
Seasonal Variations of Spodoptera frugiperda Host Plant Diversity and Parasitoid Complex in Southern and Central Benin

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
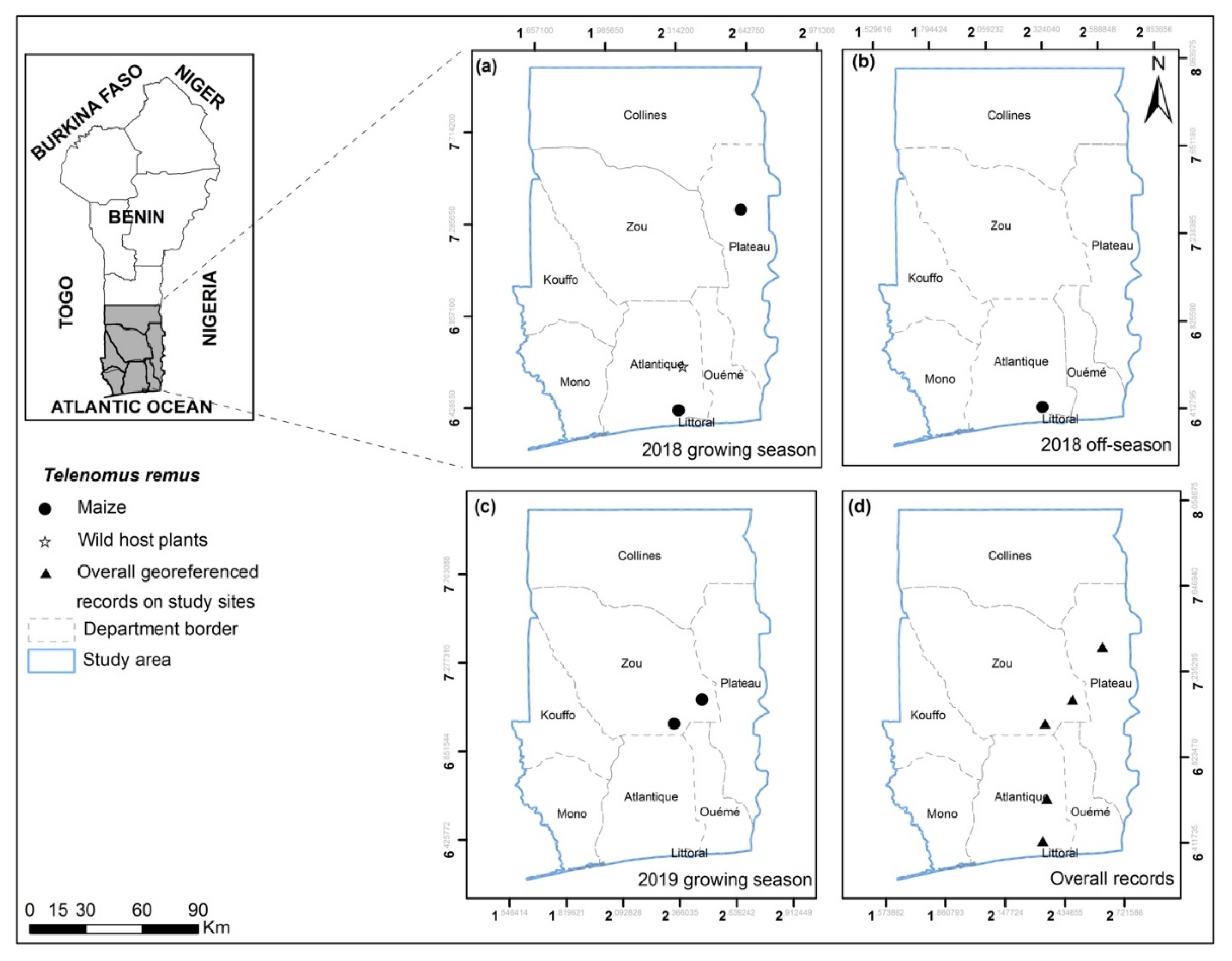
2.1. Study Sites
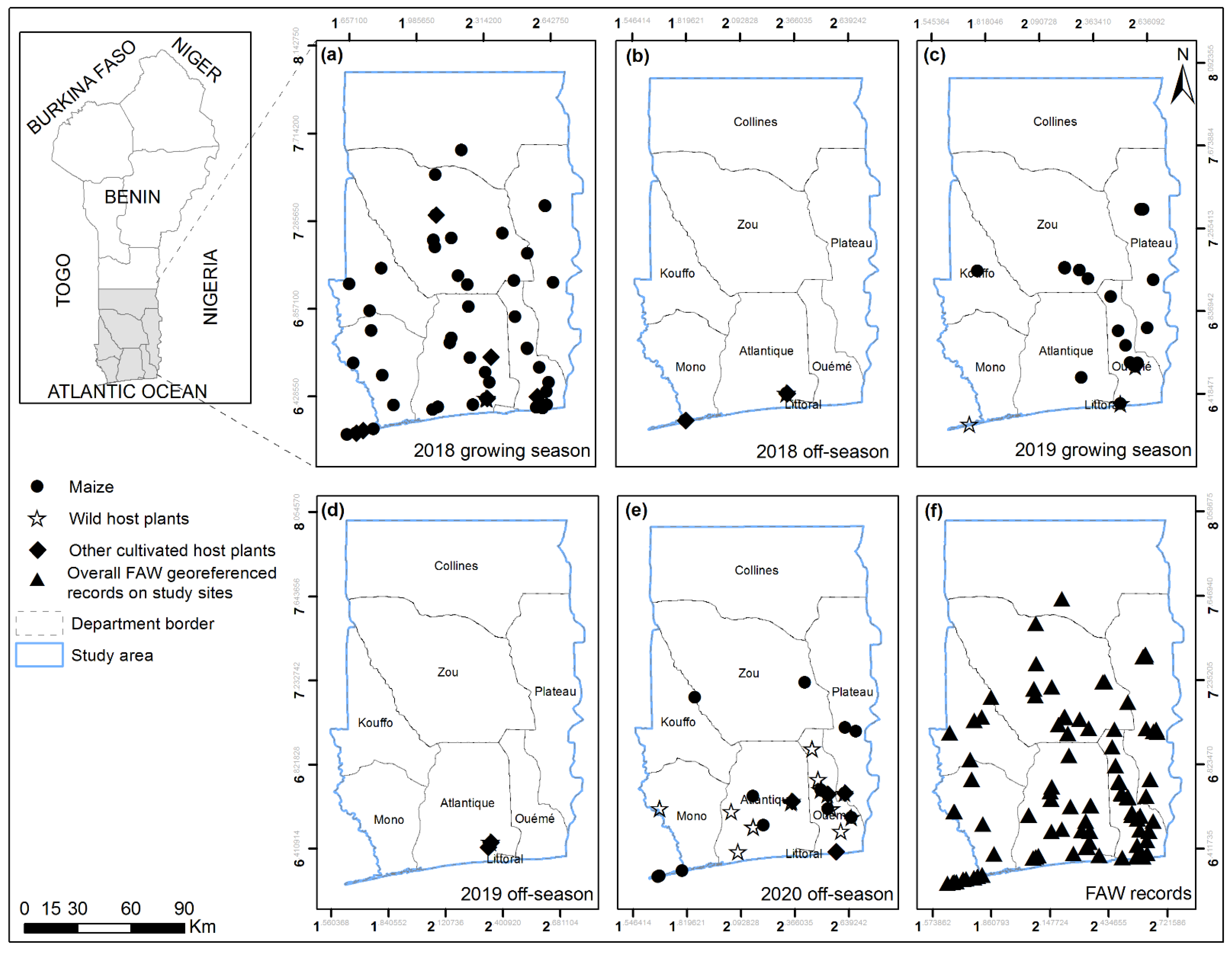
2.2. Field Surveys
2.3. Data Analysis
3. Results
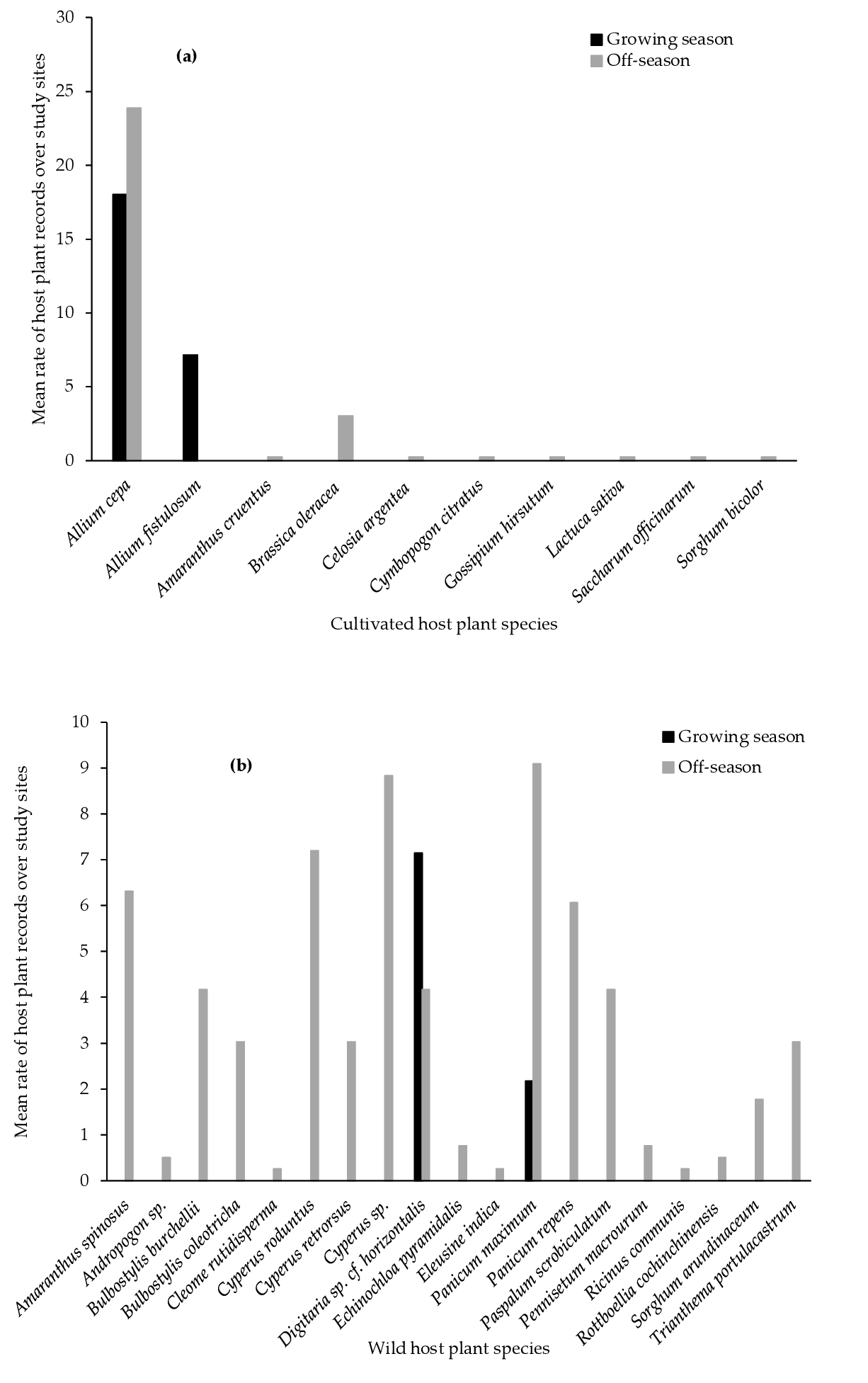
3.1. FAW Host Plants Range
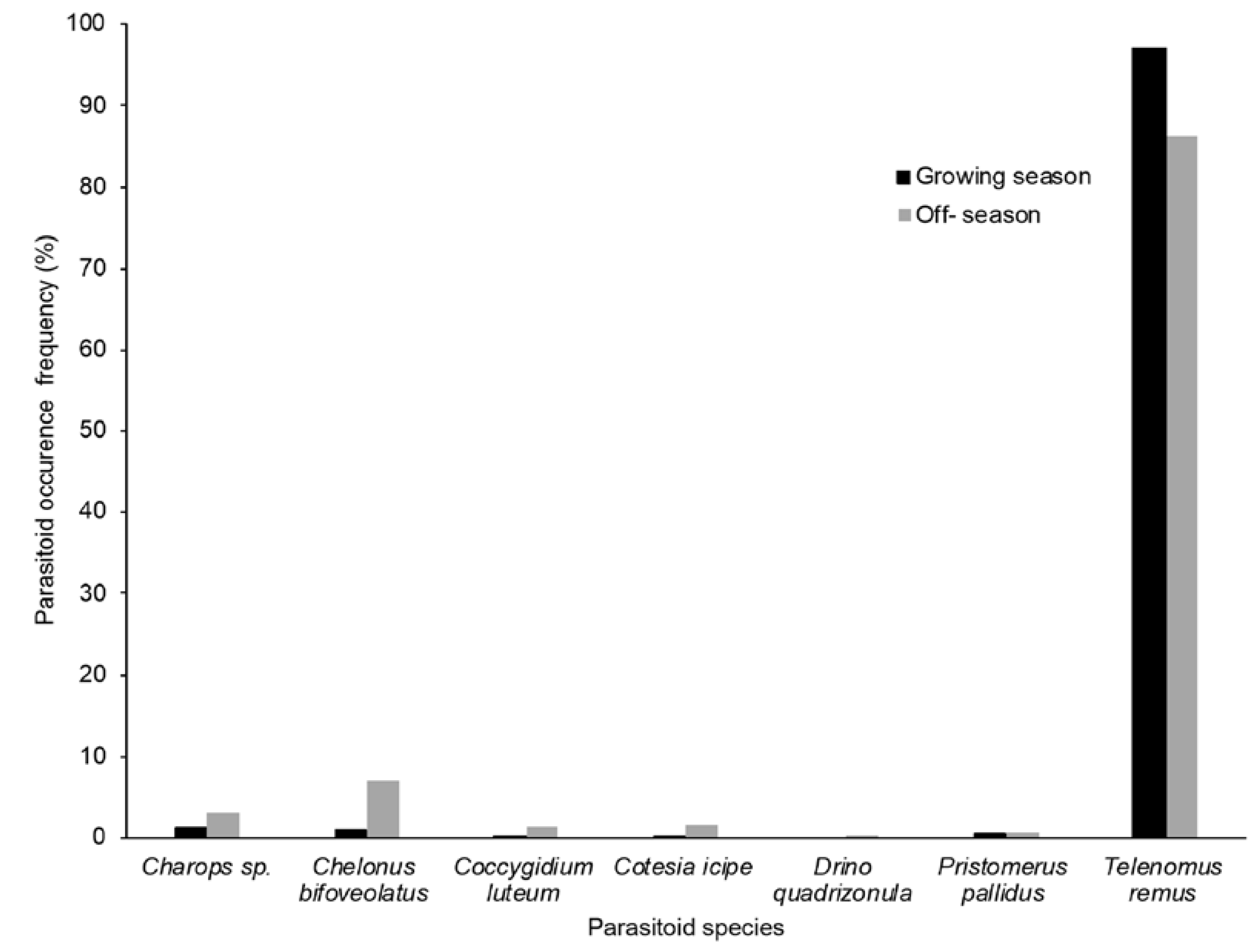
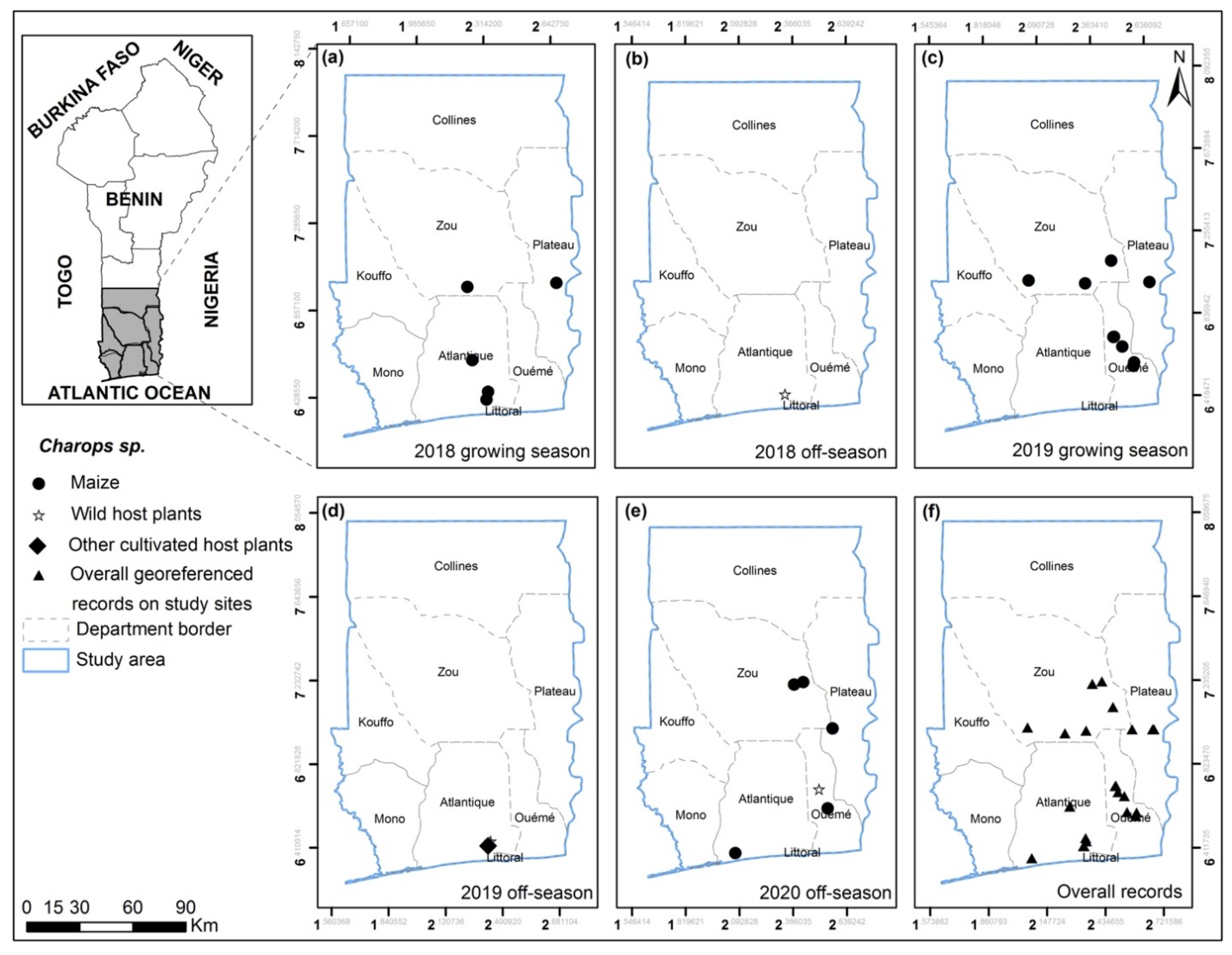
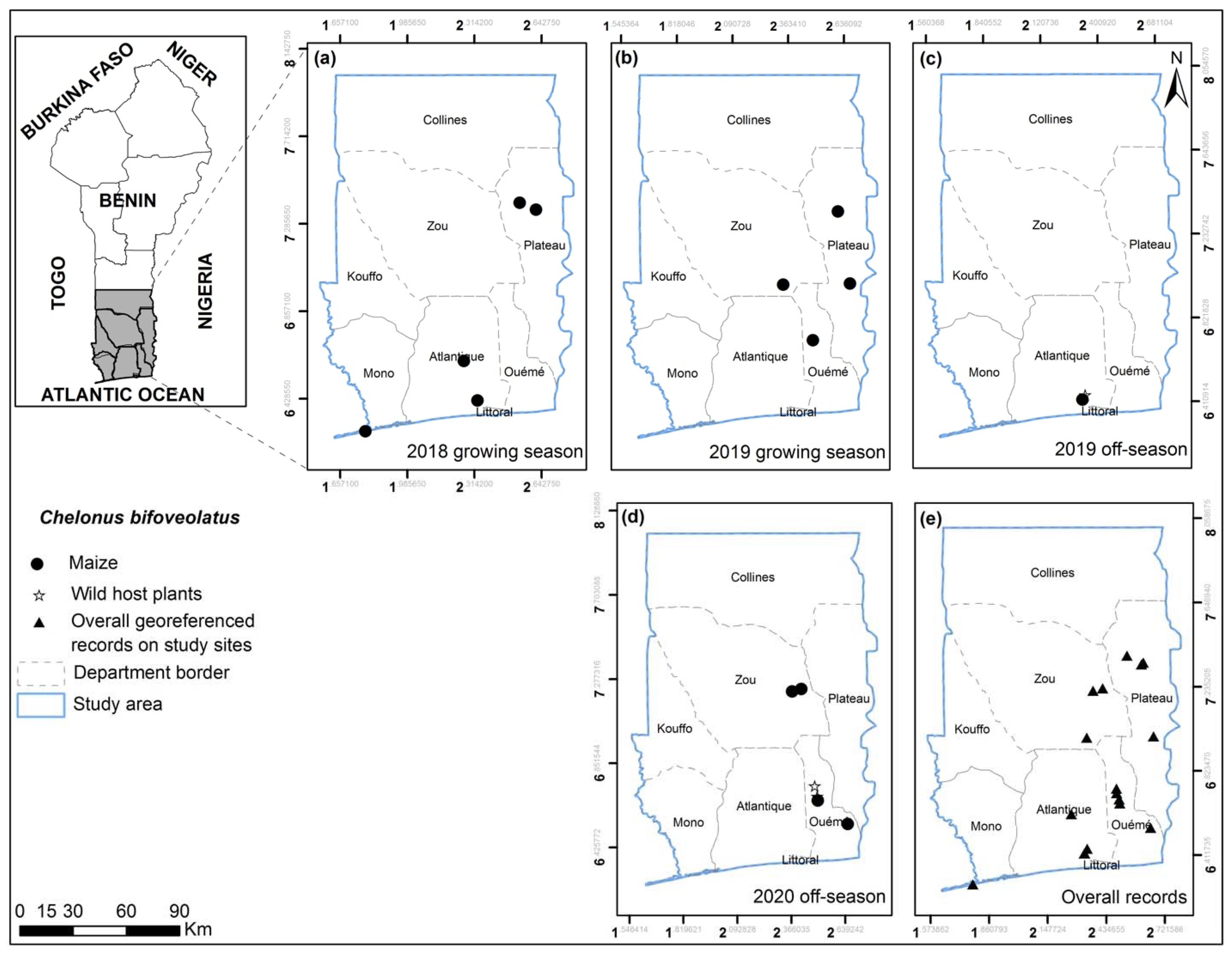
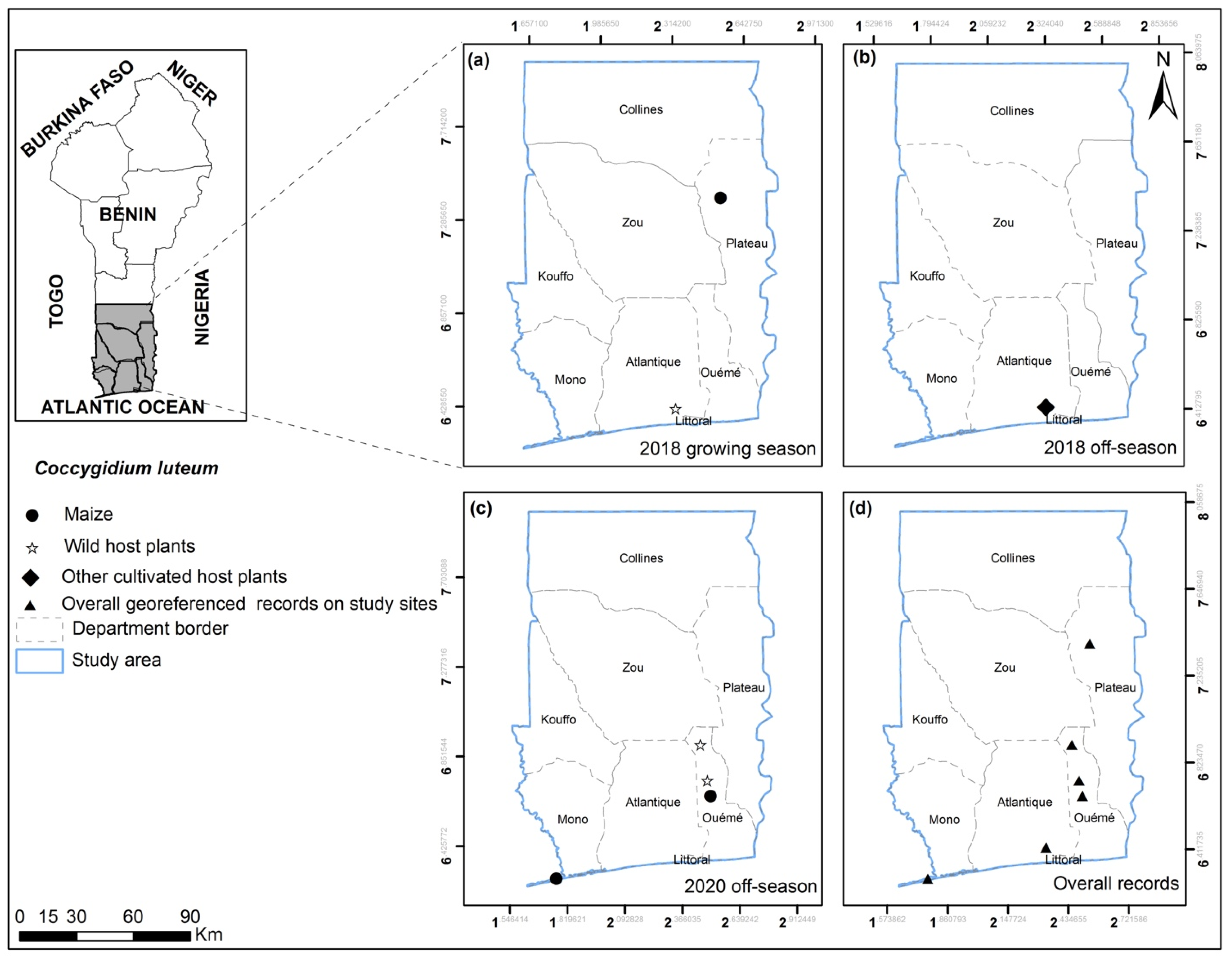
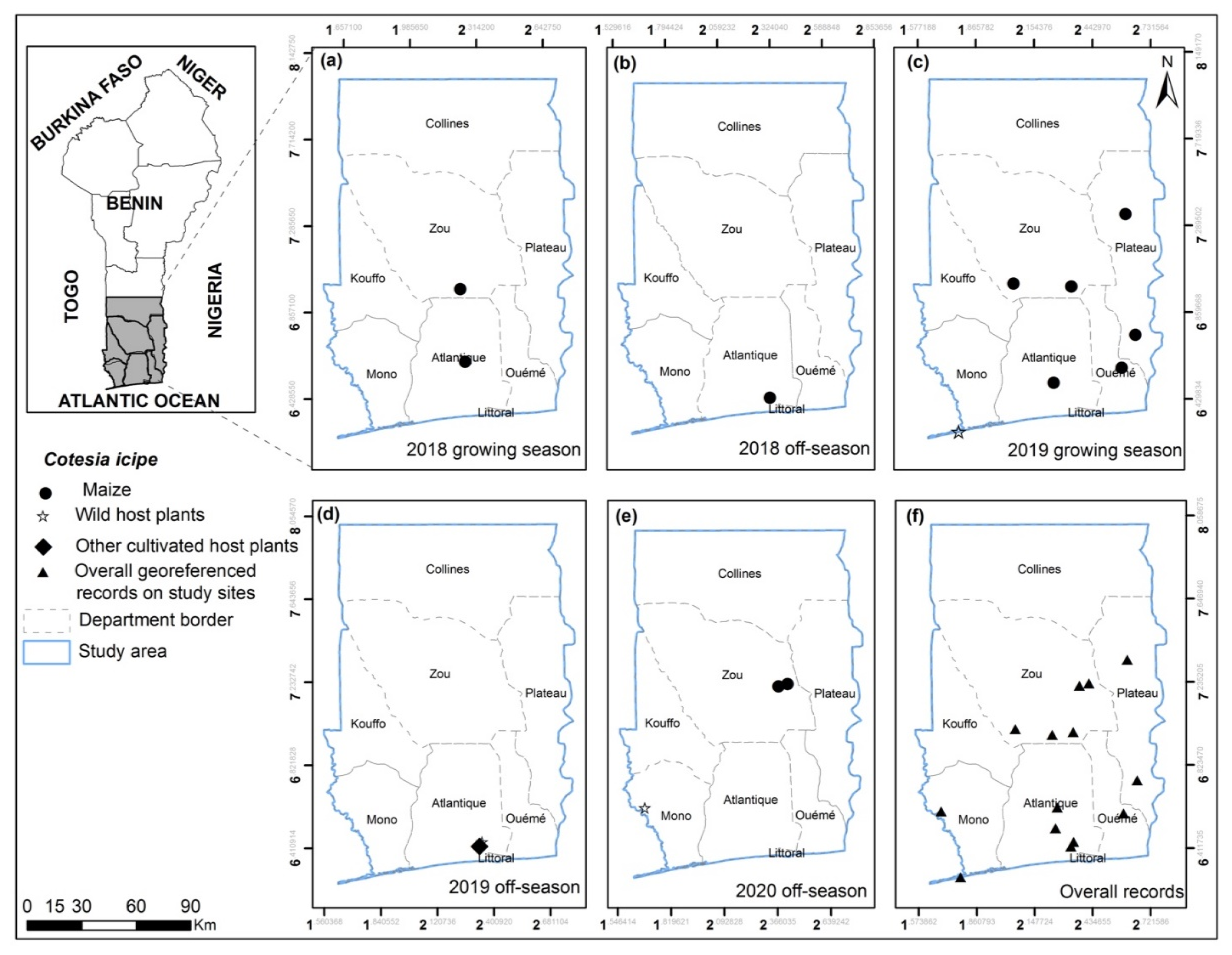
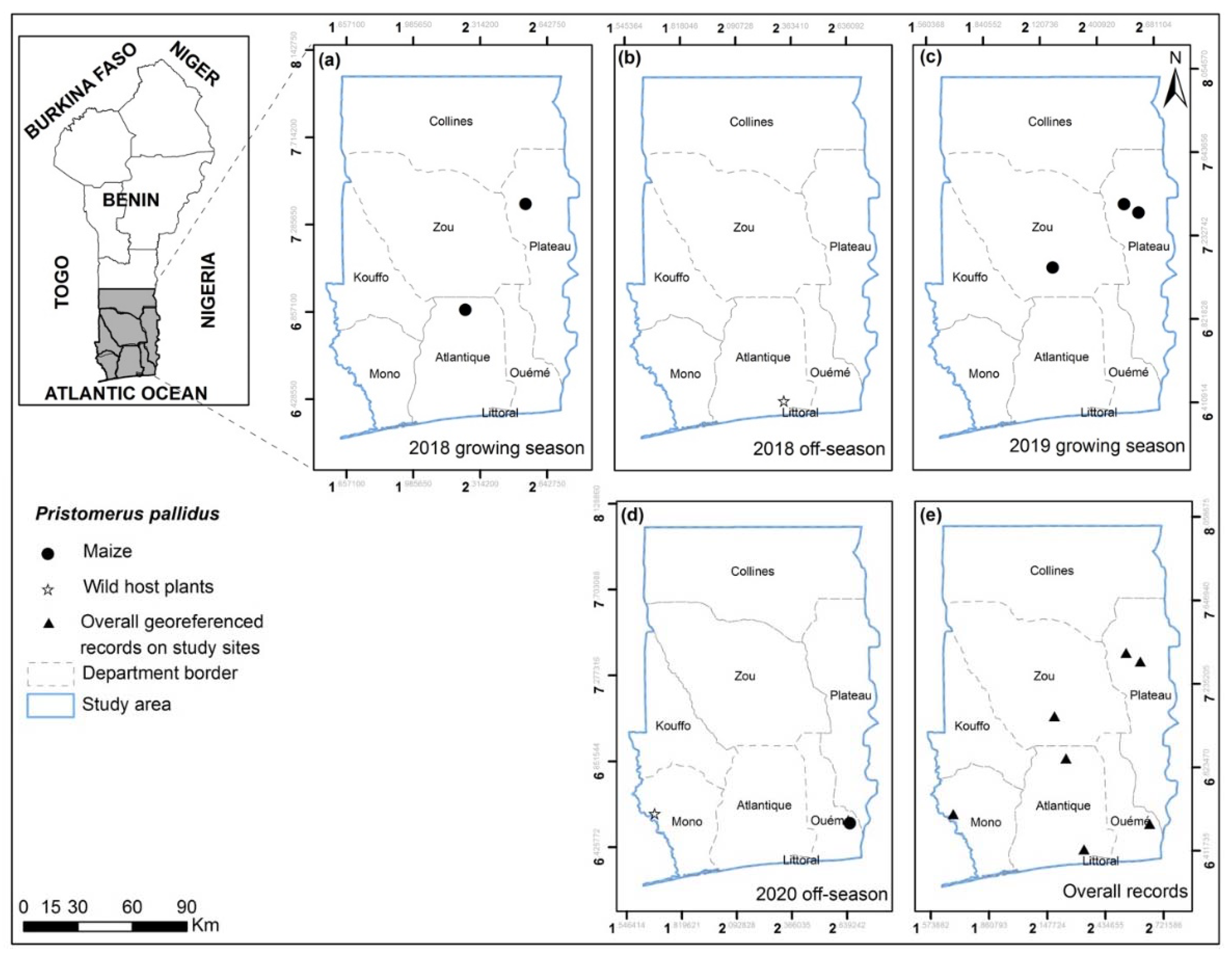
3.2. FAW Parasitoid Complex
4. Discussion
4.1. FAW Host Plant Range
4.2. FAW Parasitoid Complex
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Casmuz, A.; Socías, M.G.; Murúa, M.G.; Prieto, S.; Medina, S.; Willink, E.; Gastaminza, G. Revisión de los hospederos del gusano cogollero del maíz, Spodoptera frugiperda (Lepidoptera: Noctuidae). Entomológica Argent. 2010, 24, 209–231. [Google Scholar]
- Montezano, D.G.; Sosa-Gómez, D.R.; Paula-Moraes, S.V.; Roque-Specht, V.F.; Fronza, E.; Barros, N.M.; Specht, A. Biotic Potential and Reproductive Parameters of Spodoptera dolichos (Lepidoptera: Noctuidae) in the Laboratory. Zool. Curitiba 2015, 32, 485–491. [Google Scholar] [CrossRef] [Green Version]
- Pogue, M.G. A World Revision of the Genus Spodoptera (Guenée) (Lepidoptera: Noctuidae). Available online: https://bugguide.net/node/view/815156 (accessed on 22 June 2021).
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First Report of Outbreaks of the Fall Armyworm Spodoptera frugiperda (J.E. Smith) (Lepidoptera, Noctuidae), a New Alien Invasive Pest in West and Central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Groote, H.; Kimenju, S.C.; Munyua, B.; Palmas, S.; Kassie, M.; Bruce, A. Spread and Impact of Fall Armyworm (Spodoptera frugiperda J.E. Smith) in Maize Production Areas of Kenya. Agric. Ecosyst. Environ. 2020, 292, 106804. [Google Scholar] [CrossRef] [PubMed]
- FAO Global Action for Fall Armyworm Control 2021. Available online: https://www.fao.org/fall-armyworm/en/ (accessed on 15 October 2021).
- Njuguna, E.; Nethononda, P.; Maredia, K.; Mbabazi, R.; Kachapulula, P.; Rowe, A.; Ndolo, D. Experiences and Perspectives on Spodoptera frugiperda (Lepidoptera: Noctuidae) Management in Sub-Saharan Africa. J. Integr. Pest Manag. 2021, 12, 7. [Google Scholar] [CrossRef]
- Sisay, B.; Simiyu, J.; Malusi, P.; Likhayo, P.; Mendesil, E.; Elibariki, N.; Wakgari, M.; Ayalew, G.; Tefera, T. First Report of the Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), Natural Enemies from Africa. J. Appl. Entomol. 2018, 142, 800–804. [Google Scholar] [CrossRef]
- Houngbo, S.; Zannou, A.; Aoudji, A.; Sossou, H.C.; Sinzogan, A.; Sikirou, R.; Zossou, E.; Vodounon, H.S.T.; Adomou, A.; Ahanchédé, A. Farmers’ Knowledge and Management Practices of Fall Armyworm, Spodoptera frugiperda (J.E. Smith) in Benin, West Africa. Agriculture 2020, 10, 430. [Google Scholar] [CrossRef]
- De Freitas Bueno, R.C.O.; de Freitas Bueno, A.; Moscardi, F.; Postali Parra, J.R.; Hoffmann-Campo, C.B. Lepidopteran Larva Consumption of Soybean Foliage: Basis for Developing Multiple-Species Economic Thresholds for Pest Management Decisions: Lepidopteran Larva Consumption of Soybean Foliage. Pest Manag. Sci. 2011, 67, 170–174. [Google Scholar] [CrossRef] [Green Version]
- Hardke, J.T.; Lorenz, G.M.; Leonard, B.R. Fall Armyworm (Lepidoptera: Noctuidae) Ecology in Southeastern Cotton. J. Integr. Pest Manag. 2015, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host Plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Molina-Ochoa, J.; Carpenter, J.E.; Heinrichs, E.A.; Foster, J.E. Parasitoids and Parasites of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas and Caribbean Basin: An Inventory. Fla. Entomol. 2003, 86, 254–289. [Google Scholar] [CrossRef]
- Kenis, M.; du Plessis, H.; Van den Berg, J.; Ba, M.; Goergen, G.; Kwadjo, K.; Baoua, I.; Tefera, T.; Buddie, A.; Cafà, G.; et al. Telenomus remus, a Candidate Parasitoid for the Biological Control of Spodoptera frugiperda in Africa, is Already Present on the Continent. Insects 2019, 10, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agboyi, L.K.; Goergen, G.; Beseh, P.; Mensah, S.A.; Clottey, V.A.; Glikpo, R.; Buddie, A.; Cafà, G.; Offord, L.; Day, R.; et al. Parasitoid Complex of Fall Armyworm, Spodoptera frugiperda, in Ghana and Benin. Insects 2020, 11, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Issa, S.U.; Frimpong-Anin, K.; Adama, I.; Brandford, M.; Braimah, H.; Obeng, P. Indigenous Natural Enemies Attacking Fall Armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae) in Ghana. J. Entomol. 2020, 18, 1–7. [Google Scholar] [CrossRef]
- Da Silva, D.M.; Bueno, A.d.F.; Andrade, K.; Stecca, C.d.S.; Neves, P.M.O.J.; de Oliveira, M.C.N. Biology and Nutrition of Spodoptera frugiperda (Lepidoptera: Noctuidae) Fed on Different Food Sources. Sci. Agric. 2017, 74, 18–31. [Google Scholar] [CrossRef]
- Lewter, J.A.; Szalanski, A.L.; Nagoshi, R.N.; Meagher, R.L.; Owens, C.B.; Luttrell, R.G. Genetic Variation within and between Strains of the Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). Fla. Entomol. 2006, 89, 63–68. [Google Scholar] [CrossRef]
- Nagoshi, R.N.; Silvie, P.; Meagher, R.L.; Lopez, J.; Machado, V. Identification and Comparison of Fall Armyworm (Lepidoptera: Noctuidae) Host Strains in Brazil, Texas, and Florida. Ann. Entomol. Soc. Am. 2007, 100, 394–402. [Google Scholar] [CrossRef]
- Barros, E.M.; Torres, J.B.; Ruberson, J.R.; Oliveira, M.D. Development of Spodoptera frugiperda on Different Hosts and Damage to Reproductive Structures in Cotton: Fall Armyworm Performance on Different Hosts. Entomol. Exp. Appl. 2010, 137, 237–245. [Google Scholar] [CrossRef]
- Andrews, K.L. The Worlworm, Spodoptera frugiperda, in Central America and Neighboring Areas. Fla. Entomol. 1980, 63, 456–467. [Google Scholar] [CrossRef]
- Segoli, M.; Kishinevsky, M.; Rozenberg, T.; Hoffmann, I. Parasitoid Abundance and Community Composition in Desert Vineyards and Their Adjacent Natural Habitats. Insects 2020, 11, 580. [Google Scholar] [CrossRef]
- Tepa-Yotto, G.T.; Tonnang, H.E.Z.; Goergen, G.; Subramanian, S.; Kimathi, E.; Abdel-Rahman, E.M.; Flø, D.; Thunes, K.H.; Fiaboe, K.K.M.; Niassy, S.; et al. Global Habitat Suitability of Spodoptera frugiperda (JE Smith) (Lepidoptera, Noctuidae): Key Parasitoids Considered for Its Biological Control. Insects 2021, 12, 273. [Google Scholar] [CrossRef] [PubMed]
- Guerrieri, E.; Poppy, G.M.; Powell, W.; Tremblay, E.; Pennacchio, F. Induction and Systemic Release of Herbivore-Induced Plant Volatiles Mediating in-Flight Orientation of Aphidius ervi. J. Chem. Ecol. 1999, 25, 15. [Google Scholar] [CrossRef]
- Molina-Ochoa, J.; Carpenter, J.E.; Lezama-Gutiérrez, R.; Foster, J.E.; González-Ramírez, M.; Angel-Sahagún, C.A.; Farías-Larios, J. Natural Distribution of Hymenopteran Parasitoids of Spodoptera frugiperda (Lepidoptera: Noctuidae) Larvae in Mexico. Fla. Entomol. 2004, 87, 461–472. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host Plant Type | Family Name | Scientific Name | Sites of Collection |
|---|---|---|---|
| Cultivated | Amaranthaceae | Amaranthus cruentus L. | Sèmè |
| Celosia argentea L. | Azowlissè | ||
| Amaryllidaceae | Allium cepa L. | Abomey-Calavi, Agoué, Grand-Popo, Sèmè, Zinvié | |
| Allium fistulosum L. | Abomey-Calavi | ||
| Asteraceae | Lactuca sativa L. | Sèmè | |
| Brassicaceae | Brassica oleracea L. | Abomey-Calavi | |
| Malvaceae | Gossypium hirsutum L. | Dan | |
| Poaceae | Cymbopogon citratus (de Candolle) Stapf | Sèmè | |
| Saccharum officinarum L. | Zinvié | ||
| Sorghum bicolor (L.) Mönch | Sakété | ||
| Wild | Aizoaceae | Trianthema portulacastrum L. | Sèmè |
| Amaranthaceae | Amaranthus spinosus L. | Grand-Popo, Sèmè | |
| Cleomaceae | Cleome rutidosperma de Candolle | Azowlissè | |
| Cyperacea | Bulbostylis burchellii (Ficalho and Hiern) C.B. Clarke | Abomey-Calavi | |
| Cyperacea | Bulbostylis coleotricha (A. Richard) Clarke | Abomey-Calavi | |
| Cyperus rotundus L. | Abomey-Calavi | ||
| Cyperus retrorsus Chapman | Abomey-Calavi | ||
| Cyperus sp. | Adjohoun, Tori-Avamè | ||
| Euphorbiaceae | Ricinus communis L. | Zinvié | |
| Poaceae | Andropogon sp. | Adjohoun, Athiémé | |
| Digitaria sp. cf. horizontalis Willdenow | Abomey-Calavi | ||
| Echinochloa pyramidalis(Lamarck) Hitchcock and Chase | Zinvié, Kpomassè | ||
| Eleusine indica (L.) Grtner | Tori-Avamè | ||
| Panicum maximum Jacquin | Abomey-Calavi, Zinvié, Misserete, Bonou, Dangbo, Adjohoun | ||
| Panicum repens L. | Abomey-Calavi | ||
| Paspalum scrobiculatum L. | Abomey-Calavi | ||
| Pennisetum macrourumvon Trinius | Adjohoun, Azowlissè | ||
| Sorghum arundinaceum (Desvaux.) Stapf. | Aguégués, Athiémé, Avrankou, Tori-Avamè, Ouidah, Bonou | ||
| Rottboellia cochinchinensis (Loureiro) W.D.Clayton | Sakété, Athiémé |
| Order, Family and Species | Locality | Host Plant | FAW Stage Collected |
|---|---|---|---|
| Hymenoptera: Platygastridae | |||
| Telenomus remus Nixon | Ab-Cal; Aid (Ket); Zinv; Ouin; Zog | Maize; Panicum maximum | Egg |
| Hymenoptera: Braconidae Chelonus bifoveolatus Szépligeti | GP; Ag; Ket; Aid (Ket); Glo; Ab-Cal; Adj; Kpa; Pob; Zog; Avr; Zag; Azo | Maize; Bulbostylis coleotrica; Panicum maximum; Andropogon sp. | Larva; Egg |
| Coccygidium luteum (Brullé) | Ket; Ab-Cal; Bon; Adj; Azo; GP | Maize; Digitaria cf. horizontalis; Allium cepa; Sorghum arundinaceum; Cyperus sp.; Panicum maximum | Larva |
| Cotesia icipe Fernandez-Triana and Fiaboe | Glo; Mas; GP; Dra; Kpa; Sak; Mis; Ab-Cal; Zog; Zag; Ath | Maize; Amaranthus spinosus; Bulbostylis coleotrica; Allium cepa; Sorghum arundinaceum | Larva |
| Hymenoptera: Ichneumonidae Charops sp. | Glod; Mas; Pob; Ab-Cal; Dan; Adj; Mis; Pob; Azo; Ak-Mis; Ouin; Zog; Ab-Cal; Azo; Zag; All; Dan; Adja; Adj | Maize; Digitaria sp.; Allium cepa; Bulbostylis coleotrica; Andropogon sp. | Larva |
| Pristomerus pallidus (Kriechbaumer) | Ket; Hou (Ag); Ab-Cal; Kpa; Zog; Ath; Avr | Maize; Cyperus roduntus; Sorghum arundinaceum | Larva |
| Diptera: Tachinidae Drino quadrizonula (Thomson) | Ab-Cal | Maize | Larva |
| Order, Family and Species | Alternatives Host Plants | Number of Collected Specimens |
|---|---|---|
| Hymenoptera: Platygastridae Telenomus remus | Panicum maximum | 100 |
| Hymenoptera: Braconidae Chelonus bifoveolatus | ||
| Bulbostylis coleotrica | 1 | |
| Panicum maximum | 2 | |
| Andropogon sp. | 1 | |
| Coccygidium luteum | Digitaria cf. horizontalis | 1 |
| Allium cepa | 1 | |
| Sorghum arundinaceum | 1 | |
| Cyperus sp. | 1 | |
| Panicum maximum | 2 | |
| Cotesia icipe | Amaranthus spinosus | 1 |
| Bulbostylis coleotrica | 4 | |
| Allium cepa | 1 | |
| Sorghum arundinaceum | 1 | |
| Hymenoptera: Ichneumonidae | Digitaria sp. | 1 |
| Charops sp. | Allium cepa | 3 |
| Bulbostylis coleotrica | 3 | |
| Andropogon sp. | 3 | |
| Pristomerus pallidus | Cyperus roduntus | 1 |
| Sorghum arundinaceum | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Winsou, J.K.; Tepa-Yotto, G.T.; Thunes, K.H.; Meadow, R.; Tamò, M.; Sæthre, M.-G. Seasonal Variations of Spodoptera frugiperda Host Plant Diversity and Parasitoid Complex in Southern and Central Benin. Insects 2022, 13, 491. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13060491
Winsou JK, Tepa-Yotto GT, Thunes KH, Meadow R, Tamò M, Sæthre M-G. Seasonal Variations of Spodoptera frugiperda Host Plant Diversity and Parasitoid Complex in Southern and Central Benin. Insects. 2022; 13(6):491. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13060491
Chicago/Turabian StyleWinsou, Jeannette K., Ghislain T. Tepa-Yotto, Karl H. Thunes, Richard Meadow, Manuele Tamò, and May-Guri Sæthre. 2022. "Seasonal Variations of Spodoptera frugiperda Host Plant Diversity and Parasitoid Complex in Southern and Central Benin" Insects 13, no. 6: 491. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13060491