
Abnormal Antennal Olfactory Sensilla Phenotypes Involved in Olfactory Deficit in Bactrocera correcta (Diptera: Tephritidae)

 ,
, 


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Chemicals
2.3. Scanning Electron Microscopy
2.4. Electrophysiological Recordings
2.5. Behavioral Bioassay
2.6. Statistical Analysis
3. Results
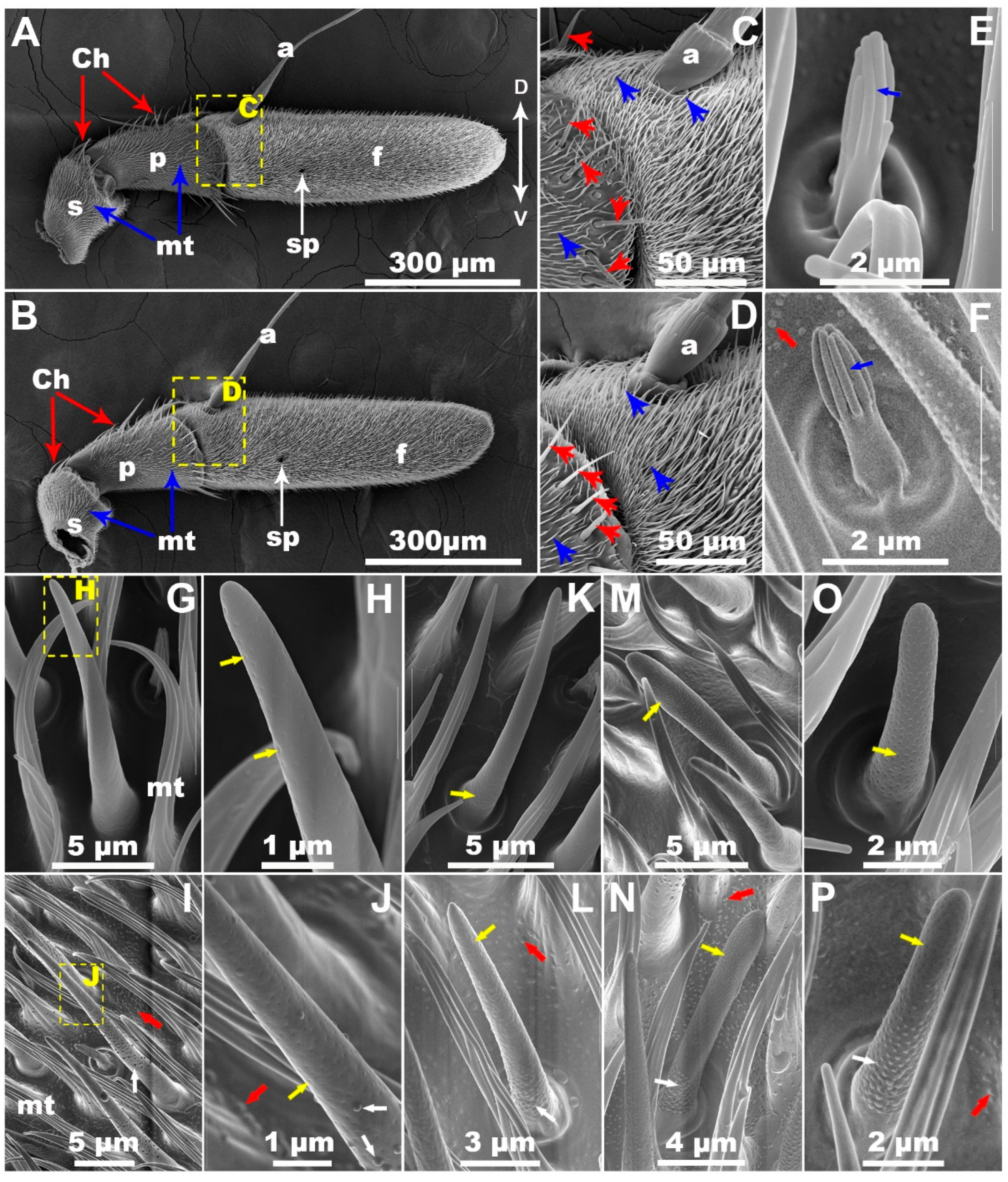
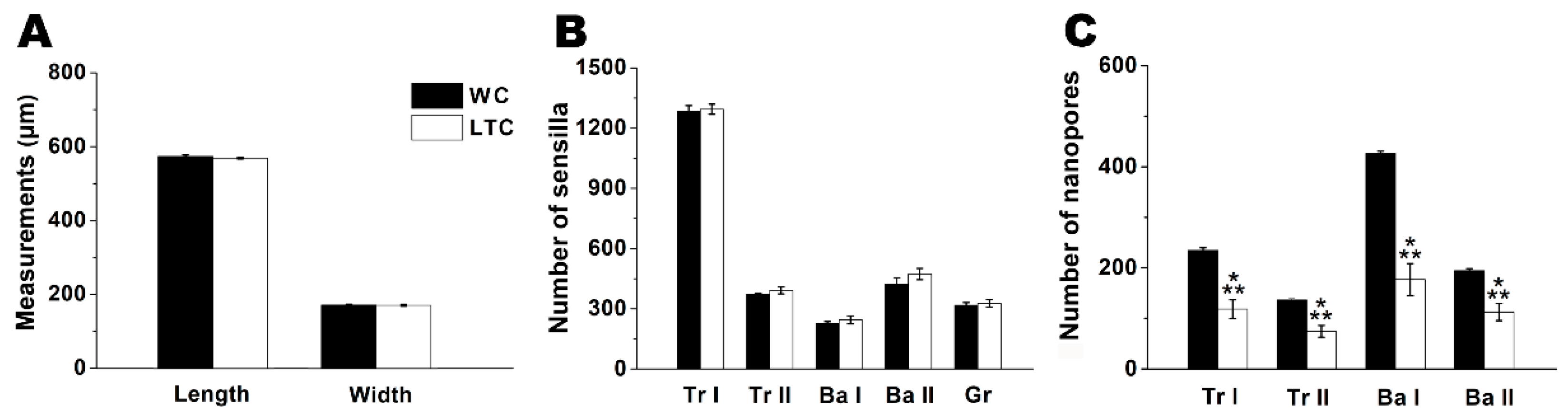
3.1. Antennal Sensilla of LTC and WC
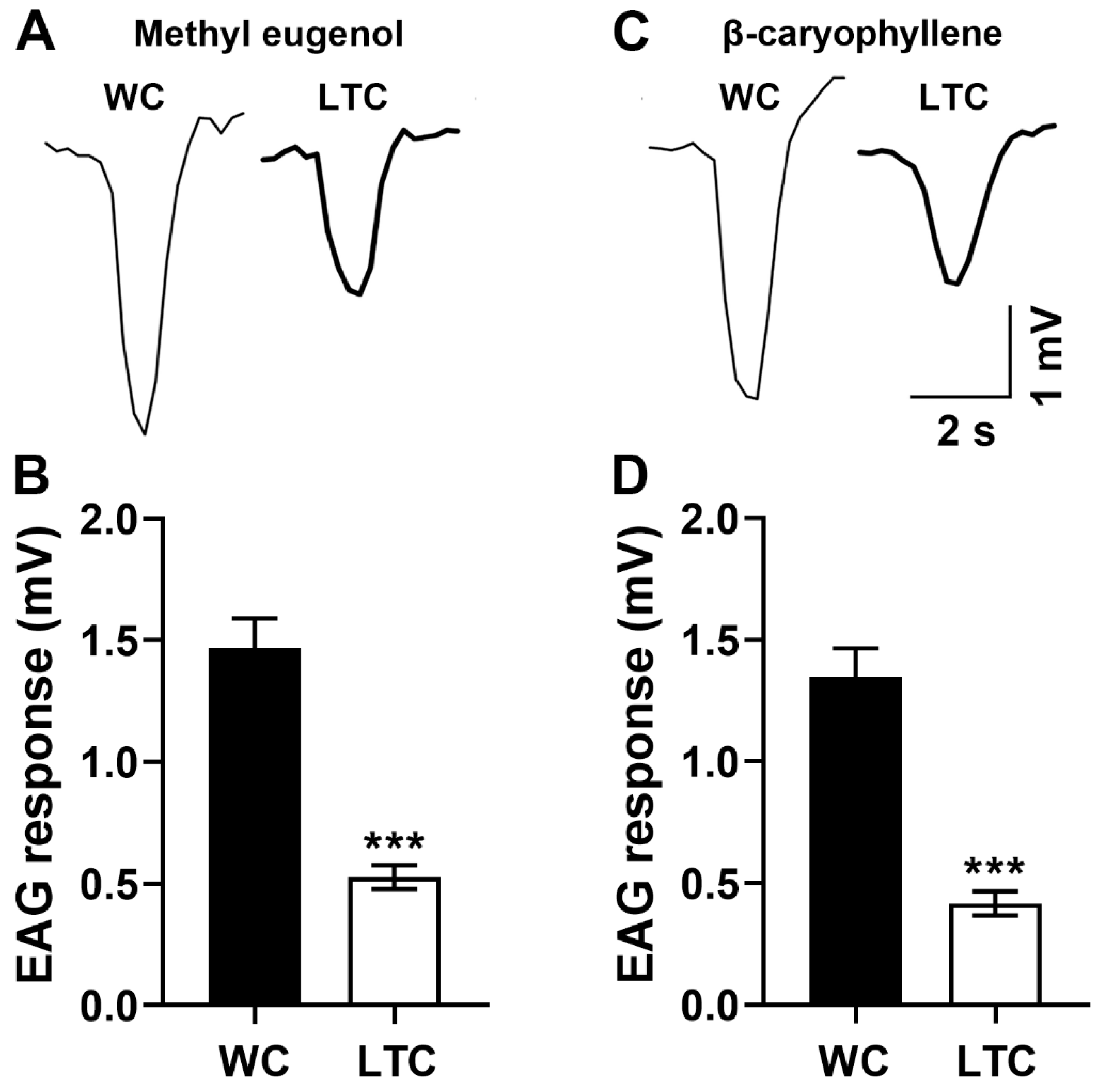
3.2. The Abnormal Antennal Olfactory Sensilla Phenotypes Inhibited Antenna Function
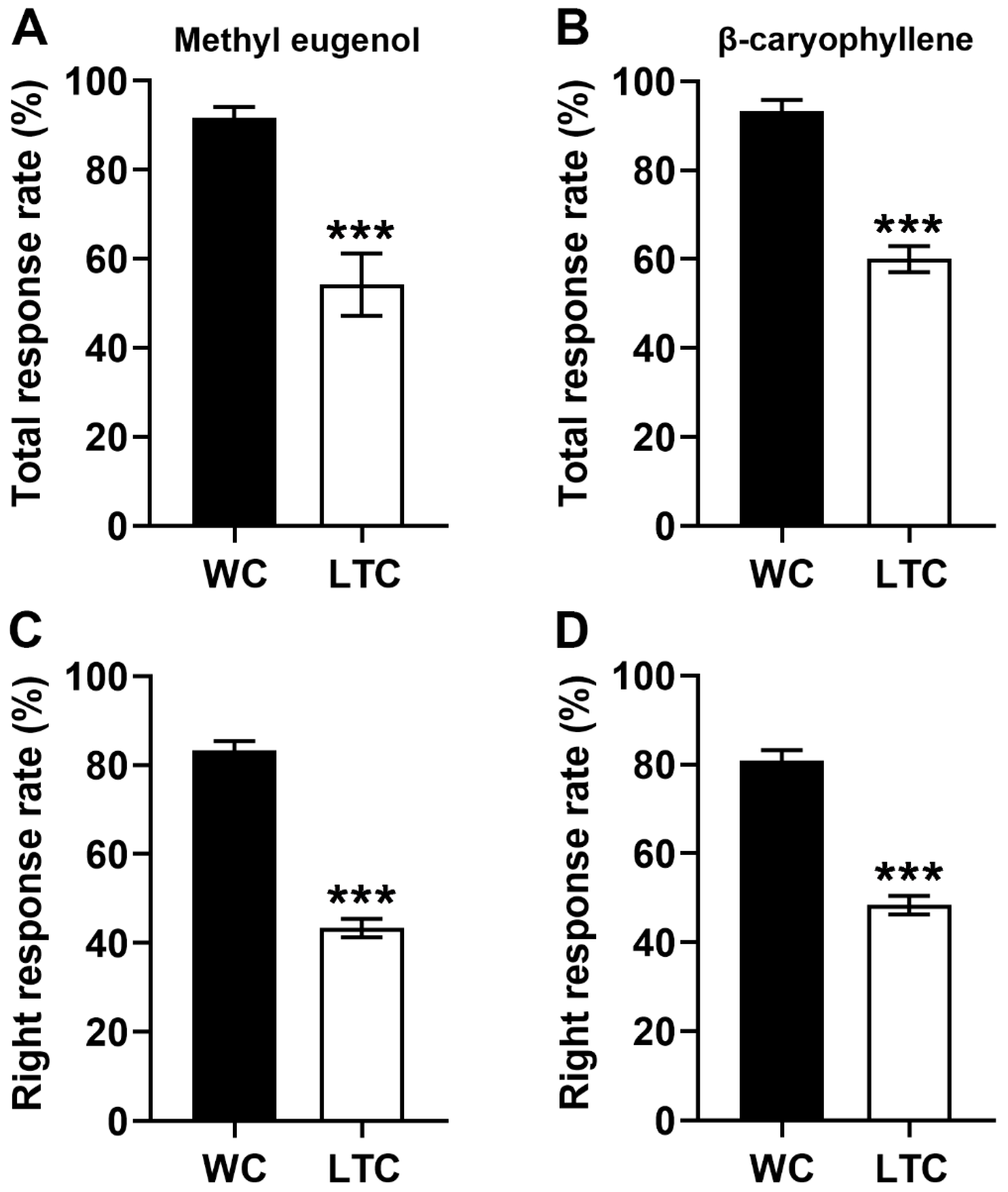
3.3. The Abnormal Antennal Olfactory Sensilla Phenotypes Impaired Olfactory Behavior
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, X.J. The fruit flies (Diptera: Tephritidae) of the East Asian region. Acta Zool. Sin. 1996, 21, 52. [Google Scholar]
- Vargas, R.I.; Piñero, J.C.; Leblanc, L. An overview of pest species of Bactrocera fruit flies (Diptera: Tephritidae) and the integration of biopesticides with other biological approaches for their management with a focus on the Pacific region. Insects 2015, 6, 297–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, I.M.; Elson-Harris, M.M. Fruit Flies of Economic Significance: Their Identification and Bionomics; CAB International: Wallingford, UK, 1992. [Google Scholar]
- Allwood, A.J.; Chinajariyawong, A.; Drew, R.A.I.; Hamacek, E.L.; Hancock, D.L.; Hengsawad, C.; Jinapin, J.; Jirasurat, M.; Krong, K.C.; Leong, C.T.S.; et al. Host plant records for fruit flies (Diptera: Tephritidae) in South East Asia. Raffles Bull. Zool. 1999, 47 (Suppl. S7), 1–92. [Google Scholar]
- United States Department of Agriculture. Provisional List of Host Plants of Guava Fruit Fly, Bactrocera correcta (Bezzi) (Diptera: Tephritidae). Available online: https://www.aphis.usda.gov/plant_health/plant_pest_info/fruit_flies/downloads/host-lists//bactrocera-correcta-provisional-host-plants.pdf (accessed on 1 November 2021).
- Drew, R.A.I.; Raghu, S. The fruit fly fauna (Diptera: Tephritidae: Dacinae) of the rainforest habitat of the Western Ghats, India. Raffles Bull. Zool. 2002, 50, 327–352. [Google Scholar]
- Liu, X.F.; Jin, Y.; Ye, H. Recent spread and climatic ecological niche of the invasive guava fruit fly, Bactrocera correcta, in mainland China. J. Pest Sci. 2013, 86, 449–458. [Google Scholar] [CrossRef]
- Shanbhag, S.R.; Müller, B.; Steinbrecht, R.A. Atlas of olfactory organs of Drosophila melanogaster: 1. Types, external organization, innervation and distribution of olfactory sensilla. Int. J. Insect Morphol. Embryol. 1999, 28, 377–397. [Google Scholar] [CrossRef]
- Yan, X.Z.; Deng, C.P.; Xie, J.X.; Wu, L.J.; Sun, X.J.; Hao, C. Distribution patterns and morphology of sensilla on the antennae of Plutella xylostella (L.)-A scanning and transmission electron microscopic study. Micron 2017, 103, 1–11. [Google Scholar] [CrossRef]
- Schneider, D. Insect olfaction: Deciphering system for chemical messages. Science 1969, 163, 1031–1037. [Google Scholar] [CrossRef]
- Zacharuk, R.Y. Ultrastructure and function of insect chemosensilla. Annu. Rev. Entomol. 1980, 25, 27–47. [Google Scholar] [CrossRef]
- Ando, T.; Sekine, S.; Inagaki, S.; Misaki, K.; Badel, L.; Moriya, H.; Sami, M.M.; Itakura, Y.; Chihara, T.; Kazama, H.; et al. Nanopore formation in the cuticle of an insect olfactory sensillum. Curr. Biol. 2019, 29, 1512–1520. [Google Scholar] [CrossRef]
- Oh, H.W.; Jeong, S.A.; Kim, J.; Park, K.C. Morphological and functional heterogeneity in olfactory perception between antennae and maxillary palps in the pumpkin fruit fly, Bactrocera depressa. Arch. Insect Biochem. Physiol. 2019, 101, e21560. [Google Scholar] [CrossRef] [PubMed]
- Steinbrecht, R.A. Olfactory receptors. In Atlas of Arthropod Sensory Receptors-Dynamic Morphology in Relation to Function; Eguchi, E., Tominaga, Y., Eds.; Springer: Tokyo, Japan, 1999; pp. 155–176. [Google Scholar]
- Lee, W.Y.; Chang, J.C.; Hwang, Y.; Lin, T.L. Morphology of the Antennal Sensilla of the Oriental Fruit Fly, Dacus dorsalis Hendel (Diptera: Tephrltidae). Zool. Stud. 1994, 33, 65–71. Available online: http://zoolstud.sinica.edu.tw/Journals/33.1/65.pdf (accessed on 1 November 2021).
- Hu, F.; Zhang, G.N.; Jia, F.X.; Dou, W.; Wang, J.J. Morphological characterization and distribution of antennal sensilla of six fruit flies (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 2010, 103, 661–670. [Google Scholar] [CrossRef]
- Zheng, W.; Zhu, C.; Peng, T.; Zhang, H. Odorant receptor co-receptor Orco is upregulated by methyl eugenol in male Bactrocera dorsalis (Diptera: Tephritidae). J. Insect Physiol. 2012, 58, 1122–1127. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, X.F.; Fu, L.; Han, Y.Y.; Chen, J.; Lu, Y.Y. BdorOBP2 plays an indispensable role in the perception of methyl eugenol by mature males of Bactrocera dorsalis (Hendel). Sci. Rep. 2017, 7, 15894. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Liang, X.F.; Xu, L.; Keesey, I.W.; Lei, Z.R.; Smagghe, G.; Wang, J.J. An antennae-specific odorant-binding protein is involved in Bactrocera dorsalis olfaction. Front. Ecol. Evol. 2020, 8, 63. [Google Scholar] [CrossRef] [Green Version]
- Wee, S.L.; Chinvinijkul, S.; Tan, K.H.; Nishida, R. A new and highly effective male lure for the guava fruit fly Bactrocera correcta. J. Pest Sci. 2018, 91, 691–698. [Google Scholar] [CrossRef]
- Tokushima, I.; Orankanok, W.; Tan, K.H.; Ono, H.; Nishida, R. Accumulation of phenylpropanoid and sesquiterpenoid volatiles in male rectal pheromonal glands of the guava fruit fly, Bactrocera correcta. J. Chem. Ecol. 2010, 36, 1327–1334. [Google Scholar] [CrossRef]
- De Rose, F.; Corda, V.; Solari, P.; Sacchetti, P.; Belcari, A.; Poddighe, S.; Kasture, S.; Solla, P.; Marrosu, F.; Liscia, A. Drosophila mutant model of parkinson’s disease revealed an unexpected olfactory performance: Morphofunctional evidences. Parkinson’s Dis. 2016, 2016, 3508073. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.W.; Wen, P.; Qu, Y.F.; Dong, S.H.; Li, J.J.; Tan, K.; Nieh, J.C. Bees eavesdrop upon informative and persistent signal compounds in alarm pheromones. Sci. Rep. 2016, 6, 25693. [Google Scholar] [CrossRef]
- Gu, H.Y.; O’Dowd, D.K. Cholinergic synaptic transmission in adult Drosophila Kenyon cells in situ. J. Neurosci. 2006, 26, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.G.; Wei, C.M.; Miao, J.; Zhang, X.J.; Wei, B.; Dong, W.X.; Xiao, C. Chemical compounds from female and male rectal pheromone glands of the guava fruit fly, Bactrocera correcta. Insects 2019, 10, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, S.Z.; Li, X.Y.; Wang, Z.M.; Wang, J.B.; Han, D.Y.; Fan, J.H.; Zhao, Q.; Liu, H.; Wang, X.S. Assessment of 2-allyl-4,5-dimethoxyphenol safety and attractiveness to mature males of Bactrocera dorsalis (Hendel). Ecotoxicol. Environ. Saf. 2021, 223, 112567. [Google Scholar] [CrossRef] [PubMed]
- Ju, Q.; Guo, X.Q.; Li, X.; Jiang, X.J.; Jiang, X.G.; Ni, W.L.; Qu, M.J. Plant volatiles increase sex pheromone attraction of Holotrichia parallela (Coleoptera: Scarabaeoidea). J. Chem. Ecol. 2017, 43, 236–242. [Google Scholar] [CrossRef]
- Wigglesworth, V.B. The insect cuticle. Biol. Rev. 1948, 23, 408–451. [Google Scholar] [CrossRef]
- Keil, T.A.; Steiner, C. Morphogenesis of the antenna of the male silkmoth. Antheraea polyphemus, Ⅲ. Development of olfactory sensilla and the properties of hair-forming cells. Tissue Cell. 1991, 23, 821–851. [Google Scholar] [CrossRef]
- Ray, K.; Rodrigues, V. Cellular events during development of the olfactory sense organs in Drosophila melanogaster. Dev. Biol. 1995, 167, 426–438. [Google Scholar] [CrossRef] [Green Version]
- Jhaveri, D.; Sen, A.; Rodrigues, V. Mechanisms underlying olfactory neuronal connectivity in Drosophila-the atonal lineage organizes the periphery while sensory neurons and glia pattern the olfactory lobe. Dev. Biol. 2000, 226, 73–87. [Google Scholar] [CrossRef] [Green Version]
- Dorer, D.R.; Rudnick, J.A.; Moriyama, E.N.; Christensen, A.C. A family of genes clustered at the Triplo-lethal locus of Drosophila melanogaster has an unusual evolutionary history and significant synteny with Anopheles gambiae. Genetics 2003, 165, 613–621. [Google Scholar] [CrossRef]
- Shah, N.; Dorer, D.R.; Moriyama, E.N.; Christensen, A.C. Evolution of a large, conserved, and syntenic gene family in insects. G3-Genes Genom Genet. 2012, 2, 313–319. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.R.; Morandin, C.; Noureddine, M.; Pant, S. Conserved roles of Osiris genes in insect development, polymorphism and protection. J. Evolution. Biol. 2018, 31, 516–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derby, C.D.; Kozma, M.T.; Senatore, A.; Schmidt, M. Molecular mechanisms of reception and perireception in crustacean chemoreception: A comparative review. Chem. Senses 2016, 41, 381–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Olfactory Sensilla | WC | LTC | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Length (μm) | Middle Width (μm) | Wall Nanopore | Nanopore Diameter (nm) | Length (μm) | Middle Width (μm) | Wall Bulge | Wall Nanopore | Nanopore Diameter (nm) | |
| Grooved | 3.23 ± 0.07 e | 0.50 ± 0.01 e | – | – | 3.26 ± 0.06 e | 0.50 ± 0.01 e | – | – | – |
| Trichoid Ⅰ | 20.41 ± 0.22 a | 1.19 ± 0.08 b | + | 22.8 ± 0.3 b | 20.70 ± 0.18 a | 1.19 ± 0.01 b | + | + | 22.6 ± 0.3 b |
| Trichoid Ⅱ | 12.06 ± 0.15 b | 1.01 ± 0.01 d | + | 23.3 ± 0.2 b | 12.09 ± 0.10 b | 1.01 ± 0.01 d | + | + | 23.3 ± 0.2 b |
| Basiconica Ⅰ | 10.80 ± 0.22 c | 1.31 ± 0.01 a | + | 43.7 ± 0.2 a | 10.87 ± 0.15 c | 1.31 ± 0.01 a | + | + | 43.9 ± 0.3 a |
| Basiconica Ⅱ | 7.72 ± 0.19 d | 1.09 ± 0.01 c | + | 43.5 ± 0.1 a | 7.87 ± 0.09 d | 1.09 ± 0.01 c | + | + | 43.7 ± 0.2 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, K.-F.; Peng, X.-M.; Tu, J.-Y.; Jin, C.; Zhang, W.-R.; Chen, X.-Z.; Liu, Y.-J.; Zha, H.-G.; Shi, W.; Cao, J. Abnormal Antennal Olfactory Sensilla Phenotypes Involved in Olfactory Deficit in Bactrocera correcta (Diptera: Tephritidae). Insects 2022, 13, 535. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13060535
Guo K-F, Peng X-M, Tu J-Y, Jin C, Zhang W-R, Chen X-Z, Liu Y-J, Zha H-G, Shi W, Cao J. Abnormal Antennal Olfactory Sensilla Phenotypes Involved in Olfactory Deficit in Bactrocera correcta (Diptera: Tephritidae). Insects. 2022; 13(6):535. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13060535
Chicago/Turabian StyleGuo, Kai-Fei, Xiao-Mei Peng, Jie-Yu Tu, Chan Jin, Wan-Rong Zhang, Xi-Zhu Chen, Yong-Jun Liu, Hong-Guang Zha, Wei Shi, and Jun Cao. 2022. "Abnormal Antennal Olfactory Sensilla Phenotypes Involved in Olfactory Deficit in Bactrocera correcta (Diptera: Tephritidae)" Insects 13, no. 6: 535. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13060535