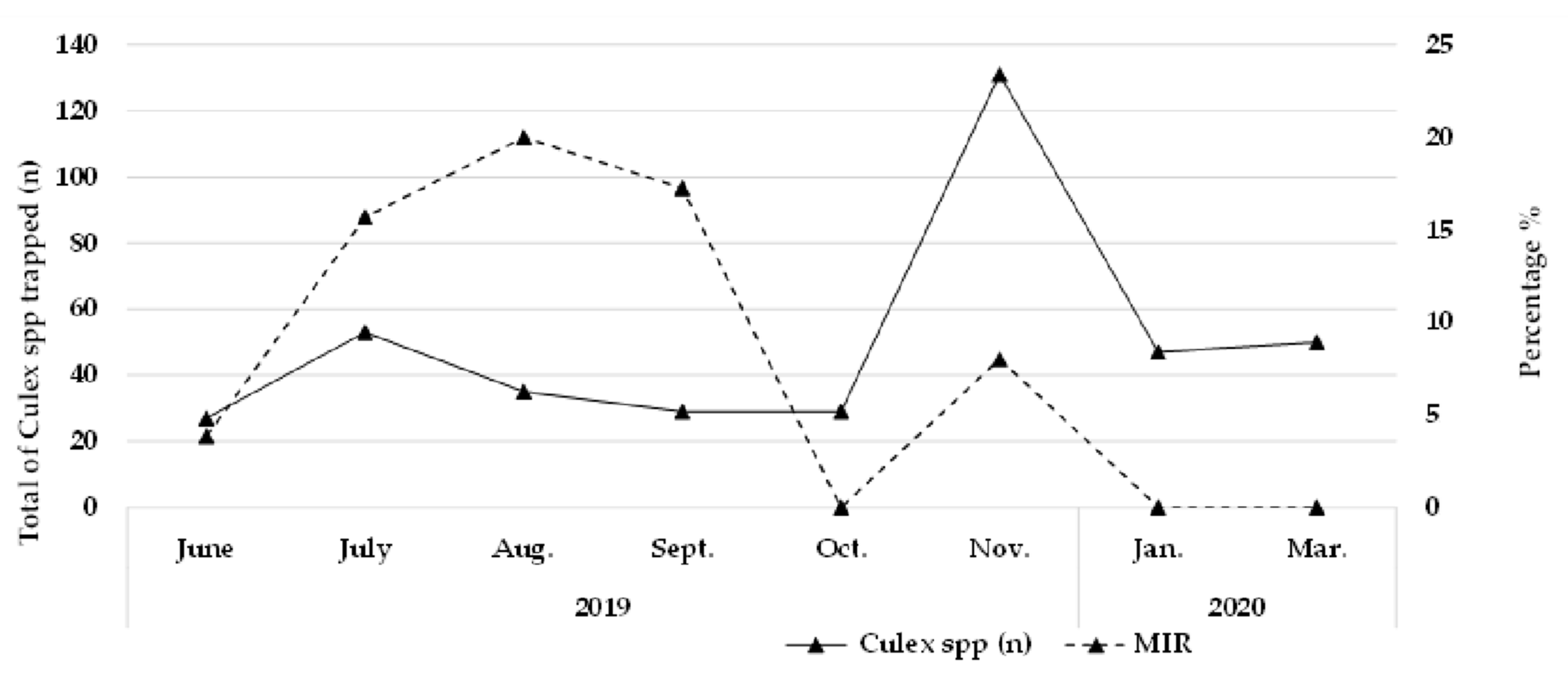
3.2. Culex Flavivirus Minimum Infection Rate (MIR)
3.3. CxFV Isolation, PFU Titration, and Microscopy
3.4. Phylogenetic Analysis of CxFV
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blitvich, B.J.; Firth, A.E. Insect-specific flaviviruses: A systematic review of their discovery, host range, mode of transmission, superinfection exclusion potential and genomic organization. Viruses 2015, 7, 1927–1959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, C.M.; Lenches, E.M.; Eddy, S.R.; Shin, S.J.; Sheets, R.L.; Strauss, J.H. Nucleotide sequence of yellow fever virus: Implications for flavivirus gene expression and evolution. Science 1985, 229, 726–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markoff, L. 5’-and 3’-noncoding regions in flavivirus RNA. Adv. Virus. Res. 2003, 59, 177–228. [Google Scholar] [CrossRef]
- Lindenbach, B.D.; Thiel, H.-J.; Rice, C.M. Flaviviridae: The viruses and their replication. In Fields Virology, 5th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2007; pp. 1101–1152. [Google Scholar]
- Newman, C.M.; Cerutti, F.; Anderson, T.K.; Hamer, G.L.; Walker, E.D.; Kitron, U.D.; Ruiz, M.O.; Brawn, J.D.; Goldberg, T.L. Culex flavivirus and West Nile virus mosquito coinfection and positive ecological association in Chicago, United States. Vector Borne Zoonotic Dis. 2011, 11, 1099–1105. [Google Scholar] [CrossRef] [Green Version]
- Hoshino, K.; Isawa, H.; Tsuda, Y.; Yano, K.; Sasaki, T.; Yuda, M.; Takasaki, T.; Kobayashi, M.; Sawabe, K. Genetic characterization of a new insect flavivirus isolated from Culex pipiens mosquito in Japan. Virology 2007, 359, 405–414. [Google Scholar] [CrossRef] [Green Version]
- Machado, D.C.; Mondini, A.; dos Santos Santana, V.; Yonamine, P.T.; Chiaravalloti, N.F.; Zanotto, P.M.; Nogueira, M.L. First identification of Culex flavivirus (Flaviviridae) in Brazil. Intervirology 2012, 55, 475–483. [Google Scholar] [CrossRef]
- Goenaga, S.; Fabbri, C.M.; García, J.B.; Rondán, J.C.; Gardenal, N.; Calderón, G.E.; Enria, D.A.; Levis, S.M. New strains of Culex flavivirus isolated in Argentina. J. Med. Entomol. 2014, 51, 900–906. [Google Scholar] [CrossRef]
- Morales-Betoulle, M.E.; Monzón Pineda, M.L.; Sosa, S.M.; Panella, N.; López, M.R.; Cordón-Rosales, C.; Komar, N.; Powers, A.; Johnson, B.W. Culex flavivirus isolates from mosquitoes in Guatemala. J. Med. Entomol. 2008, 45, 1187–1190. [Google Scholar] [CrossRef]
- Kim, D.Y.; Guzman, H.; Bueno, R., Jr.; Dennett, J.A.; Auguste, A.J.; Carrington, C.V.; Popov, V.L.; Weaver, S.C.; Beasley, D.W.; Tesh, R.B. Characterization of Culex Flavivirus (Flaviviridae) strains isolated from mosquitoes in the United States and Trinidad. Virology 2009, 386, 154–159. [Google Scholar] [CrossRef] [Green Version]
- Cook, S.; Bennett, S.N.; Holmes, E.C.; De Chesse, R.; Moureau, G.; de Lamballerie, X. Isolation of a new strain of the flavivirus cell fusing agent virus in a natural mosquito population from Puerto Rico. J. Gen. Virol. 2006, 87 Pt 4, 735–748. [Google Scholar] [CrossRef]
- Blitvich, B.J.; Lin, M.; Dorman, K.S.; Soto, V.; Hovav, E.; Tucker, B.J.; Staley, M.; Platt, K.B.; Bartholomay, L.C. Genomic sequence and phylogenetic analysis of Culex flavivirus, an insect-specific flavivirus, isolated from Culex pipiens (Diptera: Culicidae) in Iowa. J. Med. Entomol. 2009, 46, 934–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Wang, H.; Fu, S.; Liu, G.; Liu, H.; Gao, X.; Song, Z.; Rayner, S.; Xu, A.; Liang, G. Isolation and identification of a distinct strain of Culex Flavivirus from mosquitoes collected in Mainland China. Virol. J. 2012, 9, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.Y.; Lin, J.W.; Fan, Y.C.; Tu, W.C.; Chang, G.J.; Chiou, S.S. First detection of the Africa/Caribbean/Latin Americansubtype of Culex flavivirus in Asian country, Taiwan. Comp. Immunol. Microbiol. Infect Dis. 2013, 36, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Moraes, O.S.; Cardoso, B.F.; Pacheco, T.A.; Pinto, A.Z.L.; Carvalho, M.S.; Hahn, R.C.; Burlamaqui, T.C.T.; Oliveira, L.F.; Oliveira, R.S.; Vasconcelos, J.M.; et al. Natural infection by Culex flavivirus in Culex quinquefasciatus mosquitoes captured in Cuiabá, Mato Grosso Mid-Western Brazil. Med. Vet. Entomol. 2019, 33, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Araújo, J.M.; Freire, R.C.; Costa, D.M.; Sousa, D.M.; Sousa, D.M.; Monteiro, J.D.; Branco, M.S.; Farias, K.J.; Fernandes, J.V.; Gama, R.; et al. Detection of Culex flavivirus (Flaviviridae) from a natural Culex (Culex) chidesteri Dyar, 1921 population, Caatinga Biome, Semiarid Scrubland, Brazil. Entomol. Commun. 2021, 3, 1305–2675. [Google Scholar] [CrossRef]
- Gravina, H.D.; Suzukawa, A.A.; Zanluca, C.; Cardozo, S.F.M.; Tschá, M.K.; Martins da Silva, A.; Faoro, H.; da Silva Ribeiro, R.; Mendoza, T.L.P.; Rojas, A.; et al. Identification of insect-specific flaviviruses in areas of Brazil and Paraguay experiencing endemic arbovirus transmission and the description of a novel flavivirus infecting Sabethes belisarioi. Virology 2019, 527, 98–106. [Google Scholar] [CrossRef]
- Bolling, B.G.; Olea-Popelka, F.J.; Eisen, L.; Moore, C.G.; Blair, C.D. Transmission dynamics of an insect-specific flavivirus in a naturally infected Culex pipiens laboratory colony and effects of co-infection on vector competence for West Nile virus. Virology 2012, 427, 90–97. [Google Scholar] [CrossRef] [Green Version]
- Principais Mosquitos De Importância Sanitária No Brasil. Available online: http://books.scielo.org (accessed on 20 June 2019).
- Cao, Y.X.; He, X.X.; Fu, S.H.; He, Y.; Li, H.; Gao, X.Y.; Liang, G.D.; Wang, H.Y. Real-time RT-PCR Assay for the Detection of Culex flavivirus. Biomed. Environ. Sci. 2015, 28, 917–919. [Google Scholar] [CrossRef]
- Kuno, G.; Chang, G.J.; Tsuchiya, K.R.; Karabatsos, N.; Cropp, C.B. Phylogeny of the genus Flavivirus. J. Virol. 1998, 72, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Velez, J.O.; Lambert, A.J.; Johnson, A.J.; Stanfield, S.M.; Duffy, M.R. Genetic and serologic properties of Zika virus associated with an epidemic, Yap State, Micronesia, 2007. Emerg. Infect. Dis. 2008, 14, 1232–1239. [Google Scholar] [CrossRef]
- Panning, M.; Grywna, K.; van Esbroeck, M.; Emmerich, P.; Drosten, C. Chikungunya fever in travelers returning to Europe from the Indian Ocean region, 2006. Emerg. Infect. Dis. 2008, 14, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Leparc-Goffart, I.; Baragatti, M.; Temmam, S.; Tuiskunen, A.; Moureau, G.; Charrel, R.; de Lamballerie, X. Development and validation of real-time one-step reverse transcription-PCR for the detection and typing of dengue viruses. J. Clin. Virol. 2009, 45, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Fischer, C.; Torres, M.C.; Patel, P.; Moreira-Soto, A.; Gould, E.A.; Charrel, R.N.; de Lamballerie, X.; Nogueira, R.M.R.; Sequeira, P.C.; Rodrigues, C.D.S.; et al. Lineage-Specific Real-Time RT-PCR for Yellow Fever Virus Outbreak Surveillance, Brazil. Emerg. Infect. Dis. 2017, 23, 1867–1871. [Google Scholar] [CrossRef] [PubMed]
- Machado, D.C. Sequenciamento do Genoma Completo do vírus Culex Flavivirus (Flaviviridae) Isolado no Brasil. Master’s Thesis, Universidade Estadual Paulista Julio de Mesquita Filho, Instituto de Biociências, Letras e Ciências Exatas, São Paulo, Brazil, 2016. Available online: http://hdl.handle.net/11449/154714 (accessed on 18 May 2022).
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis programfor Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Kumar, S.; Glen, S. MEGA 7: Molecular Evolutionary Genetics Analysis (MEGA) Software Version 7.0.2. Mol. Biol. Evolut. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.r-project.org/ (accessed on 8 May 2021).
- RStudio Team. RStudio: Integrated Development Environment for R; RStudio: Boston, MA, USA, 2021; Available online: http://www.rstudio.com/ (accessed on 8 May 2021).
- Degener, C.M.; Geier, M.; Kline, D.; Urban, J.; Willis, S.; Ramirez, K.; Cloherty, E.R.; Gordon, S.W. Field trials to evaluate the effectiveness of the biogents sweetscent lure in combination with several commercial mosquito traps and to assess the effectiveness of the biogents mosquitaire trap with and without carbon dioxide. J. Am. Mosq. Control Assoc. JAMCA 2019, 35, 32–391. [Google Scholar] [CrossRef] [Green Version]
- Unlu, I.; Farajollahi, A. A multiyear surveillance for Aedes albopictus with Biogents Sentinel trap counts for males and species composition of othermosquito species. J. Am. Mosq. Control. Assoc. 2014, 30, 122–125. [Google Scholar] [CrossRef]
- Maciel-de-Freitas, R.; Eiras, E.A.; Lourenço-de-Oliveira, R. Field evaluation of effectiveness of the BG-Sentinel, a new trap for capturing adult Aedes aegypti (Diptera: Culicidae). Mem. Inst. Oswaldo Cruz. 2006, 101, 321–325. [Google Scholar] [CrossRef] [Green Version]
- Haddow, A.D.; Guzman, H.; Popov, V.L.; Wood, T.G.; Widen, S.G.; Haddow, A.D.; Tesh, R.B.; Weaver, S.C. First isolation of Aedes flavivirus in the Western Hemisphere and evidence of vertical transmission in the mosquito Aedes (Stegomyia) albopictus (Diptera: Culicidae). Virology 2013, 440, 134–139. [Google Scholar] [CrossRef] [Green Version]
- Bittar, C.; Machado, D.C.; Vedovello, D.; Ullmann, L.S.; Rahal, P.; Araújo Junior, J.P.; Nogueira, M.L. Genome sequencing and genetic characterization of Culex Flavirirus (CxFV) provides new information about its genotypes. Virol. J. 2016, 13, 158. [Google Scholar] [CrossRef] [Green Version]
- Liang, W.; He, X.; Liu, G.; Zhang, S.; Fu, S.; Wang, M.; Chen, W.; He, Y.; Tao, X.; Jiang, H.; et al. Distribution and phylogenetic analysis of Culex flavivirus in mosquitoes in China. Arch. Virol. 2015, 160, 2259–2268. [Google Scholar] [CrossRef] [PubMed]
- Kent, R.J.; Crabtree, M.B.; Miller, B.R. Transmission of West Nile Virus by Culex quinquefasciatus Say Infected with Culex Flavivirus Izabal. PLoS Negl. Trop. Dis. 2010, 4, e671. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specie | Sex | n/pool | Mean | Median | Min. | Max. | Standard Deviation | Percent |
|---|---|---|---|---|---|---|---|---|
| Aedes spp. | Female | 209/69 | 3.60 | 2 | 0 | 18 | 4.43 | 27.50 |
| Female-eng 1 | 4 | 0.069 | 0 | 0 | 2 | 0.32 | 0.53 | |
| Male | 146/49 | 2.53 | 1 | 0 | 19 | 3.96 | 19.21 | |
| Culex spp. | Female | 135/50 | 2.33 | 1 | 0 | 10 | 2.74 | 17.76 |
| Female-eng 1 | 12 | 0.21 | 0 | 0 | 2 | 0.52 | 1.58 | |
| Male | 254/72 | 4.38 | 3 | 0 | 15 | 3.87 | 33.42 |
| Year | Month | Males n | Positive pools | Females n | Positive pools | MIR * |
|---|---|---|---|---|---|---|
| 2019 | June | 16 | 1 | 11 | 1 | 3.7037 |
| July | 39 | 7 | 14 | 1 | 15.0943 | |
| Aug. | 27 | 6 | 8 | 1 | 20 | |
| Sept. | 21 | 2 | 8 | 3 | 17.2414 | |
| Oct. | 12 | 0 | 17 | 0 | 0.0000 | |
| Nov. | 79 | 6 | 52 | 4 | 7.63359 | |
| 2020 | Jan. | 28 | 0 | 19 | 0 | 0.000000 |
| Mar. | 32 | 0 | 18 | 0 | 0.000000 | |
| Total | 254 | 22 | 147 | 9 | 7,730,673 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amaral, C.; Câmara, D.; Salles, T.; Meneses, M.D.; Araújo-Silva, C.d.; Dias, V.; Costa, F.d.; Caldas, L.; Azevedo, R. Culex Flavivirus Isolation from Naturally Infected Mosquitoes Trapped at Rio de Janeiro City, Brazil. Insects 2022, 13, 477. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050477
Amaral C, Câmara D, Salles T, Meneses MD, Araújo-Silva Cd, Dias V, Costa Fd, Caldas L, Azevedo R. Culex Flavivirus Isolation from Naturally Infected Mosquitoes Trapped at Rio de Janeiro City, Brazil. Insects. 2022; 13(5):477. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050477
Chicago/Turabian StyleAmaral, Cinthya, Daniel Câmara, Tiago Salles, Marcelo Damião Meneses, Carlla de Araújo-Silva, Vanessa Dias, Fábio da Costa, Lúcio Caldas, and Renata Azevedo. 2022. "Culex Flavivirus Isolation from Naturally Infected Mosquitoes Trapped at Rio de Janeiro City, Brazil" Insects 13, no. 5: 477. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050477