Differential Hatching, Development, Oviposition, and Longevity Patterns among Colombian Aedes aegypti Populations


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Mosquito Collections
2.2. Rearing and Maintenance of Ae. aegypti Populations
2.3. Effects of Temperature on Immature Stage Mortality
2.4. Biological Features and Statistical Analysis
3. Results
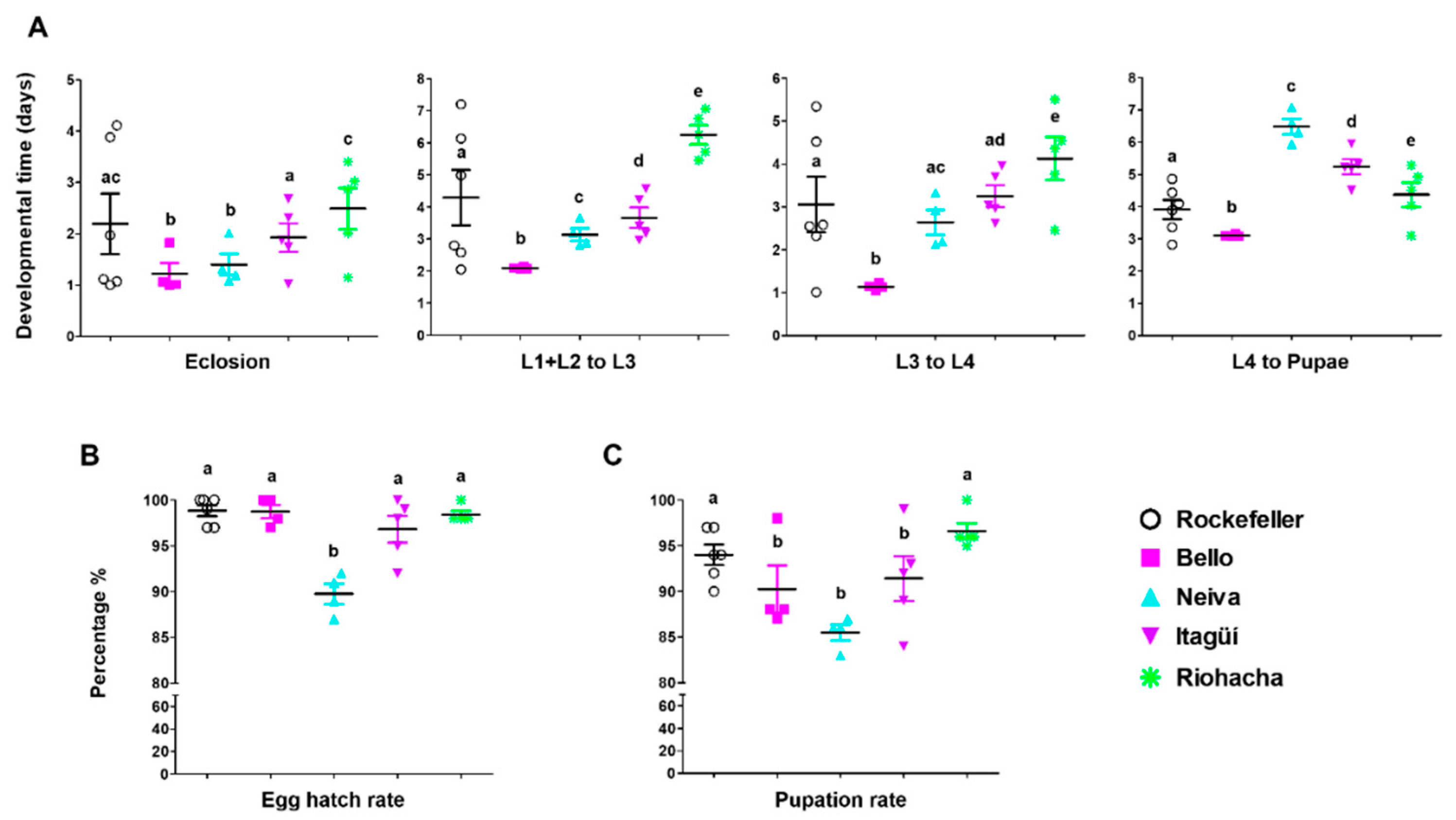
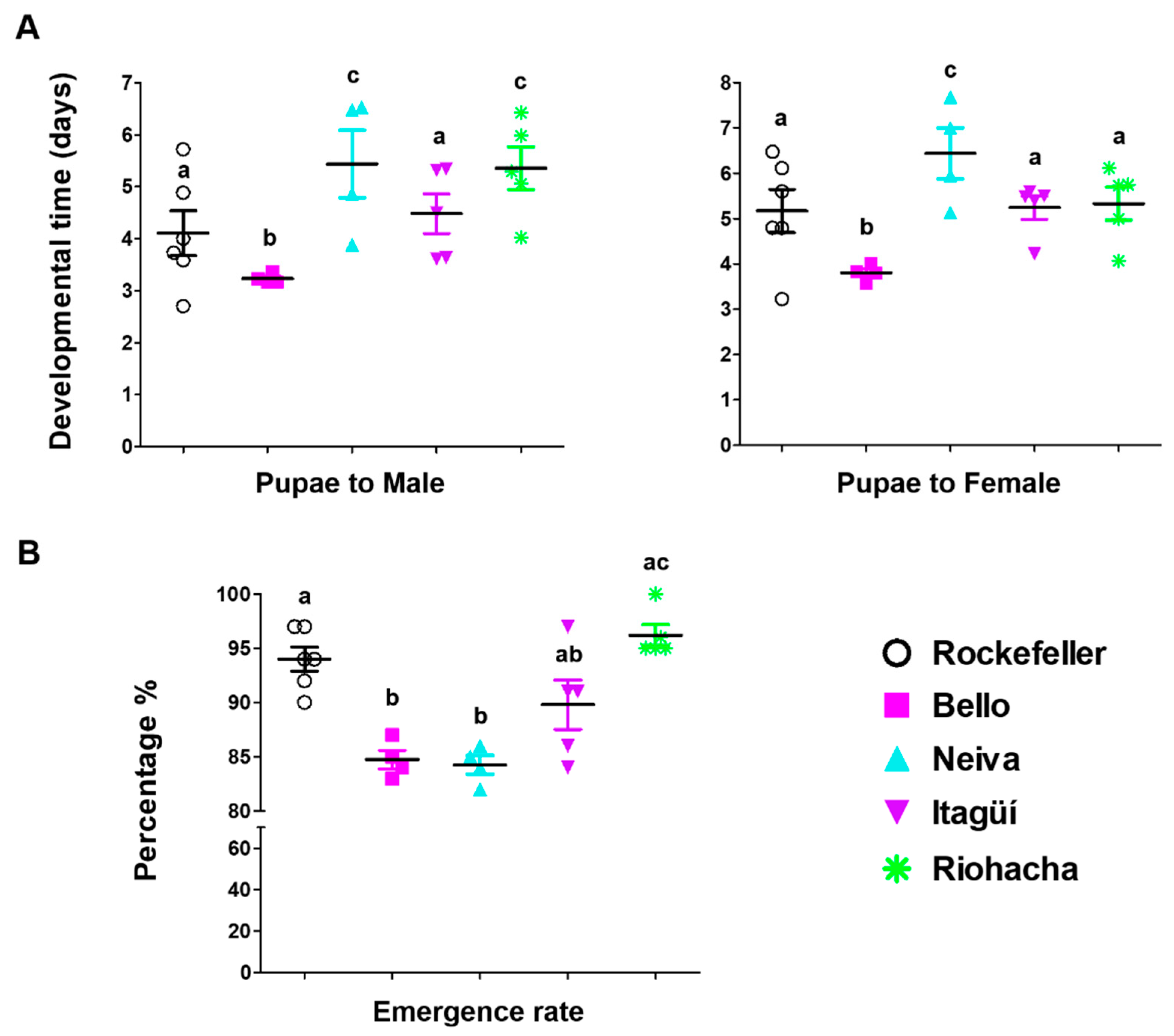
3.1. Differences in Immature Stage Development Times, Egg Hatching, Pupation, and Emergence Rates in Ae. aegypti Populations
3.2. Life Table Attributes of Immature Stages in Ae. aegypti Populations
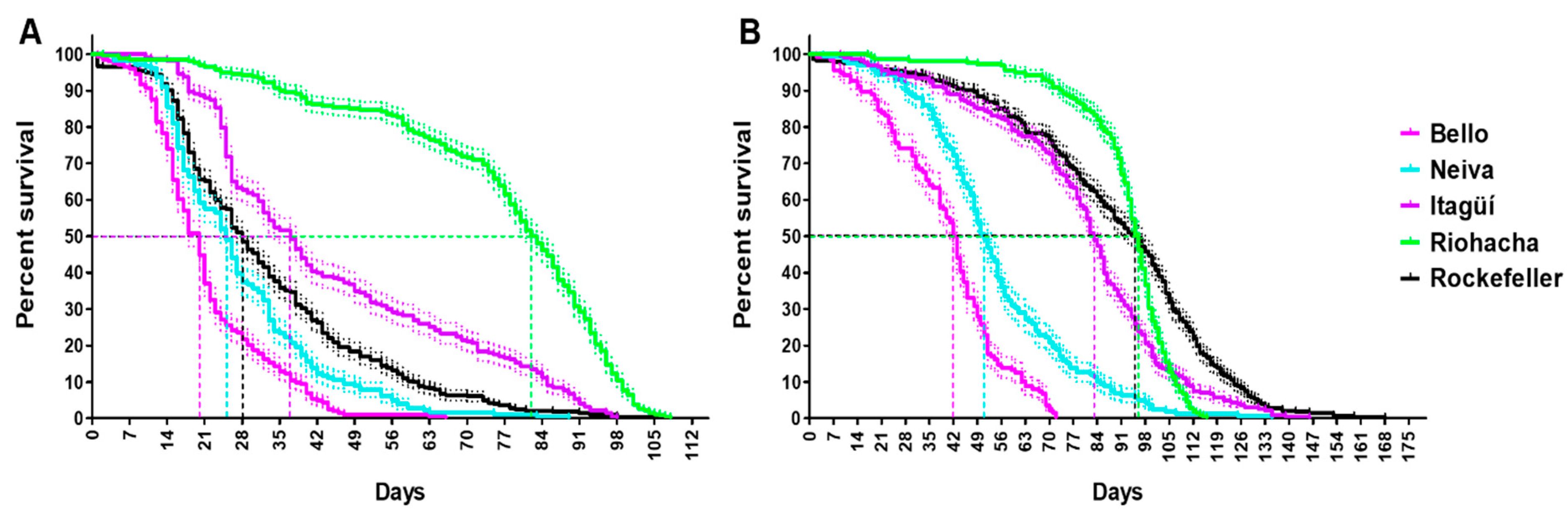
3.3. Sex Ratio, Longevity, and Survivorship of Adult Ae. aegypti Populations
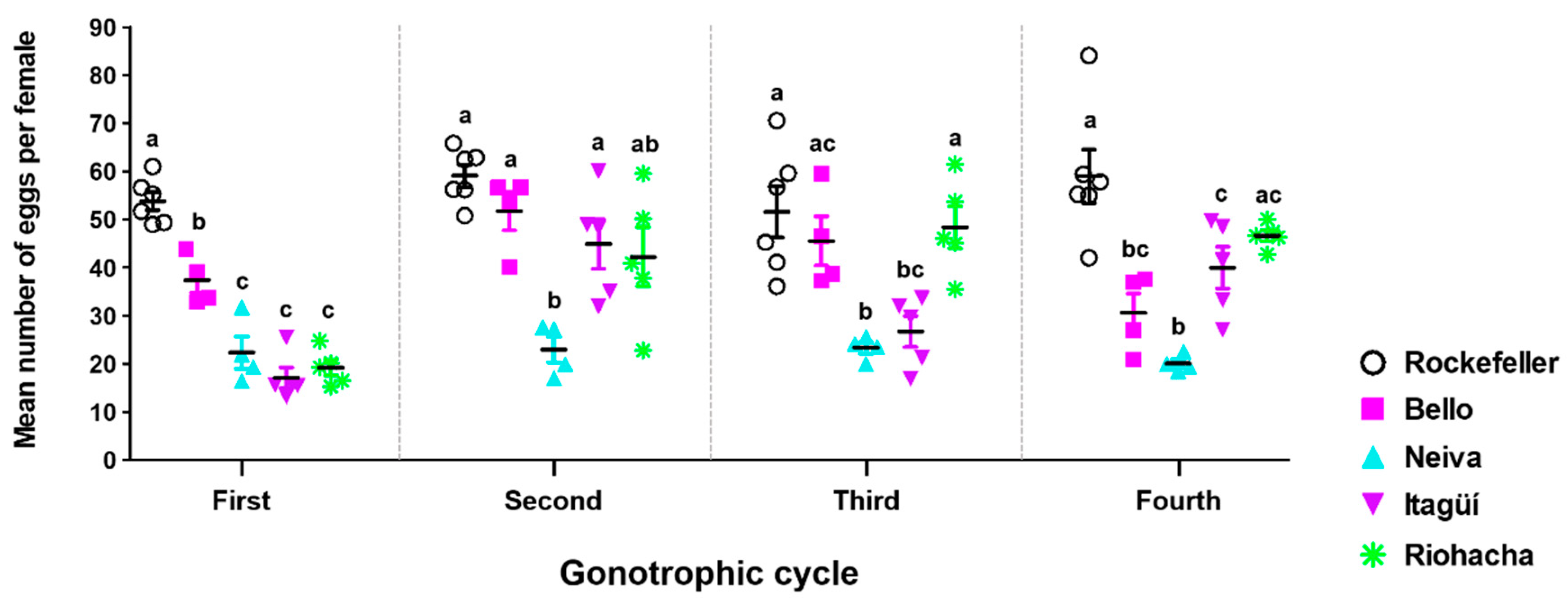
3.4. Fecundity Profile of Ae. aegypti Populations
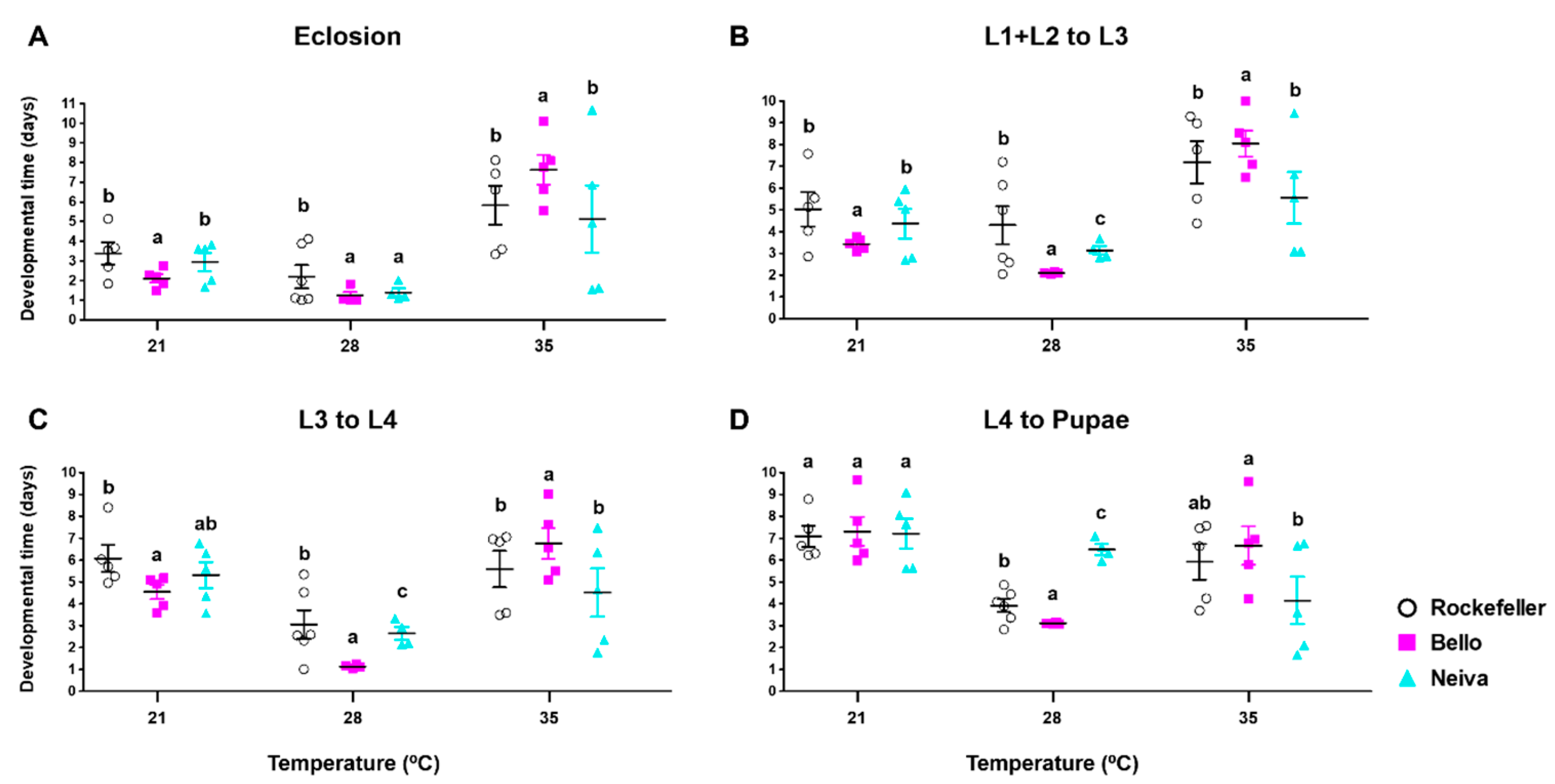
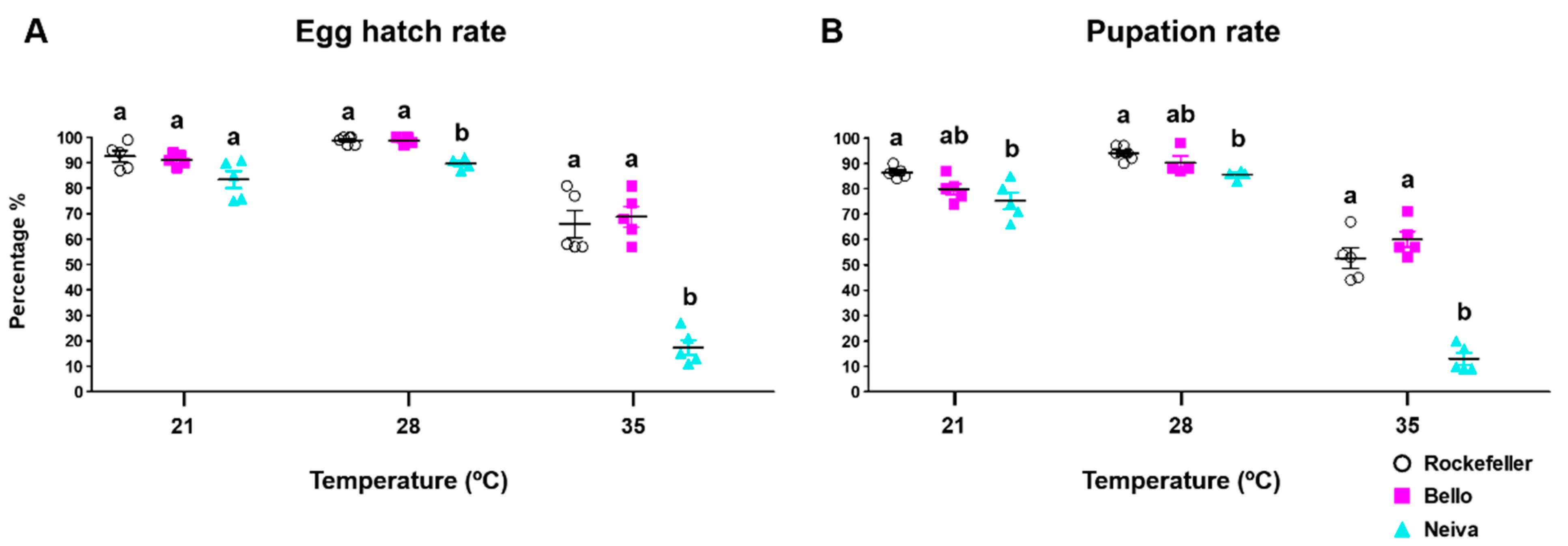
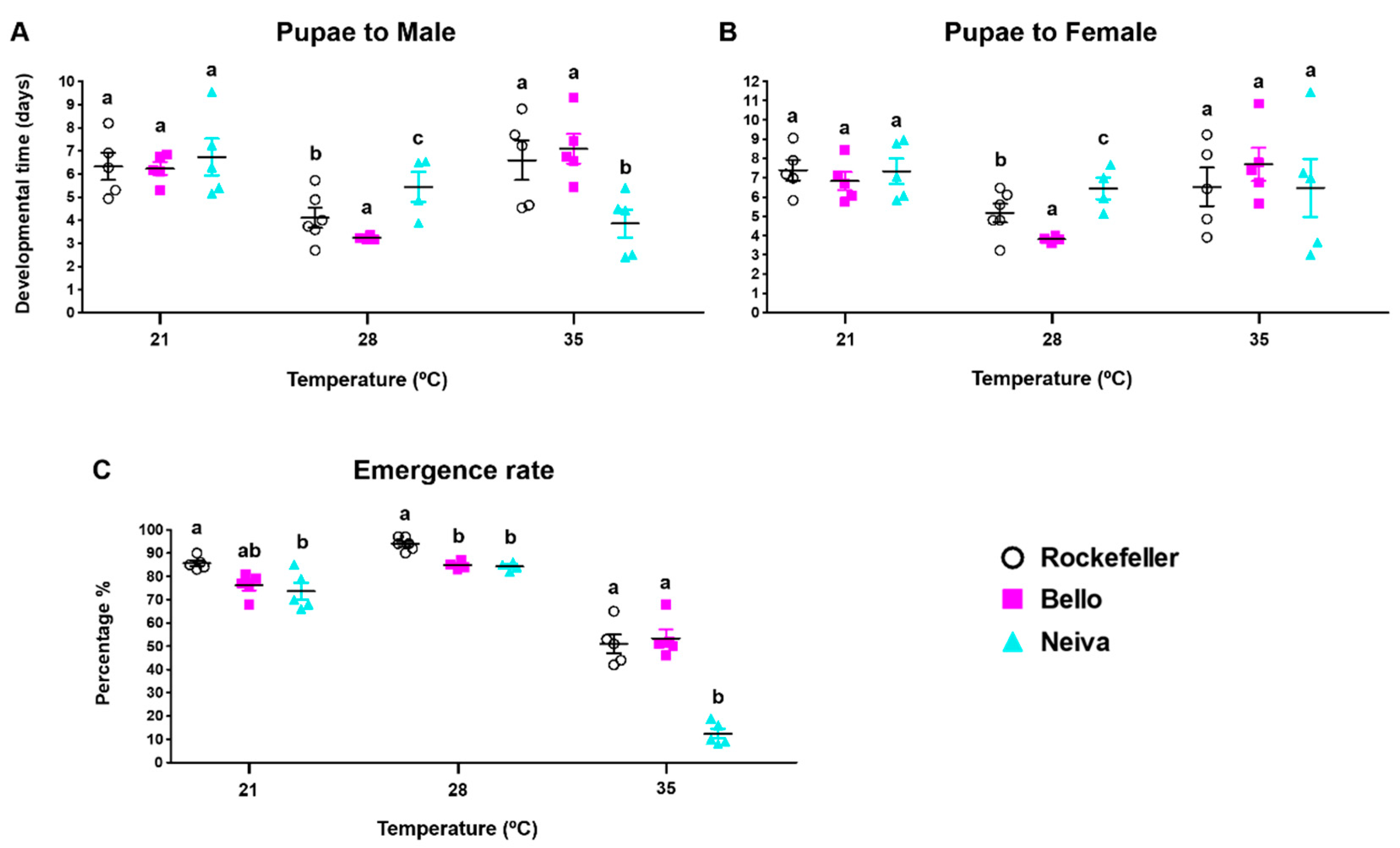
3.5. Effect of Temperature on Immature Stage Development Times, Egg Hatching, Pupation, and Emergence Rates for Bello and Neiva
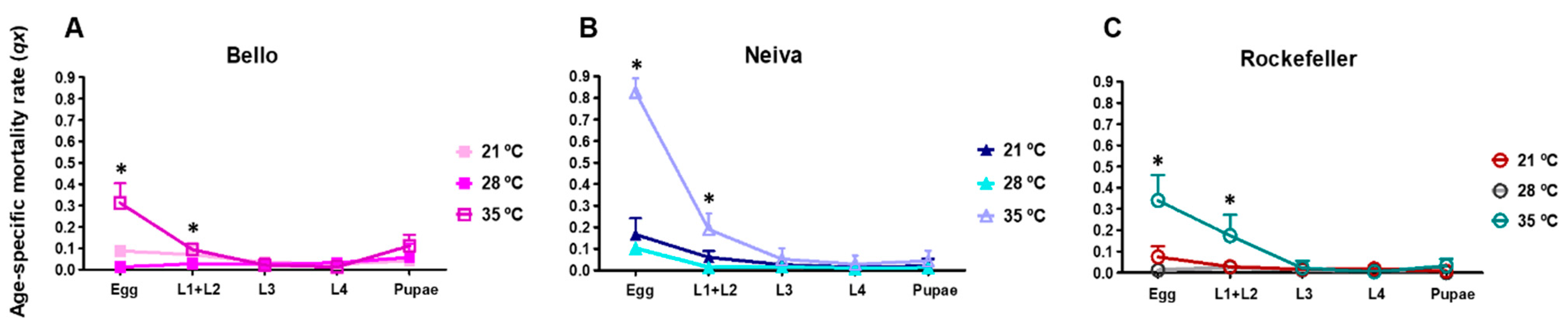
3.6. Effect of Temperature on Mortality in Immature Stages from Bello and Neiva
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simmons, C.P.; Farrar, J.J.; van Vinh Chau, N.; Wills, B. Dengue. N. Engl. J. Med. 2012, 366, 1423–1432. [Google Scholar] [CrossRef]
- Jansen, C.C.; Beebe, N.W. The dengue vector Aedes aegypti: What comes next. Microbes Infect. 2010, 12, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Chouin-Carneiro, T.; Vega-Rua, A.; Vazeille, M.; Yebakima, A.; Girod, R.; Goindin, D.; Dupont-Rouzeyrol, M.; Lourenco-de-Oliveira, R.; Failloux, A.B. Differential susceptibilities of Aedes aegypti and Aedes albopictus from the Americas to Zika virus. PLoS Negl. Trop. Dis. 2016, 10, e0004543. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhou, T.; Lai, Z.; Zhang, Z.; Jia, Z.; Zhou, G.; Williams, T.; Xu, J.; Gu, J.; Zhou, X.; et al. Competence of Aedes aegypti, Ae. albopictus, and Culex quinquefasciatus mosquitoes as Zika virus vectors, China. Emerg. Infect. Dis. 2017, 23, 1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahid, B.; Ali, A.; Rafique, S.; Idrees, M. Global expansion of chikungunya virus: Mapping the 64-year history. Int. J. Infect. Dis. 2017, 58, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Burt, F.J.; Rolph, M.S.; Rulli, N.E.; Mahalingam, S.; Heise, M.T. Chikungunya: A re-emerging virus. Lancet 2012, 379, 662–671. [Google Scholar] [CrossRef]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- PAHO/WHO. Epidemiological Update: Dengue; Pan American Health Organization: Washington, DC, USA, 7 February 2020; pp. 1–21. [Google Scholar]
- PAHO/WHO. Epidemiological Update: Dengue and other Arboviruses; Pan American Health Organization: Washington, DC, USA, 10 June 2020; pp. 1–15. [Google Scholar]
- Boeuf, P.; Drummer, H.; Richards, J.S.; Scoullar, M.; Beeson, J.G. The global threat of Zika virus to pregnancy: Epidemiology, clinical perspectives, mechanisms, and impact. BMC Med. 2016, 14, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Instituto Nacional de Salud. Boletín Epidemiológico Semanal (BES). Semana Epidemiológica Número 10 de 2016. BES-Sivigila. 2016. Available online: https://www.ins.gov.co/buscador-eventos/Paginas/Vista-Boletin-Epidemilogico.aspx (accessed on 15 April 2022).
- Instituto Nacional de Salud. Boletín Epidemiológico Semanal (BES). Semana Epidemiológica Número 52 de 2016. BES-Sivigila. 2016. Available online: https://www.ins.gov.co/buscador-eventos/Paginas/Vista-Boletin-Epidemilogico.aspx (accessed on 15 April 2022).
- Granada, Y.; Mejía-Jaramillo, A.M.; Strode, C.; Triana-Chavez, O. A Point Mutation V419L in the Sodium Channel Gene from Natural Populations of Aedes aegypti Is Involved in Resistance to λ-Cyhalothrin in Colombia. Insects 2018, 9, 23. [Google Scholar] [CrossRef] [Green Version]
- Serrano, R.M.; Gomez-Camargo, D.; Ponce-Garcia, G.; Flores, A.E. Susceptibility to insecticides and resistance mechanisms in Aedes aegypti from the Colombian Caribbean Region. Pestic. Biochem. Physiol. 2014, 116, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Aponte, A.; Penilla, R.P.; Rodríguez, A.D.; Ocampo, C.B. Mechanisms of pyrethroid resistance in Aedes (Stegomyia) aegypti from Colombia. Acta Trop. 2018, 191, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, L.C.; Ponce, G.; Saavedra-Rodriguez, K.; Lopez, B.; Flores, A.E. Frequency of V1016I and F1534C mutations in the voltage-gated sodium channel gene in Aedes aegypti in Venezuela. Pest Manag. Sci. 2015, 71, 863–869. [Google Scholar] [CrossRef]
- Linss, J.G.B.; Brito, L.P.; Garcia, G.A.; Araki, A.S.; Bruno, R.V.; Lima, J.B.P.; Valle, D.; Martins, A.J. Distribution and dissemination of the Val1016Ile and Phe1534Cys Kdr mutations in Aedes aegypti Brazilian natural populations. Parasites Vectors 2014, 7, 25. [Google Scholar] [CrossRef] [Green Version]
- Ahsan, M.T.; Ahmad, I.; Shoaib, M. Horizontal and vertical life tables and survivorship curves of cotton leafhopper Amrasca devastans (Distant) (Homoptera: Cicadellidae) and the distribution of mortality with age. Int. J. Biol. Res. 2014, 2, 97–100. [Google Scholar]
- Wang, X.; Zhou, G.; Zhong, D.; Wang, Y.; Yang, Z.; Cui, L.; Yan, G.; Wang, X. Life-table studies revealed significant effects of deforestation on the development and survivorship of Anopheles minimus larvae. Parasites Vectors 2016, 9, 323. [Google Scholar] [CrossRef] [Green Version]
- Okogun, G.R.A. Life-table analysis of Anopheles malaria vectors: Generational mortality as tool in mosquito vector abundance and control studies. J. Vector Borne Dis. 2005, 42, 45–53. [Google Scholar]
- Olayemi, I.K.; Ande, A.T. Life table analysis of Anopheles gambiae (diptera: Culicidae) in relation to malaria transmission. J. Vector Borne Dis. 2009, 46, 295–298. [Google Scholar] [PubMed]
- Quispe, E.; Carbajal, A.; Gozzer, J.; Moreno, B. Ciclo biológico y Tabla de Vida de Aedes aegypti, en laboratorio: Trujillo (Perú), 2014. Rev. Rebiolest 2015, 3, 91–101. [Google Scholar]
- Löwenberg Neto, P.; Navarro-Silva, M.A. Development, longevity, gonotrophic cycle and oviposition of Aedes albopictus Skuse (Diptera: Culicidae) under cyclic temperatures. Neotrop. Entomol. 2004, 33, 29–33. [Google Scholar] [CrossRef] [Green Version]
- De Majo, M.S.; Montini, P.; Fischer, S. Egg Hatching and Survival of Immature Stages of Aedes aegypti (Diptera: Culicidae) Under Natural Temperature Conditions During the Cold Season in Buenos Aires, Argentina. J. Med. Entomol. 2016, 54, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Chubachi, R. An Analysis of the Generation-Mean Life Table of the Mosquito, Culex tritaeniorhynchus summorosus, with Particular Reference to Population Regulation. J. Anim. Ecol. 1979, 48, 681. [Google Scholar] [CrossRef]
- Abouzied, E.M. Life Table Analysis of Culex pipiens Under Simulated Weather Conditions In Egypt. J. Am. Mosq. Control Assoc. 2017, 33, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Arévalo, A.; Carranza, J.C.; Guhl, F.; Clavijo, J.A.; Vallejo, G.A. Comparación del ciclo de vida de Rhodnius colombiensis Moreno, Jurberg & Galvão, 1999 y Rhodnius prolixus Stal, 1872 (Hemiptera, Reduviidae, Triatominae) en condiciones de laboratorio. Biomédica 1999, 27, 119–129. [Google Scholar]
- Villacís, A.G.; Arcos-Terán, L.; Grijalva, M.J. Life cycle, feeding and defecation patterns of Rhodnius ecuadoriensis (Lent & León 1958) (Hemiptera: Reduviidae: Triatominae) under laboratory conditions. Mem. Do Inst. Oswaldo Cruz 2008, 103, 690–695. [Google Scholar]
- Williams, B.G.; Dransfield, R.D.; Brightwell, R. Tsetse fly (Diptera: Glossinidae) population dynamics and the estimation of mortality rates from life-table data. Bull. Èntomol. Res. 1990, 80, 479–485. [Google Scholar] [CrossRef]
- Hargrove, J.; Vale, G. Models for the rates of pupal development, fat consumption and mortality in tsetse (Glossina spp.). Bull. Èntomol. Res. 2019, 110, 44–56. [Google Scholar] [CrossRef]
- Ferro, C.; Cárdenas, E.; Corredor, D.; Morales, A.; Munstermann, L.E. Life Cycle and Fecundity Analysis of Lutzomyia shannoni (Dyar) (Diptera: Psychodidae). Mem. Do Inst. Oswaldo Cruz 1998, 93, 195–199. [Google Scholar] [CrossRef] [Green Version]
- Escovar, J.; Bello, F.J.; Morales, A.; Moncada, L.; Cárdenas, E. Life tables and reproductive parameters of Lutzomyia spinicrassa (Diptera: Psychodidae) under laboratory conditions. Mem. Do Inst. Oswaldo Cruz 2004, 99, 603–607. [Google Scholar] [CrossRef] [Green Version]
- Legros, M.; Otero, M.; Aznar, V.R.; Solari, H.; Gould, F.; Lloyd, A.L. Comparison of two detailed models of Aedes aegypti population dynamics. Ecosphere 2016, 7, e01515. [Google Scholar] [CrossRef]
- Healy, K.B.; Dugas, E.; Fonseca, D.M. Development of a Degree-Day Model to Predict Egg Hatch of Aedes albopictus. J. Am. Mosq. Control Assoc. 2019, 35, 249–257. [Google Scholar] [CrossRef] [Green Version]
- Christiansen-Jucht, C.; Erguler, K.; Shek, C.Y.; Basáñez, M.-G.; Parham, P.E. Modelling Anopheles gambiae s.s. Population Dynamics with Temperature- and Age-Dependent Survival. Int. J. Environ. Res. Public Health 2015, 12, 5975–6005. [Google Scholar] [CrossRef] [Green Version]
- Jorge, M.R.; de Souza, A.P.; dos Passos, R.A.; Martelli, S.M.; Rech, C.R.; Barufatti, A.; do Amaral Crispim, B.; dos Santos Nascimento, H.; de Arruda, E.J. The Yellow Fever Mosquito Aedes aegypti (Linnaeus): The Breeding Sites. In Life Cycle and Development of Diptera; Sarwar, M., Ed.; IntechOpen: London, UK, 2020; Available online: https://www.intechopen.com/books/life-cycle-and-development-of-diptera/the-yellow-fever-mosquito-em-aedes-aegypti-em-linnaeus-the-breeding-sites (accessed on 21 October 2020).
- Castro, F.P.D., Jr.; Martins, W.F.; Lucena Filho, M.L.D.; Almeida, R.P.D.; Beserra, E.B. Ciclos de vida comparados de Aedes aegypti (Diptera, Culicidae) do semiárido da Paraíba. Iheringia Série Zool. 2013, 103, 118–123. [Google Scholar] [CrossRef]
- Beserra, E.B.; Castro, F.P.D., Jr. Biologia comparada de populações de Aedes (Stegomyia) aegypti (L.) (Diptera: Culicidae) da Paraíba. Neotrop. Entomol. 2018, 37, 81–85. [Google Scholar] [CrossRef] [Green Version]
- Beserra, E.B.; Castro, F.P.D., Jr.; Santos, J.W.D.; Santos, T.D.S.; Fernandes, C.R. Biologia e exigências térmicas de Aedes aegypti (L.)(Diptera: Culicidae) provenientes de quatro regiões bioclimáticas da Paraíba. Neotrop. Entomol. 2006, 35, 853–860. [Google Scholar] [CrossRef] [Green Version]
- Beserra, E.B.; Fernandes, C.R.M.; Ribeiro, P.S. Relação entre densidade larval e ciclo de vida, tamanho e fecundidade de Aedes (Stegomyia) aegypti (L.) (Diptera: Culicidae) em laboratório. Neotropical Èntomol. 2009, 38, 847–852. [Google Scholar] [CrossRef]
- Beserra, E.B.; Fernandes, C.R.; Silva, S.A.D.O.; Silva, L.A.D.; Santos, J.W.D. Efeitos da temperatura no ciclo de vida, exigências térmicas e estimativas do número de gerações anuais de Aedes aegypti (Diptera, Culicidae). Iheringia Série Zool. 2009, 99, 142–148. [Google Scholar] [CrossRef] [Green Version]
- Suman, D.S.; Tikar, S.N.; Mendki, M.J.; Sukumaran, D.; Agrawal, O.P.; Parashar, B.D.; Prakash, S. Variations in life tables of geographically isolated strains of the mosquito Culex quinquefasciatus. Med. Veter. Èntomol. 2011, 25, 276–288. [Google Scholar] [CrossRef]
- Paduan, K.D.S.; Araújo-Júnior, J.P.; Ribolla, P.E. Genetic variability in geographical populations of Aedes aegypti (Diptera, Culicidae) in Brazil elucidated by molecular markers. Genet. Mol. Biol. 2006, 29, 391–395. [Google Scholar] [CrossRef] [Green Version]
- Jaimes-Dueñez, J.; Arboleda, S.; Triana-Chávez, O.; Gómez-Palacio, A. Spatio-Temporal Distribution of Aedes aegypti (Diptera: Culicidae) Mitochondrial Lineages in Cities with Distinct Dengue Incidence Rates Suggests Complex Population Dynamics of the Dengue Vector in Colombia. PLoS Negl. Trop. Dis. 2015, 9, e0003553. [Google Scholar] [CrossRef]
- Atencia, M.C.; Pérez, M.D.J.; Caldera, S.M.; Jaramillo, M.C.; Bejarano, E.E. Variabilidad genética de Aedes aegypti en el departamento de Sucre, Colombia, mediante el análisis de la secuencia de nucleótidos del gen mitocondrial ND4. Biomédica 2018, 38, 267–276. [Google Scholar] [CrossRef]
- Mitchell-Foster, K.; Ma, B.; Warsame-Ali, S.; Logan, C.; Rau, M.E.; Lowenberger, C. The influence of larval density, food stress, and parasitism on the bionomics of the dengue vector Aedes aegypti (Diptera: Culicidae): Implications for integrated vector management. J. Vector Ecol. 2012, 37, 221–229. [Google Scholar] [CrossRef]
- Couret, J.; Dotson, E.; Benedict, M.Q. Temperature, larval diet, and density effects on development rate and survival of Aedes aegypti (Diptera: Culicidae). PLoS ONE 2014, 9, e87468. [Google Scholar]
- Dickson, L.B.; Jiolle, D.; Minard, G.; Moltini-Conclois, I.; Volant, S.; Ghozlane, A.; Bouchier, C.; Ayala, D.; Paupy, C.; Moro, C.V.; et al. Carryover effects of larval exposure to different environmental bacteria drive adult trait variation in a mosquito vector. Sci. Adv. 2017, 3, e1700585. [Google Scholar] [CrossRef] [Green Version]
- Coon, K.L.; Vogel, K.J.; Brown, M.R.; Strand, M.R. Mosquitoes rely on their gut microbiota for development. Mol. Ecol. 2014, 23, 2727–2739. [Google Scholar] [CrossRef] [Green Version]
- Coon, K.L.; Brown, M.R.; Strand, M.R. Mosquitoes host communities of bacteria that are essential for development but vary greatly between local habitats. Mol. Ecol. 2016, 25, 5806–5826. [Google Scholar] [CrossRef] [Green Version]
- Travanty, N.V.; Apperson, C.S.; Ponnusamy, L. A Diverse Microbial Community Supports Larval Development and Survivorship of the Asian Tiger Mosquito (Diptera: Culicidae). J. Med. Èntomol. 2019, 56, 632–640. [Google Scholar] [CrossRef]
- Marinho, R.A.; Beserra, E.B.; Bezerra-Gusmão, M.A.; Porto, V.D.S.; Olinda, R.A.; Dos Santos, C.A. Effects of temperature on the life cycle, expansion, and dispersion of Aedes aegypti (Diptera: Culicidae) in three cities in Paraiba, Brazil. J. Vector Ecol. 2016, 41, 1–10. [Google Scholar] [CrossRef]
- Christiansen-Jucht, C.D.; Parham, P.E.; Saddler, A.; Koella, J.C.; Basañez, M.-G. Larval and adult environmental temperatures influence the adult reproductive traits of Anopheles gambiae s.s. Parasites Vectors 2015, 8, 456. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, A.; Chadee, D.D. Effects of different temperature regimens on the development of Aedes aegypti (L.) (Diptera: Culicidae) mosquitoes. Acta Trop. 2011, 119, 38–43. [Google Scholar] [CrossRef]
- Clark, T.M.; Flis, B.J.; Remold, S.K. pH tolerances and regulatory abilities of freshwater and euryhaline Aedine mosquito larvae. J. Exp. Biol. 2004, 207, 2297–2304. [Google Scholar] [CrossRef] [Green Version]
- Owiti, Y.J.; Misire, C. Effect of temperature and pH on egg viability and pupation of Anopheles arabiensis Patton (Diptera: Culicidae): Prospect for optimizing colony reproduction procedures. Jordan J. Biol. Sci. 2017, 10, 7–12. [Google Scholar]
- Oliver, S.V.; Brooke, B.D. The effect of elevated temperatures on the life history and insecticide resistance phenotype of the major malaria vector Anopheles arabiensis (Diptera: Culicidae). Malar. J. 2017, 16, 73. [Google Scholar] [CrossRef] [Green Version]
- Msangi, G.; Olotu, M.I.; Mahande, A.M.; Philbert, A.; Kweka, E.J.; Dash, A.P. The Impact of Insecticide Pre-Exposure on Longevity, Feeding Succession, and Egg Batch Size of Wild Anopheles gambiae s.l. J. Trop. Med. 2020, 2020, 1–8. [Google Scholar] [CrossRef]
- Martins, A.J.; Ribeiro, C.D.E.M.; Bellinato, D.F.; Peixoto, A.A.; Valle, D.; Lima, J.B.P. Effect of insecticide resistance on development, longevity and reproduction of field or laboratory selected Aedes aegypti populations. PLoS ONE 2012, 7, e31889. [Google Scholar]
- Belinato, T.A.; Martins, A.J.; Valle, D. Fitness evaluation of two Brazilian Aedes aegypti field populations with distinct levels of resistance to the organophosphate temephos. Mem. Do Inst. Oswaldo Cruz 2012, 107, 916–922. [Google Scholar] [CrossRef] [Green Version]
- Owusu, H.F.; Chitnis, N.; Müller, P. Insecticide susceptibility of Anopheles mosquitoes changes in response to variations in the larval environment. Sci. Rep. 2017, 7, 3667. [Google Scholar] [CrossRef] [Green Version]
- Zeller, M.; Koella, J.C. Effects of food variability on growth and reproduction of Aedes aegypti. Ecol. Evol. 2016, 6, 552–559. [Google Scholar] [CrossRef]
- IDEAM. Atlas Climatológico de Colombia. Instituto de Hidrología, Meteorología y Estudios Ambientales. Gobierno de Colombia. 2010. Available online: http://atlas.ideam.gov.co/visorAtlasClimatologico.html (accessed on 29 October 2021).
- Benítez-Díaz, L.; Diaz-Quijano, F.A.; Martínez-Vega, R.A. Experiencia y percepción del riesgo asociados a conocimientos, actitudes y prácticas sobre dengue en Riohacha, Colombia. Ciência Saúde Coletiva 2020, 25, 1137–1146. [Google Scholar] [CrossRef]
- Gobernación de Antioquia. Climatología Registrada en Algunas Estaciones de Hidrología, Meteorología y Estudio Ambiental en Algunos Municipios de Antioquia, Por Meses. Medio Ambiente. Hidrometeorología. Anuario estadístico de Antioquia. 2016. Available online: http://www.antioquiadatos.gov.co/index.php/2-2-3-climatologia-registrada-en-algunas-estaciones-de-hidrologia-meteorologia-y-estudio-ambiental-en-algunos-municipios-de-antioquia-por-meses-2016 (accessed on 21 November 2021).
- Secretaría Seccional de Salud de Antioquia. Eventos de Interés en Salud Pública por Subregiones y Municipios. Secretaría Seccional de Salud y Proteción Social de Antioquia. Gobernación de Antioquia. 2019. Available online: https://www.dssa.gov.co/index.php/estadisticas/eventos-en-salud-publica/item/1304-eventos-de-interes-en-salud-publica-antioquia-2019 (accessed on 21 November 2021).
- Secretaría de Salud de Neiva. Boletín Epidemiológico de Dengue; Semanas Epidemiológicas No. 1 a la 52 de 2019; Alcaldía de Neiva; Secretaría de Salud Municipa; Equipo de Vigilancia Epidemiológica Municipal: Cidra, PR, USA, 31 December 2019.
- Kuno, G. Early history of laboratory breeding of Aedes aegypti (Diptera: Culicidae) focusing on the origins and use of selected strains. J. Med. Èntomol. 2010, 47, 957–971. [Google Scholar] [CrossRef] [Green Version]
- Alvarado, W.A.; Agudelo, S.O.; Velez, I.D.; Vivero, R.J. Description of the ovarian microbiota of Aedes aegypti (L) Rockefeller strain. Acta Trop. 2020, 214, 105765. [Google Scholar] [CrossRef]
- Brito, L.P.; Linss, J.G.B.; Lima-Camara, T.N.; Belinato, T.A.; Peixoto, A.A.; Lima, J.B.P.; Valle, D.; Martins, A.J. Assessing the Effects of Aedes aegypti kdr Mutations on Pyrethroid Resistance and Its Fitness Cost. PLoS ONE 2013, 8, e60878. [Google Scholar] [CrossRef] [PubMed]
- Rueda, L.M. Pictorial keys for the identification of mosquitoes (Diptera: Culicidae) associated with Dengue Virus Transmission. Zootaxa 2004, 589, 1–60. [Google Scholar] [CrossRef]
- Christophers, S.R. Aedes aegypti (L.) the Yellow Fever Mosquito: Its Life History, Bionomics and Structure; Cambridge University Press: Cambridge, UK, 1960. [Google Scholar]
- Bargielowski, I.; Nimmo, D.; Alphey, L.; Koella, J.C. Comparison of Life History Characteristics of the Genetically Modified OX513A Line and a Wild Type Strain of Aedes aegypti. PLoS ONE 2011, 6, e20699. [Google Scholar] [CrossRef] [Green Version]
- Zapletal, J.; Erraguntla, M.; Adelman, Z.N.; Myles, K.M.; Lawley, M.A. Impacts of diurnal temperature and larval density on aquatic development of Aedes aegypti. PLoS ONE 2018, 13, e0194025. [Google Scholar] [CrossRef] [Green Version]
- Bond, J.G.; Ramírez-Osorio, A.; Marina, C.F.; Fernández, C.F.M.; Liedo, P.; Dor, A.; Williams, T. Efficiency of two larval diets for mass-rearing of the mosquito Aedes aegypti. PLoS ONE 2017, 12, e0187420. [Google Scholar] [CrossRef] [Green Version]
- Araujo, M.D.S.; Gil, L.H.S. Larval food quantity affects development time, survival and adult biological traits that influence the vectorial capacity of Anopheles darlingi under laboratory conditions. Malar. J. 2012, 11, 261. [Google Scholar] [CrossRef] [Green Version]
- Sowilem, M.M.; A Kamal, H.; Khater, E.I. Life table characteristics of Aedes aegypti (Diptera:Culicidae) from Saudi Arabia. Trop. Biomed. 2013, 30, 301–314. [Google Scholar]
- Reinhold, J.; Lazzari, C.; Lahondère, C. Effects of the Environmental Temperature on Aedes aegypti and Aedes albopictus Mosquitoes: A Review. Insects 2018, 9, 158. [Google Scholar] [CrossRef] [Green Version]
- Mazigo, E.; Kidima, W.; Myamba, J.; Kweka, E.J. The impact of Anopheles gambiae egg storage for mass rearing and production success. Malar. J. 2019, 18, 52. [Google Scholar] [CrossRef] [Green Version]
- Soma, D.D.; Maïga, H.; Mamai, W.; Bimbile-Somda, N.S.; Venter, N.; Ali, A.B.; Yamada, H.; Diabaté, A.; Fournet, F.; Ouédraogo, G.A.; et al. Does mosquito mass-rearing produce an inferior mosquito? Malar. J. 2017, 16, 357. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; He, Y.; Ni, W.; Lai, Q.; Yang, Y.; Xie, J.; Zhu, T.; Zhou, G.; Zheng, X. Semi-field life-table studies of Aedes albopictus (Diptera: Culicidae) in Guangzhou, China. PLoS ONE 2020, 15, e0229829. [Google Scholar] [CrossRef] [PubMed]
- Sylvestre, G.; Gandini, M.; Maciel-De-Freitas, R. Age-Dependent Effects of Oral Infection with Dengue Virus on Aedes aegypti (Diptera: Culicidae) Feeding Behavior, Survival, Oviposition Success and Fecundity. PLoS ONE 2013, 8, e59933. [Google Scholar] [CrossRef] [PubMed]
- Rabinovich, J. Introducción a La Ecología de Poblaciones Animales, 2nd ed.; Jorge, E., Ed.; Continental S.A.: San Luis Potosi, México, 1980; p. 313. [Google Scholar]
- Begon, M.; Townsend, C.R.; Harper, J.L. Ecology: From Individuals to Ecosystems, 4th ed.; Blackwell Publisher: Malden, MA, USA, 2006; p. 738. [Google Scholar]
- Keyfitz, N.; Caswell, H. (Eds.) The Life Table. En: Applied Mathematical Demography; Springer: New York, NY, USA, 2005; pp. 29–47. [Google Scholar] [CrossRef]
- Reisen, W.K.; Mahmood, F. Horizontal Life Table Characteristics of the Malaria Vectors Anopheles Culicifacies and Anopheles Stephensi (Diptera: Culicidae)1. J. Med Èntomol. 1980, 17, 211–217. [Google Scholar] [CrossRef]
- Richards, S.L.; Anderson, S.L.; Yost, S.A. Effects of blood meal source on the reproduction of Culex pipiens quinquefasciatus (Diptera: Culicidae). J. Vector Ecol. 2012, 37, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Phasomkusolsil, S.; Tawong, J.; Monkanna, N.; Pantuwatana, K.; Damdangdee, N.; Khongtak, W.; Kertmanee, Y.; Evans, B.P.; Schuster, A.L. Maintenance of mosquito vectors: Effects of blood source on feeding, survival, fecundity, and egg hatching rates. J. Vector Ecol. 2013, 38, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Prasadini, M.; Dayananda, D.; Fernando, S.; Harischandra, I.; De Silva, N. Blood Feeding Preference of Female Aedes aegypti Mosquitoes for Human Blood Group Types and Its Impact on Their Fecundity: Implications for Vector Control. Am. J. Èntomol. 2019, 3, 43. [Google Scholar] [CrossRef]
- Khan, S.A.; Ombugadu, A.; Ahmad, S. Host-seeking behavior and fecundity of the female Aedes aegypti to human blood types. Pest Manag. Sci. 2021, 78, 321–328. [Google Scholar] [CrossRef]
- Crawford, J.; Alves, J.; Palmer, W.J.; Day, J.P.; Sylla, M.; Ramasamy, R.; Surendran, S.N.; Iv, W.C.B.; Pain, A.; Jiggins, F.M. Population genomics reveals that an anthropophilic population of Aedes aegypti mosquitoes in West Africa recently gave rise to American and Asian populations of this major disease vector. BMC Biol. 2017, 15, 16. [Google Scholar] [CrossRef] [Green Version]
- Ayala-Sulca, Y.O.; Colos-Galindo, P.; Portal-Quicaña, E.; Ibarra-Juárez, L.; Cóndor-Alarcón, R.; Carrasco-Badajoz, C.; Ramírez, R. Respuesta biológica cuantitativa de dos depredadores (Heteroptera: Notonectidae) en el control larval de Aedes aegypti (Diptera: Culicidae). Rev. Colomb. De Entomol. 2021, 47, e10535. [Google Scholar] [CrossRef]
- Weterings, R.; Umponstira, C.; Buckley, H.L. Landscape variation influences trophic cascades in dengue vector food webs. Sci. Adv. 2018, 4, eaap9534. [Google Scholar] [CrossRef] [Green Version]
- Roux, O.; Vantaux, A.; Roche, B.; Yameogo, K.B.; Dabiré, K.R.; Diabaté, A.; Simard, F.; Lefèvre, T. Evidence for carry-over effects of predator exposure on pathogen transmission potential. Proc. R. Soc. B Biol. Sci. 2015, 282, 20152430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tejerina, E.F.; Almeida, F.F.L.; Almirón, W.R. Bionomics of Aedes aegypti subpopulations (Diptera: Culicidae) from Misiones Province, northeastern Argentina. Acta Trop. 2009, 109, 45–49. [Google Scholar] [CrossRef]
- Arévalo-Cortés, A.; Mejia-Jaramillo, A.M.; Granada, Y.; Coatsworth, H.; Lowenberger, C.; Triana-Chavez, O. The Midgut Microbiota of Colombian Aedes aegypti Populations with Different Levels of Resistance to the Insecticide Lambda-cyhalothrin. Insects 2020, 11, 584. [Google Scholar] [CrossRef] [PubMed]
- Peña-García, V.H.; Triana-Chávez, O.; Arboleda-Sánchez, S. Estimating Effects of Temperature on Dengue Transmission in Colombian Cities. Ann. Glob. Health 2017, 83, 509. [Google Scholar] [CrossRef]
- Brady, O.J.; Golding, N.; Pigott, D.M.; Kraemer, M.U.G.; Messina, J.P.; Reiner, R.C.; Scott, T.W.; Smith, D.L.; Gething, P.W.; Hay, S.I. Global temperature constraints on Aedes aegypti and Ae. albopictus persistence and competence for dengue virus transmission. Parasit Vectors 2014, 7, 338. [Google Scholar] [CrossRef] [Green Version]
- Mayton, E.H.; Tramonte, A.R.; Wearing, H.J.; Christofferson, R.C. Age-structured vectorial capacity reveals timing, not magnitude of within-mosquito dynamics is critical for arbovirus fitness assessment. Parasites Vectors 2020, 13, 310. [Google Scholar] [CrossRef]
- Dye, C. Vectorial capacity: Must we measure all its components? Parasitol. Today 1986, 2, 203–209. [Google Scholar] [CrossRef]
- Chadee, D.D.; Martinez, R.; Sutherland, J.M. Aedes aegypti (L.) mosquitoes in Trinidad, West Indies: Longevity case studies. J. Vector Ecol. 2017, 42, 130–135. [Google Scholar] [CrossRef] [Green Version]
- Maciel-De-Freitas, R.; Koella, J.; Lourenço-De-Oliveira, R. Lower survival rate, longevity and fecundity of Aedes aegypti (Diptera: Culicidae) females orally challenged with dengue virus serotype 2. Trans. R. Soc. Trop. Med. Hyg. 2011, 105, 452–458. [Google Scholar] [CrossRef]
- Goindin, D.; Delannay, C.; Ramdini, C.; Gustave, J.; Fouque, F. Parity and Longevity of Aedes aegypti According to Temperatures in Controlled Conditions and Consequences on Dengue Transmission Risks. PLoS ONE 2015, 10, e0135489. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Cui, M.; Xia, L.; Yu, Q.; Cao, Y.; Wu, Y. Age-Stage, Two-Sex Life Tables of the Predatory Mite Cheyletus Malaccensis Oudemans at Different Temperatures. Insects 2020, 11, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrero, M.I.; Dami, L.C.; Fogliata, S.V.; Casmuz, A.S.; Gómez, D.R.S.; Gastaminza, G.A.; Murúa, M.G. Fertility Life Table, Population Parameters and Biotic Potential of Helicoverpa gelotopoeon (Dyar) (Lepidoptera: Noctuidae). An. Da Acad. Bras. De Ciências 2018, 90, 3831–3838. [Google Scholar] [CrossRef] [PubMed]
- Supriya, G.; Singh, T.; Sunitha, V.; Vinod, S. Age-specific life table, fecundity table, life parameters, stable age distribution and life expectancy of Spodoptera litura (F.) on cotton hybrids during 90–120 DAS. J. Entomol. Zool. Stud. 2018, 6, 477–486. [Google Scholar]
- Maimusa, H.A.; Ahmad, A.H.; Kassim, N.F.A.; Rahim, J. Age-Stage, Two-Sex Life Table Characteristics of Aedes albopictus and Aedes aegypti in Penang Island, Malaysia. J. Am. Mosq. Control. Assoc. 2016, 32, 1–11. [Google Scholar] [CrossRef]
- Ponnusamy, L.; Böröczky, K.; Wesson, D.M.; Schal, C.; Apperson, C.S. Bacteria Stimulate Hatching of Yellow Fever Mosquito Eggs. PLoS ONE 2011, 6, e24409. [Google Scholar] [CrossRef]
- Mosquera, K.D.; Martinez Villegas, L.E.; Pidot, S.J.; Sharif, C.; Klimpel, S.; Stinear, T.P.; Moreira, L.A.; Tobias, N.J.; Lorenzo, M.G. Multi-Omic Analysis of Symbiotic Bacteria Associated With Aedes aegypti Breeding Sites. Front. Microbiol. 2021, 12, 703711. [Google Scholar] [CrossRef]
- Farnesi, L.C.; Belinato, T.A.; Gesto, J.S.M.; Martins, A.J.; Bruno, R.V.; Moreira, L.A. Embryonic development and egg viability of w Mel-infected Aedes aegypti. Parasites Vectors 2019, 12, 211. [Google Scholar] [CrossRef]
- McMeniman, C.J.; O’Neill, S.L. A Virulent Wolbachia Infection Decreases the Viability of the Dengue Vector Aedes aegypti during Periods of Embryonic Quiescence. PLoS Negl. Trop. Dis. 2010, 4, e748. [Google Scholar] [CrossRef] [Green Version]
- Yeap, H.L.; Mee, P.; Walker, T.; Weeks, A.R.; O’Neill, S.L.; Johnson, P.; Ritchie, S.A.; Richardson, K.M.; Doig, C.; Endersby, N.M.; et al. Dynamics of the “popcorn” Wolbachia infection in outbred Aedes aegypti informs prospects for mosquito vector control. Genetics 2011, 187, 583–595. [Google Scholar] [CrossRef] [Green Version]
- Coon, K.L.; Brown, M.R.; Strand, M.R. Gut bacteria differentially affect egg production in the anautogenous mosquito Aedes aegypti and facultatively autogenous mosquito Aedes atropalpus (Diptera: Culicidae). Parasites Vectors 2016, 9, 375. [Google Scholar] [CrossRef]
- Gnambani, E.J.; Bilgo, E.; Sanou, A.; Dabiré, R.K.; Diabaté, A. Infection of highly insecticide-resistant malaria vector Anopheles coluzzii with entomopathogenic bacteria Chromobacterium violaceum reduces its survival, blood feeding propensity and fecundity. Malar. J. 2020, 19, 352. [Google Scholar] [CrossRef] [PubMed]
- Correa, M.A.; Matusovsky, B.; Brackney, D.E.; Steven, B. Generation of axenic Aedes aegypti demonstrate live bacteria are not required for mosquito development. Nat. Commun. 2018, 9, 4464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, R.S.; Virginio, F.; Riback, T.I.S.; Suesdek, L.; Barufi, J.B.; Genta, F.A. Microorganism-based larval diets affect mosquito development, size and nutritional reserves in the yellow fever mosquito Aedes aegypti (Diptera: Culicidae). Front. Physiol. 2019, 10, 152. [Google Scholar] [CrossRef]
- Velez, I.D.; Santacruz, E.; Kutcher, S.C.; Duque, S.L.; Uribe, A.; Barajas, J.; Gonzalez, S.; Patino, A.C.; Zuluaga, L.; Martínez, L.; et al. The impact of city-wide deployment of Wolbachia-carrying mosquitoes on arboviral disease incidence in Medellín and Bello, Colombia: Study protocol for an interrupted time-series analysis and a test-negative design study. F1000Research 2019, 8, 1327. [Google Scholar] [CrossRef] [Green Version]
- Granada, Y.; Mejía-Jaramillo, A.M.; Zuluaga, S.; Triana-Chávez, O. Molecular surveillance of resistance to pyrethroids insecticides in Colombian Aedes aegypti populations. PLoS Negl. Trop. Dis. 2021, 15, e0010001. [Google Scholar] [CrossRef]
- Saingamsook, J.; Yanola, J.; Lumjuan, N.; Walton, C.; Somboon, P. Investigation of Relative Development and Reproductivity Fitness Cost in Three Insecticide-Resistant Strains of Aedes aegypti from Thailand. Insects 2019, 10, 265. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Thomas, A.; Samuel, T.; Sahgal, A.; Verma, A.; Pillai, M.K.K. Diminished reproductive fitness associated with the deltamethrin resistance in an Indian strain of dengue vector mosquito, Aedes aegypti L. Trop. Biomed. 2009, 26, 155–164. [Google Scholar]
- Rigby, L.M.; Rašić, G.; Peatey, C.L.; Hugo, L.E.; Beebe, N.W.; Devine, G.J. Identifying the fitness costs of a pyrethroid-resistant genotype in the major arboviral vector Aedes aegypti. Parasites Vectors 2020, 13, 358. [Google Scholar] [CrossRef]
- Diniz, M.M.C.; Henriques, A.D.D.S.; Leandro, R.D.S.; Aguiar, D.L.; Beserra, E.B. Resistance of Aedes aegypti to temephos and adaptive disadvantages. Rev. De Saude Publica 2014, 48, 775–782. [Google Scholar] [CrossRef] [Green Version]
- Farnesi, L.C.; Martins, A.; Valle, D.; Rezende, G.L. Embryonic development of Aedes aegypti (Diptera: Culicidae): Influence of different constant temperatures. Mem. Do Inst. Oswaldo Cruz 2009, 104, 124–126. [Google Scholar] [CrossRef] [Green Version]
- Carrington, L.B.; Armijos, M.V.; Lambrechts, L.; Barker, C.; Scott, T.W. Effects of Fluctuating Daily Temperatures at Critical Thermal Extremes on Aedes aegypti Life-History Traits. PLoS ONE 2013, 8, e58824. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, M.U.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.; Shearer, F.M.; Barker, C.M.; Hay, S.I. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. eLife 2015, 4, e08347. [Google Scholar] [CrossRef] [PubMed]
- Lafferty, K.D. The ecology of climate change and infectious diseases. Ecology 2009, 90, 888–900. [Google Scholar] [CrossRef] [PubMed]
- Carrington, L.B.; Seifert, S.; Armijos, M.V.; Lambrechts, L.; Scott, T.W. Reduction of Aedes aegypti Vector Competence for Dengue Virus under Large Temperature Fluctuations. Am. J. Trop. Med. Hyg. 2013, 88, 689–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitek, C.J.; Livdahl, T. Hatch plasticity in response to varied inundation frequency in Aedes albopictus. J. Med. Èntomol. 2009, 46, 766–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- do Amaral, E.O.; da Silva, A.F.; da Silva, W.R.; da Silva, K.R.; Roque, R.A.; Tadei, W.P.; Zequi, J.A.C. Post-Embryonic Development of Aedes (Stegomyia) aegypti Linnaeus, 1762 at Different Temperatures and CO2 Concentrations, and Their Influences on Hatching and Development of Stabilized Population. In Life Cycle and Development of Diptera; Sarwar, M., Ed.; IntechOpen: London, UK, 2020; Available online: https://www.intechopen.com/books/life-cycle-and-development-of-diptera/post-embryonic-development-of-em-aedes-em-em-stegomyia-em-em-aegypti-em-linnaeus-1762-at-different-t (accessed on 23 November 2021).
- Zapata-Úsuga, G.; Zuleta-Ruiz, B.; Gómez-Vargas, W.; Mejía-Salazar, P.; Zuluaga-Ramírez, W. Presencia de Aedes albopictus (Diptera: Culicidae) en algunos municipios del área de influencia de la Proyecto Hidroeléctrico Ituango, Antioquia, Colombia. Actual. Biol. 2021, 44, 111. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Lowest Value | Highest Value |
|---|---|---|
| Eclosion time | Bello and Neiva | Riohacha |
| Time from L1 + L2 to L4 | Bello | Riohacha |
| Pupation time | Bello | Neiva |
| Female emergence time | Bello | Neiva |
| Male emergence time | Bello | Neiva and Riohacha |
| Egg hatching rate | Neiva | |
| Pupation rate | Neiva | |
| Emergence rate | Riohacha | |
| Female longevity | Bello | Riohacha |
| Male longevity | Bello | Riohacha |
| Egg mortality | Neiva | |
| Pupae mortality | Bello | |
| Fecundity | Neiva |
| Trait | Lowest Value | Highest Value |
|---|---|---|
| Eclosion time | Bello and Neiva 28 °C | Bello and Neiva 35 °C |
| Time from L1 + L2 to L4 | Bello and Neiva 28 °C | Bello and Neiva 35 °C Neiva (L3 to L4) 21 °C |
| Pupation time | Bello 28 °C Neiva 35 °C | Bello and Neiva 21 °C |
| Female emergence time | Bello 28 °C | |
| Male emergence time | Bello 28 °C | |
| Egg hatching rate | Bello and Neiva 35 °C | Bello and Neiva 21 and 28 °C |
| Pupation rate | Bello and Neiva 35 °C | Bello and Neiva 21 and 28 °C |
| Emergence rate | Bello and Neiva 35 °C | Bello and Neiva 21 and 28 °C |
| Egg mortality | Neiva 21, 28 and 35 °C Bello 35 °C | |
| L1 + L2 mortality | Bello and Neiva 35 °C | |
| Pupae mortality | Bello 28 °C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arévalo-Cortés, A.; Granada, Y.; Torres, D.; Triana-Chavez, O. Differential Hatching, Development, Oviposition, and Longevity Patterns among Colombian Aedes aegypti Populations. Insects 2022, 13, 536. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13060536
Arévalo-Cortés A, Granada Y, Torres D, Triana-Chavez O. Differential Hatching, Development, Oviposition, and Longevity Patterns among Colombian Aedes aegypti Populations. Insects. 2022; 13(6):536. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13060536
Chicago/Turabian StyleArévalo-Cortés, Andrea, Yurany Granada, David Torres, and Omar Triana-Chavez. 2022. "Differential Hatching, Development, Oviposition, and Longevity Patterns among Colombian Aedes aegypti Populations" Insects 13, no. 6: 536. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13060536