
Two New Phoretic Species of Heterostigmatic Mites (Acari: Prostigmata: Neopygmephoridae and Scutacaridae) on Australian Hydrophilid Beetles (Coleoptera: Hydrophilidae) †
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
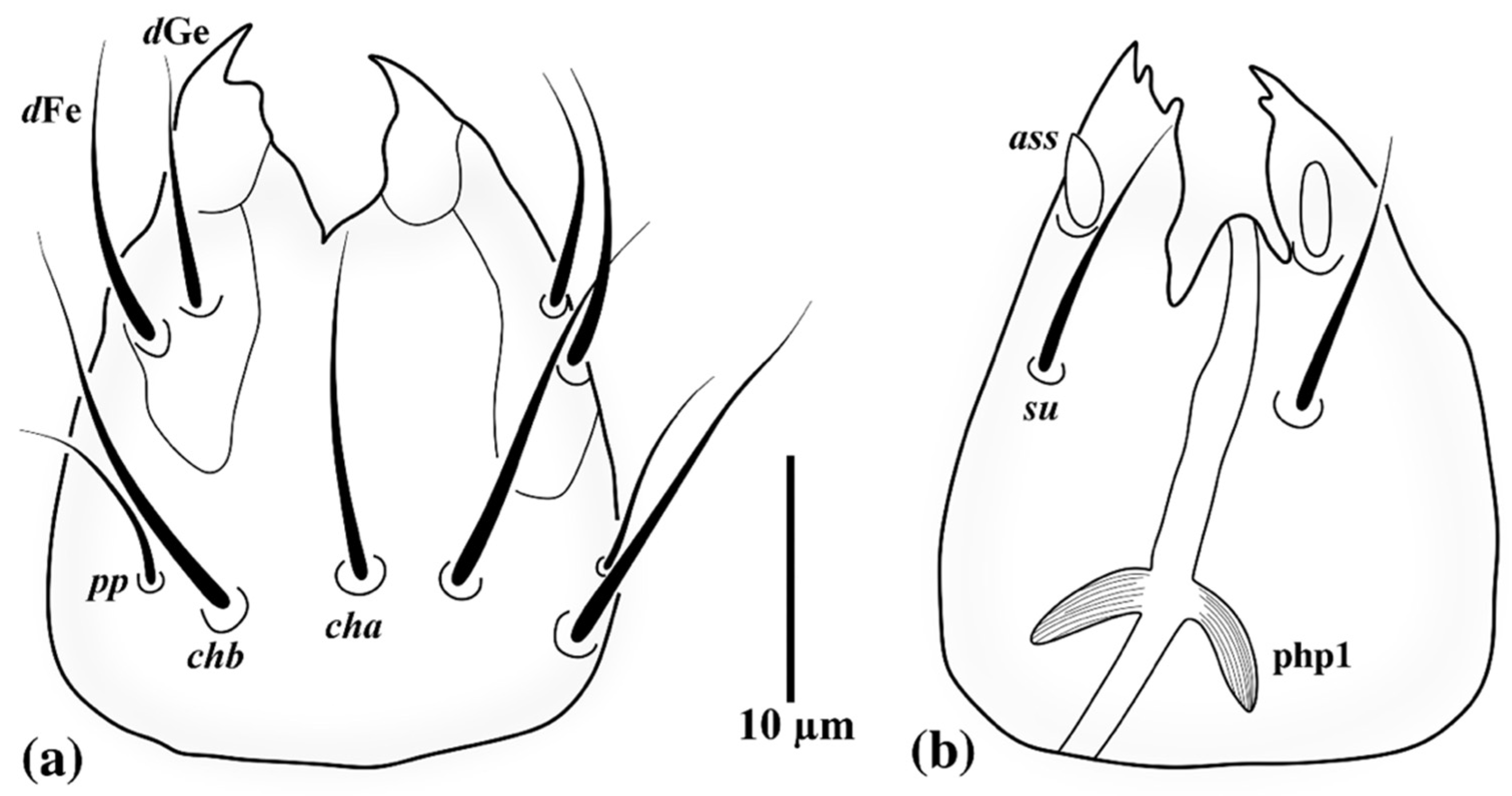
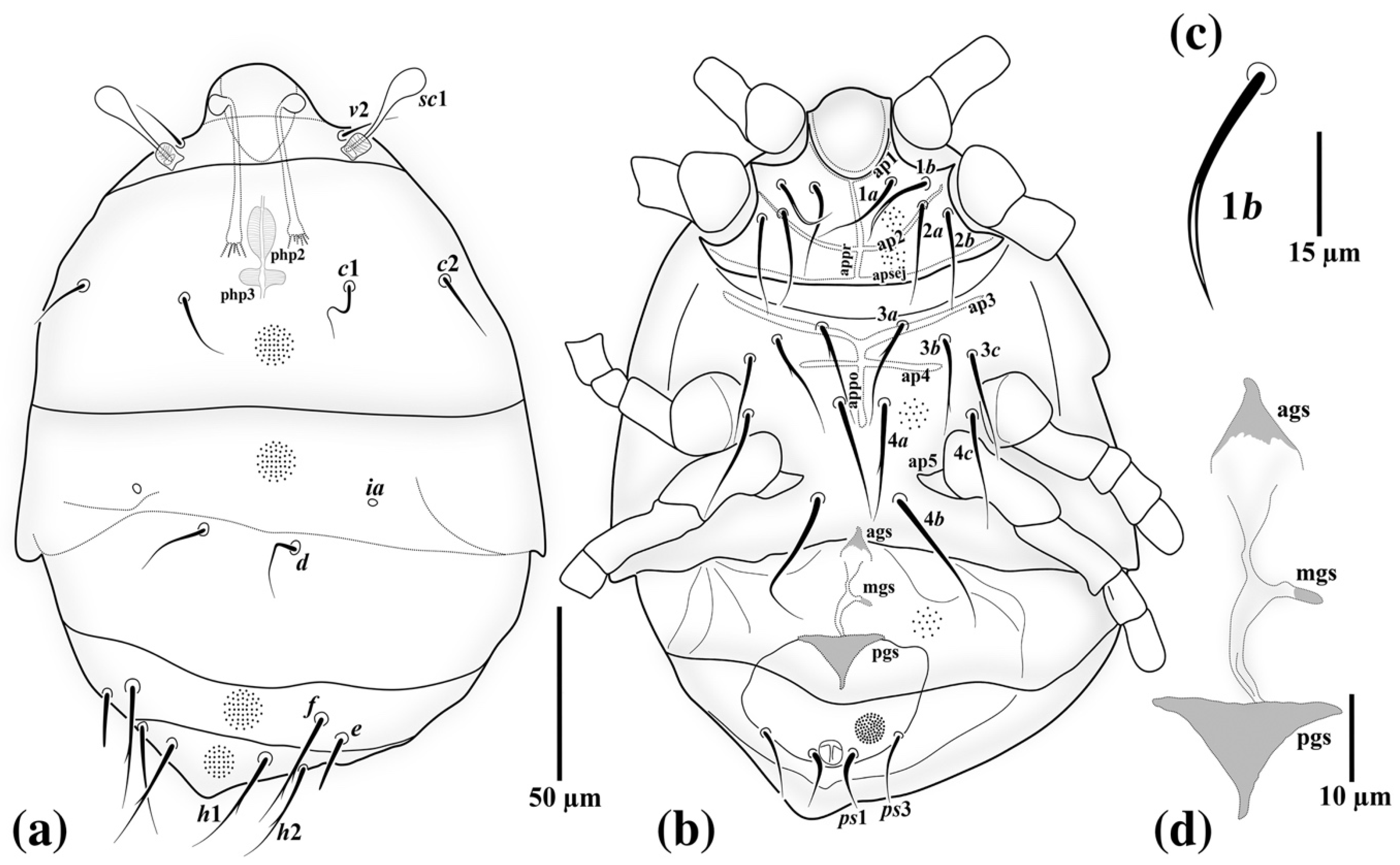
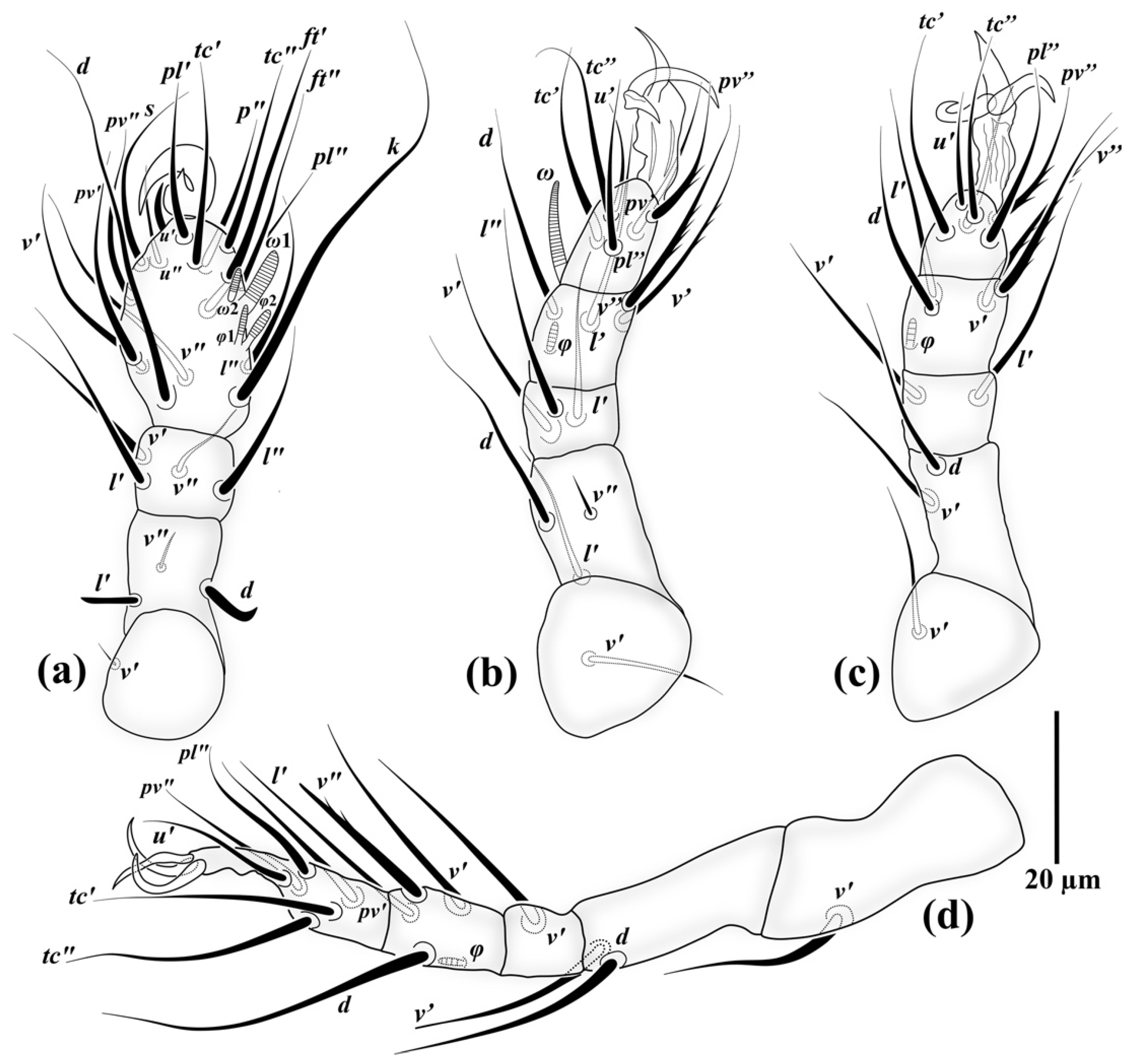
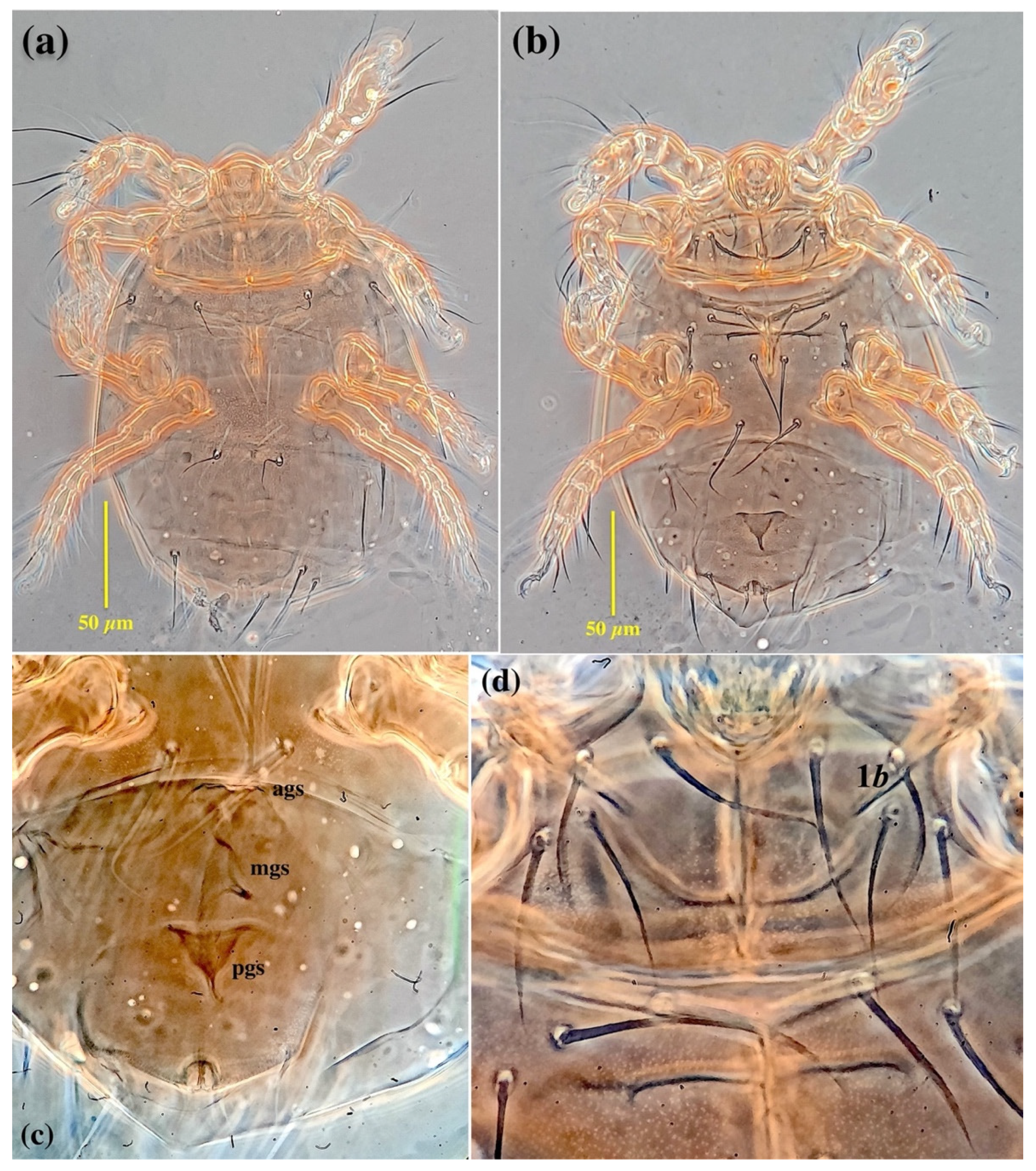
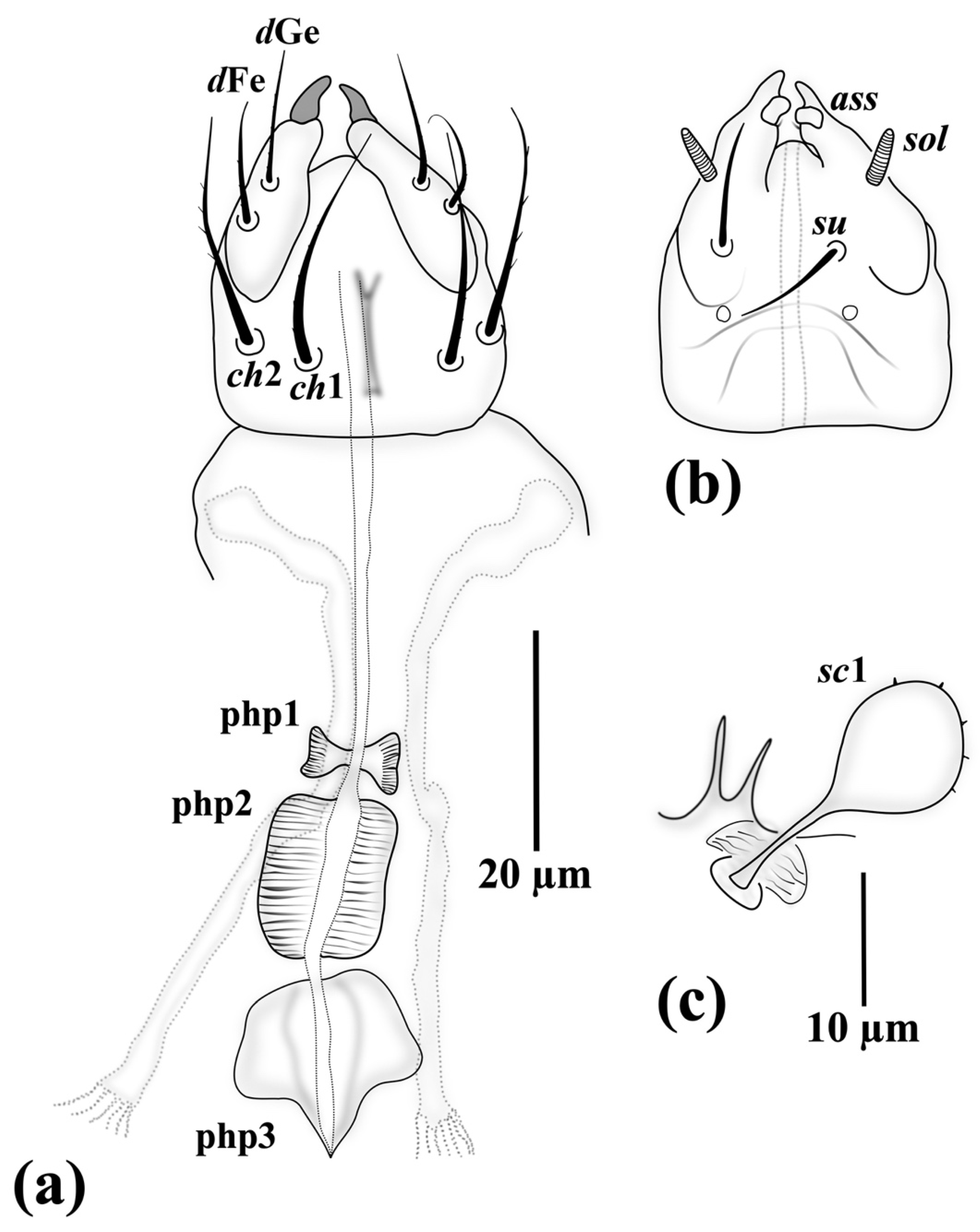
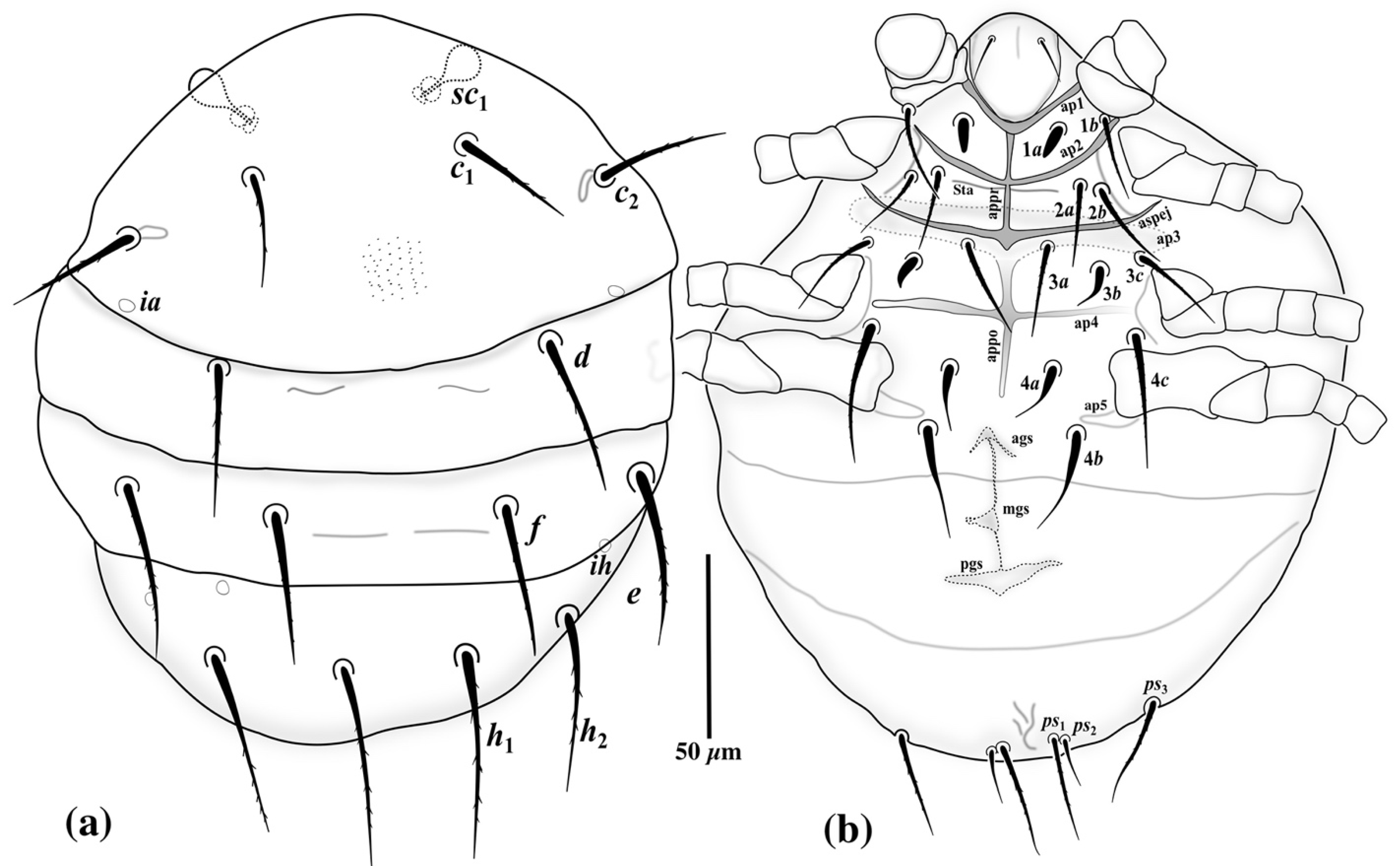
3.1. Family Neopygmephoridae Cross, 1965
3.1.1. Differential Diagnosis
3.1.2. Etymology
- Key to the world species of Allopygmephorus
- 1.
- With three solenidia on tibiotarsus I .......................................................................................................................................................................................... 2
- -
- With four solenidia on tibiotarsus I ............................................................................................................................................................................................ 3
- 2.
- Seta e subequal to setae h1 and h2, pgs absence ........................................................................................................ A. nanhuiensis Gao, Zou and Ma, 1989
- -
- Seta e about half the length of h1 and h2, pgs presence ............................................................................................................... A. chinensis Mahunka 1975
- 3.
- Seta 1a, modified, with very long barbs; Setae f, h1 and h2 blunt-ended .......................................................................................................................................................................................... A. bakaninae Khaustov and Ermilov, 2008
- -
- Seta 1a normality; Setae f, h1 and h2 pointed or blunt-ended ...................................................................................................................................................................................... 4
- 4.
- Tibiotarsus I with a weak-stalked small claw ............................................................................................................................................................................... 5
- -
- Tibiotarsus I with a normal or large claw .................................................................................................................................................................................... 6
- 5.
- Solenidion ω2 on tibiotarsus I long, approximately more than half the length of segment; ω2 > ω1, φ1 ≈ φ2 ........................ A. cunae Mahunka, 1970
- -
- Solenidion ω2 on tibiotarsus I shorter than half the length of segment; ω2 ≈ ω1, φ1 > φ2 .......................................................... A. brasiliensis Mahunka, 1970
- 6.
- Setae c1, c2 and d basally thickened, bulbiform; ps2 absence .................................................................... A. orientalis Mahunka and Mahunka-Papp, 1988
- -
- Setae c1, c2 and d not basally thickened, normality; ps2 absence or presence ......................................................................................................................... 7
- 7.
- Seta c1 considerably shorter than c2; 4b long, arising end of body; three pairs of ps; h1 and h2 blunt-ended, weakly barbed ............................................................................................. A. tuberosus Mahunka, 1969
- -
- Seta c1 = c2 or c1 somewhat shorter than c2; 4b not arising end of body; three or two pairs of ps; h1 and h2 blunt-ended or pointed ...................................................................................................................................................................................................................................... 8
- 8.
- Seta 1b modified, with a notch or slit in the distal half; ags, mgs and pgs presence ................................................................. A. coelostomus sp. nov.
- -
- Seta 1b without big notch or slit, mgs absence, ags and pgs presence or absence .......................................................................................................................... 9
- 9.
- Seta 1b thickened and lanceolate; seta h2 blunt-ended, barbed ....................................................................................................................................... 10
- -
- Seta 1b normality, barbed or smooth; seta h2 blunt-ended or pointed ............................................................................................................................... 11
- 10.
- Seta h1 blunt-ended; seta v″ on femur II spiniform .......................................................................... A. spinisetus Khaustov and Sazhnev, 2016
- -
- Seta h1 pointed; seta v″ on femur II unmodified ........................................................................................ A. punctatus Khaustov and Sazhnev, 2016
- 11.
- Seta h1 pointed; h1 and h2 subequal .......................................................................................... A. persicus Khaustov and Hajiqanbar, 2006
- -
- Seta h1 blunt-ended, h1 and h2 subequal or h1 > h2 .......................................................................................................................................................... 12
- 12.
- Setae h1 and h2 subequal; solenidia ω2 > φ1 ≈ φ2 > ω1 on tibiotarsus I ........................................................ ... A. baoshanensis Gao, Zou and Ma, 1989
- -
- Setae h1 > h2; solenidia otherwise ......................................................................................................................................................................................... 13
- 13.
- Solenidia ω1 > φ1 > φ2 > ω2 on tibiotarsus I; seta e shorter than half of the seta f........................................................................................................................................................................................................ A. matthesi Krczal, 1959
- -
- Solenidia ω1 > ω2 ≈ φ2 > φ1 on tibiotarsus I; seta e about half the length of seta f .............................................................. A. heterodactylus Mahunka, 1973
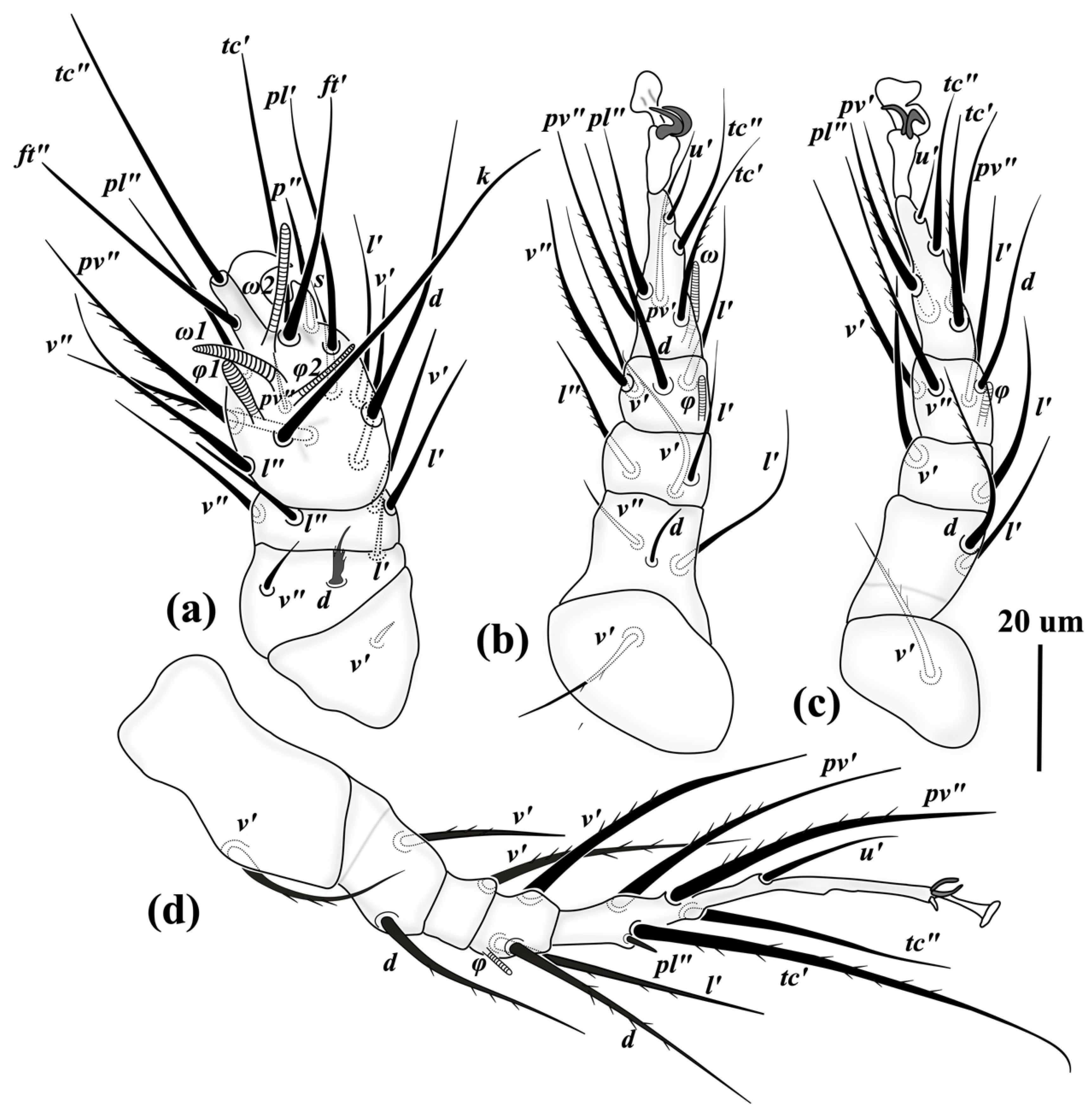
3.2. Family Scutacaridae Oudemans, 1916
3.2.1. Differential Diagnosis
3.2.2. Etymology
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaliszewski, M.; Athias-Binche, F.; Lindquist, E.E. Parasitism and parasitoidism in Tarsonemina (Acari: Heterostigmata) and evolutionary considerations. Adv. Parasitol. 1995, 35, 335–367. [Google Scholar] [PubMed]
- Walter, D.E.; Lindquist, E.E.; Smith, I.M.; Cook, D.R.; Krantz, G.W. Order Trombidiformes. In A Manual of Acarology, 3rd ed.; Krantz, G.W., Walter, D.E., Eds.; Texas Tech University Press: Lubbock, TX, USA, 2009; pp. 233–420. [Google Scholar]
- Baumann, J.; Ferragut, F.; Simic, S. Lazy hitchhikers? Preliminary evidence for within-habitat phoresy in pygmephoroid mites (Acari, Scutacaridae). Soil Org. 2018, 90, 95–99. [Google Scholar]
- Katlav, A.; Hajiqanbar, H.; Riegler, M.; Seeman, O.D. Sheltered life beneath elytra: Three new species of Eutarsopolipus (Acari, Heterostigmatina, Podapolipidae) parasitizing Australian ground beetles. Parasite 2021, 28, 75. [Google Scholar] [CrossRef] [PubMed]
- Rahiminejad, V.; Hajiqanbar, H.; Talebi, A.A. An alpha diversity survey of heterostigmatic mites (Trombidiformes, Prostigmata) phoretic on scarabaeoid beetles in Hyrcani forest, northern Iran. Syst. Appl. Acarol. 2020, 25, 2033–2046. [Google Scholar]
- Baumann, J. Patterns of intraspecific morphological variability in soil mites reflect their dispersal ability. Exp. Appl. Acarol. 2021, 83, 241–255. [Google Scholar] [CrossRef]
- Zhang, Z.; Fan, Q.H.; Pesic, V.; Smit, H.; Bochkov, A.V.; Khaustov, A.A.; Baker, A.; Wohltmann, A.; Wen, T.; Amrine, J.W. Order Trombidiformes Reuter, 1909. In Animal Biodiversity: An Outline of Higher-Level Classification and Survey of Taxonomic Richness; Zhang, Z.-Q., Ed.; Magnolia Press: Waco, TX, USA, 2011; Volume 3148, pp. 129–138. [Google Scholar]
- Khaustov, A.A. Myrmecophilous pygmephoroid mites (Acari: Pygmephoroidea) associated with Lasius fuliginosus (Hymenoptera: Formicidae) in Western Siberia, Russia. Int. J. Acarol. 2016, 42, 92–105. [Google Scholar] [CrossRef]
- Khaustov, A.A. Mites of the Family Scutacaridae of Eastern Palaearctic; Akademperiodyka: Kiev, Ukraine, 2008; 291p. [Google Scholar]
- Hajiqanbar, H.; Khaustov, A.A.; Kamali, K.; Saboori, A.; Kamali, H. New taxa of the family Trochometridiidae (Acari: Heterostigmata) associated with insects from Iran. J. Nat. Hist. 2009, 43, 2701–2722. [Google Scholar] [CrossRef]
- Khaustov, A.A.; Trach, V.A. A new species and new records of mites of the genus Spatulaphorus Rack (Acari: Heterostigmata: Pygmephoridae) from Ukraine. Int. J. Acarol. 2012, 38, 480–485. [Google Scholar] [CrossRef]
- Katlav, A.; Hajiqanbar, H.; Talebi, A.A. Pseudopygmephorellus mazandaranicus sp. nov. (Acari: Heterostigmata: Pygmephoridae), phoretic on scarabaeid dung beetles (Coleoptera: Scarabaeidae) from Iran. Zootaxa 2015, 3919, 100–110. [Google Scholar] [CrossRef]
- Khaustov, A.A.; John, O., Jr. Two new genera and two new species of the mite family Neopygmephoridae (Acari: Heterostigmata) associated with small mammals from USA. Acarologia 2019, 59, 308–322. [Google Scholar] [CrossRef]
- Rack, G. Brennandania scolopendrae sp. n. von Scolopendra morsitans L. (Acarina, Tarsonemida, Pygmephoroidea, Microdispidae). Rev. Zool. Afr. 1979, 93, 376–381. [Google Scholar]
- Khaustov, A.A.; Minor, M.A. New taxa of the mite family Neopygmephoridae (Acari: Heterostigmata) from alpine New Zealand. Zootaxa 2018, 4415, 276–296. [Google Scholar] [CrossRef] [PubMed]
- Khaustov, A.A.; Oconnor, M.A. Three new species of Rhinopygmephorus (Acari: Heterostigmata: Neopygmephoridae) associated with halictid bees (Hymenoptera: Halictidae). Int. J. Acarol. 2019, 45, 438–449. [Google Scholar] [CrossRef]
- Eickwort, G.C. Associations of mites with social insects. Annu. Rev. Entomol. 1990, 35, 469–488. [Google Scholar] [CrossRef]
- Hajiqanbar, H.; Khaustov, A.A. New species and record of the genus Petalomium (Acari: Heterostigmatina: Neopygmephoridae) associated with ants (Hymenoptera: Formicoidea) from Iran. Biologia 2013, 68, 712–719. [Google Scholar] [CrossRef]
- Khaustov, A.A. A new species of mites of the genus Pseudopygmephorus (Acari: Heterostigmata: Neopygmephoridae) associated with Aphodius fimetarius (Coleoptera: Scarabaeidae) from Crimea. Acarina 2010, 18, 261–264. [Google Scholar]
- Khaustov, A.A. A new species and a new record of mite genus Coleopygmephorus (Acari: Neopygmephoridae) associated with bark beetles (Coleoptera: Curculionidae: Scolytinae) from Russia. Int. J. Acarol. 2017, 43, 351–358. [Google Scholar] [CrossRef]
- Sobhi, M.; Hajiqanbar, H.; Mortazavi, A. New species and records of heterostigmatic mites (Acari: Prostigmata: Heterostigmata) phoretic on scarabaeid dung beetles (Coleoptera: Scarabaeidae) from northwestern Iran. Zootaxa 2017, 4276, 427–434. [Google Scholar] [CrossRef]
- Walter, D.E.; Proctor, H.C. Mites: Ecology, Evolution and Behavior; University of New South Wales Press: Sydney, Australia, 1999. [Google Scholar]
- Ebermann, E. Scutacarus acarorum (Goeze, 1780); Heterostigmata, Scutacaridae—An example for the interrelationship between phoresy and polymorphism in mites. The Acari; Proceedings of the 2nd Symposium EURAAC, 1992; 193–196. [Google Scholar]
- Baumann, J. Tiny mites on a great journey–A review on scutacarid mites as phoronts and inquilines (Heterostigmatina, Pygmephoroidea, Scutacaridae). Acarologia 2018, 58, 192–251. [Google Scholar] [CrossRef]
- Alekseev, V.I. Dytiscidae, Noteridae, Hali-plidae, Hydraenidae, Hydrophilidae, Hydrochidae, Helophoridae, Spercheidae and Dryopidae (Coleoptera) of the Russian part of the Curonian Spit: An attempt of complex faunal analysis. Zool. Ecol. 2012, 22, 57–63. [Google Scholar] [CrossRef]
- Karafiat, H. Systematik und Ökologie der Scutacariden. In Beiträge zur Systematik und Ökologie mitteleuropaischer Acarina; Stammer, H.J., Ed.; Akademische Verlagsgesellschaft Geest & Portig: Leipzig, Germany, 1959; Volume 1, pp. 627–712. [Google Scholar]
- Kurosa, K. The Scutacarid mites of Japan. VII. Revised definition of the genera Archidispus and Imparipes, with description of five new Archidispus. Annot. Zool. Jpn. 1983, 56, 315–332. [Google Scholar]
- Ebermann, E. Das Phänomen Polymorphismus in der Milbenfamilie Scutacaridae (Acari, Heterostigmata, Tarsonemina, Scutacaridae). Zoologia 1991, 141, 1–76. [Google Scholar]
- Khaustov, A.A. Two new species of mites of the genus Archidispus (acarina: Heterostigmata: Scutacaridae) from Byelorussia and Crimea. Acarina 2002, 10, 155–159. [Google Scholar]
- Kurosa, K. The scutacarid mites of Japan. XIV. A new species of Archidispus phoretic on stenolophine carabid beetles. J. Acarol. Soc. Jpn. 2009, 18, 85–90. [Google Scholar] [CrossRef] [Green Version]
- Katlav, A.; Riegler, M.; Seeman, O.D. Tiny hitchhikers and parasites: A review of Australian heterostigmatic mites (Acari: Prostigmata) associated with insects, with description of three new species. Austral Entomol. 2020, 59, 401–421. [Google Scholar] [CrossRef]
- Seeman, O.D. Contrasting species diversification of Eutarsopolipus (Acariformes: Podapolipidae) on Castelnaudia and Notonomus (Coleoptera: Carabidae). Zootaxa 2021, 4971, 1–74. [Google Scholar] [CrossRef]
- Mahunka, S.; Philips, J.R. Tarsonemid mites associated with birds in the USA (Acarina: Tarsonemida). Folia Entomol. Hung. 1978, 31, 191–200. [Google Scholar]
- Philips, J.R. The mite (Acarina) fauna of trogid beetles (Coleoptera: Trogidae). Int. J. Acarol. 2009, 36, 1–17. [Google Scholar] [CrossRef]
- Lindquist, E.E. The world genera of Tarsonemidae (Acari: Heterostigmata): A morphological, phylogenetic, and systematic revision, with reclassification of family-group taxa in the Heterostigmata. Mem. Entomol. Soc. Can. 1986, 136, 1–517. [Google Scholar] [CrossRef]
- Khaustov, A.A.; Hajiqanbar, H. Redefinition of the genus Allopygmephorus (Acari: Heterostigmata: Neopygmephoridae), with description of a new species from Iran. Acarina 2006, 14, 191–194. [Google Scholar]
- Khaustov, A.A.; Ermilov, S.G. Two new species of mites of the superfamily Pygmephoroidea (Acari: Heterostigmata: Pygmephoridae, Neopygmephoridae) from the European part of Russia. Acarina 2008, 16, 39–43. [Google Scholar]
- Gao, J.; Zou, P.; Ma, E. Two new species of Allopygmephorus associated with commercial mushroom from China (Acari: Pygmephoridae). Acta Zootaxonomica Sin. 1989, 14, 453–456. (In Chinese) [Google Scholar]
- Mahunka, S. The scientific results of the Hungarian soil zoological expedition to South America. 21. Acari: Tarsonemine species from Brasil. Acta Zool. Acad. Sci. Hung. 1970, 16, 371–408. [Google Scholar]
- Mahunka, S. Neue und interessante Milben aus dem Genfer Museum XIV. Tarsonemida aus Hong Kong. Arch. Sci. 1975, 28, 183–188. [Google Scholar]
- Mahunka, S. Auf Insekten lebende Milben (Acari: Acarida, Tarsonemida) aus Afrika. II. Acta Zool. Acad. Sci. Hung. 1973, 19, 289–337. [Google Scholar]
- Krczal, H. Systematik und Ökologie der Pyemotiden. In Beiträge zur Systematik und Ökologie mitteleuropaischer Acarina; Stammer, H.J., Ed.; Akademische Verlagsgesellschaft Geest & Portig: Leipzig, Germany, 1959; pp. 385–625. [Google Scholar]
- Rahiminejad, V.; Yazdanian, M.; Zeitounli, S. New records of heterostigmatic mites (Acari: Heterostimatina) from dry rice cultivation of Gorgan, northern Iran. J. Insect Biodivers. Syst. 2022, 8, 121–129. [Google Scholar] [CrossRef]
- Mahunka, S.; Mahunka-Papp, L. Neue und interessante Milben aus dem Genfer Museum. LIX. Hauseripes hungarorum gen. n., sp. n. and some other new Tarsonemina (Acari) from Sabah (East Malaysia). Rev. Suisse Zooogiel 1988, 95, 581–594. [Google Scholar] [CrossRef]
- Katlav, A.; Hajiqanbar, H.; Talebi, A.A. A contribution to the knowledge of heterostigmatic mites (Acari: Prostigmata) in western Mazandaran Province, Northern Iran. Acarologia 2015, 55, 311–320. [Google Scholar] [CrossRef]
- Khaustov, A.A.; Sazhnev, A.S. Mites of the families Neopygmephoridae and Scutacaridae associated with variegated mud-loving beetles (Coleoptera: Heteroceridae) from Russia and Kazakhstan. Zootaxa 2016, 4175, 261–273. [Google Scholar] [CrossRef]
- Navabi, A.; Hajiqanbar, H.; Mortazavi, A. New records of the genus Allopygmephorus (Acari: Neopygmephoridae) with a list of the species of the family Neopygmephoridae in Iran. Persian J. Acarol. 2018, 7, 315–322. [Google Scholar]
- Mahunka, S. The Scientific Results of the Hungarian Soil Zoological Expeditions to South America. 9. Acari: Pyemotidae and Scutacaridae from the Guayaramerin region in Bolivia. Acta Zool. Acad. Sci. Hung. 1969, 15, 63–90. [Google Scholar]
- Short, A.E.Z.; Hebauer, F. World Catalogue of Hydrophiloidea—Additions and corrections, 1 (1999–2005) (Coleoptera). Koleopterol. Rundsch. 2006, 76, 315–359. [Google Scholar]
- Short, A.E.Z.; Fikáček, M. World catalogue of the Hydrophiloidea (Coleoptera): Additions and corrections II (2006–2010). Acta Entomol. Musei Natl. Pragae 2011, 51, 83–122. [Google Scholar]
- Jia, F.; Aston, P.; Fikáček, M. Review of the Chinese species of the genus Coelostoma Brullé, 1835 (Coleoptera: Hydrophilidae: Sphaeridiinae). Zootaxa 2014, 3887, 354–376. [Google Scholar] [PubMed]
- Fikáček, M. Neotropical leaf litter beetle genus Motonerus (Coleoptera: Hydrophilidae): New species, distribution data, and description of third instar larva. Neotrop. Entomol. 2019, 48, 788–808. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mite Species | Distribution | Host and/or Habitat | Reference |
|---|---|---|---|
| Allopygmephorus bakaninae Khaustov and Ermilov, 2008 | Russia | In wet soil | [37] |
| Allopygmephorus baoshanensis Gao, Zou and Ma, 1989 | China | Unknown | [38] |
| Allopygmephorus brasiliensis Mahunka, 1970 | Brazil | Litter and humus among roots surrounded by water | [39] |
| Allopygmephorus chinensis Mahunka 1975 | Hong Kong | Wet and rotting leaves | [40] |
| Allopygmephorus cunae Mahunka, 1970 | Brazil | Very wet decaying grass and detritus on river shore | [39] |
| Allopygmephorus heterodactylus Mahunka, 1973 | Ghana | Unidentified beetle; Cercyon sp. (Col.: Hydrophilidae) | [41] |
| Allopygmephorus matthesi Krczal, 1959 | Germany; Tanzania; Iran | Berosus luridus (Linnaeus, 1760), Coelostoma orbiculare (Fabricius, 1775), Enochrus quadripunctatus (Herbst, 1797), Helochares griseus (Fabricius, 1787), H. lividus (Forster, 1771), Hydrobius fuscipes (Linnaeus, 1758), Hydrophilus caraboides (Linnaeus, 1758), Philydrus melanocephalus Kuwert, 1888 (Col.: Hydrophilidae); Heterocerus marginatus (Fabricius, 1787) (Col.: Heteroceridae); Dryops auriculatus (Geoffroy, 1785) (Col.: Dryopidae); Xyleborus sp. (Col.: Curculionidae: Scolytinae) | [41,42,43] |
| Allopygmephorus nanhuiensis Gao, Zou and Ma, 1989 | China | Unknown | [38] |
| Allopygmephorus orientalis Mahunka and Mahunka-Papp, 1988 | Malaysia | Rotting wood and leaves | [44] |
| Allopygmephorus persicus Khaustov and Hajiqanbar, 2006 | Iran | Cercyon laminates Sharp, 1873, Enochrus bicolor (Fabricius, 1792) (Col.: Hydrophilidae) | [36,45] |
| Allopygmephorus punctatus Khaustov and Sazhnev, 2016 | Russia; Iran | Heterocerus fenestratus (Thunberg, 1784), Heterocerus flexuosus Stephens, 1828 (Col.: Heteroceridae); Drasterius bimaculatus (Rossi, 1790) (Col.: Elateridae); Augyles sp. (Col.: Heteroceridae) | [46,47] |
| Allopygmephorus spinisetus Khaustov and Sazhnev, 2016 | Russia Iran | Heterocerus fenestratus (Col.: Heteroceridae); Drasterius bimaculatus (Col.: Elateridae); Augyles sp. (Col.: Heteroceridae) | [46,47] |
| Allopygmephorus tuberosus Mahunka, 1969 | Bolivia | Unknown | [48] |
| Allopygmephorus coelostomus sp. nov. | Australia | Coelostoma (Coelostoma) fabricii (Col.: Hydrophilidae) | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khadem-Safdarkhani, H.; Hajiqanbar, H.; Riegler, M.; Seeman, O.; Katlav, A. Two New Phoretic Species of Heterostigmatic Mites (Acari: Prostigmata: Neopygmephoridae and Scutacaridae) on Australian Hydrophilid Beetles (Coleoptera: Hydrophilidae). Insects 2022, 13, 483. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050483
Khadem-Safdarkhani H, Hajiqanbar H, Riegler M, Seeman O, Katlav A. Two New Phoretic Species of Heterostigmatic Mites (Acari: Prostigmata: Neopygmephoridae and Scutacaridae) on Australian Hydrophilid Beetles (Coleoptera: Hydrophilidae). Insects. 2022; 13(5):483. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050483
Chicago/Turabian StyleKhadem-Safdarkhani, Hamid, Hamidreza Hajiqanbar, Markus Riegler, Owen Seeman, and Alihan Katlav. 2022. "Two New Phoretic Species of Heterostigmatic Mites (Acari: Prostigmata: Neopygmephoridae and Scutacaridae) on Australian Hydrophilid Beetles (Coleoptera: Hydrophilidae)" Insects 13, no. 5: 483. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050483