
Colour Response in Western Flower Thrips Varies Intraspecifically

 , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
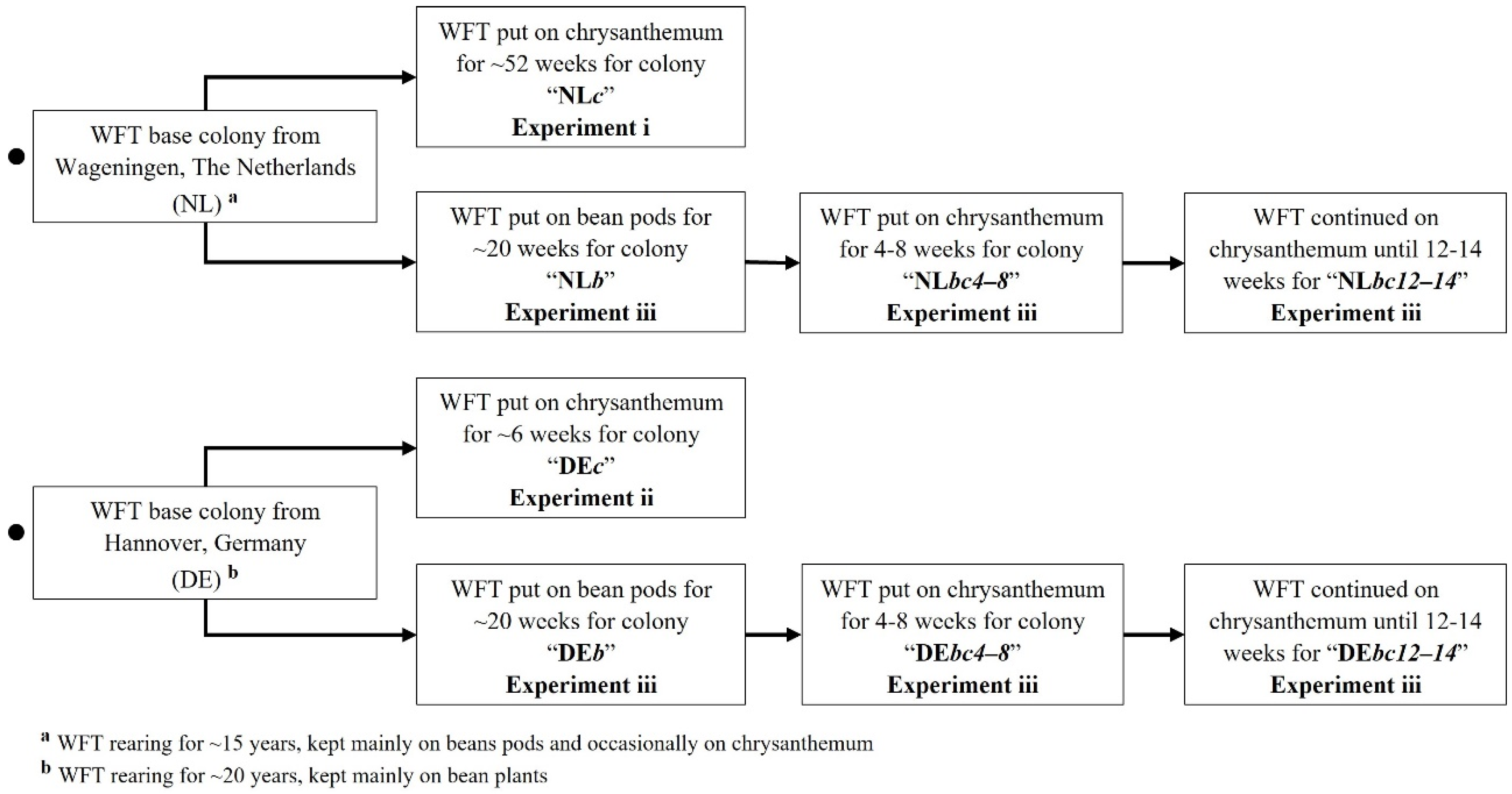
2.1. Insects: Management and Rearing of Colonies from The Netherlands and Germany
2.2. DNA Analysis
2.3. Wind Tunnel Setup
2.4. Experiments with LED Colours
2.5. Data Analysis
- Analysis a: WFT that landed on blue or yellow on the glass plate of the LED lamp.
- Analysis b: Response of WFT relative to the thrips that left the release container.
3. Results
3.1. Species Status of the Dutch (NL) and German (DE) Colonies
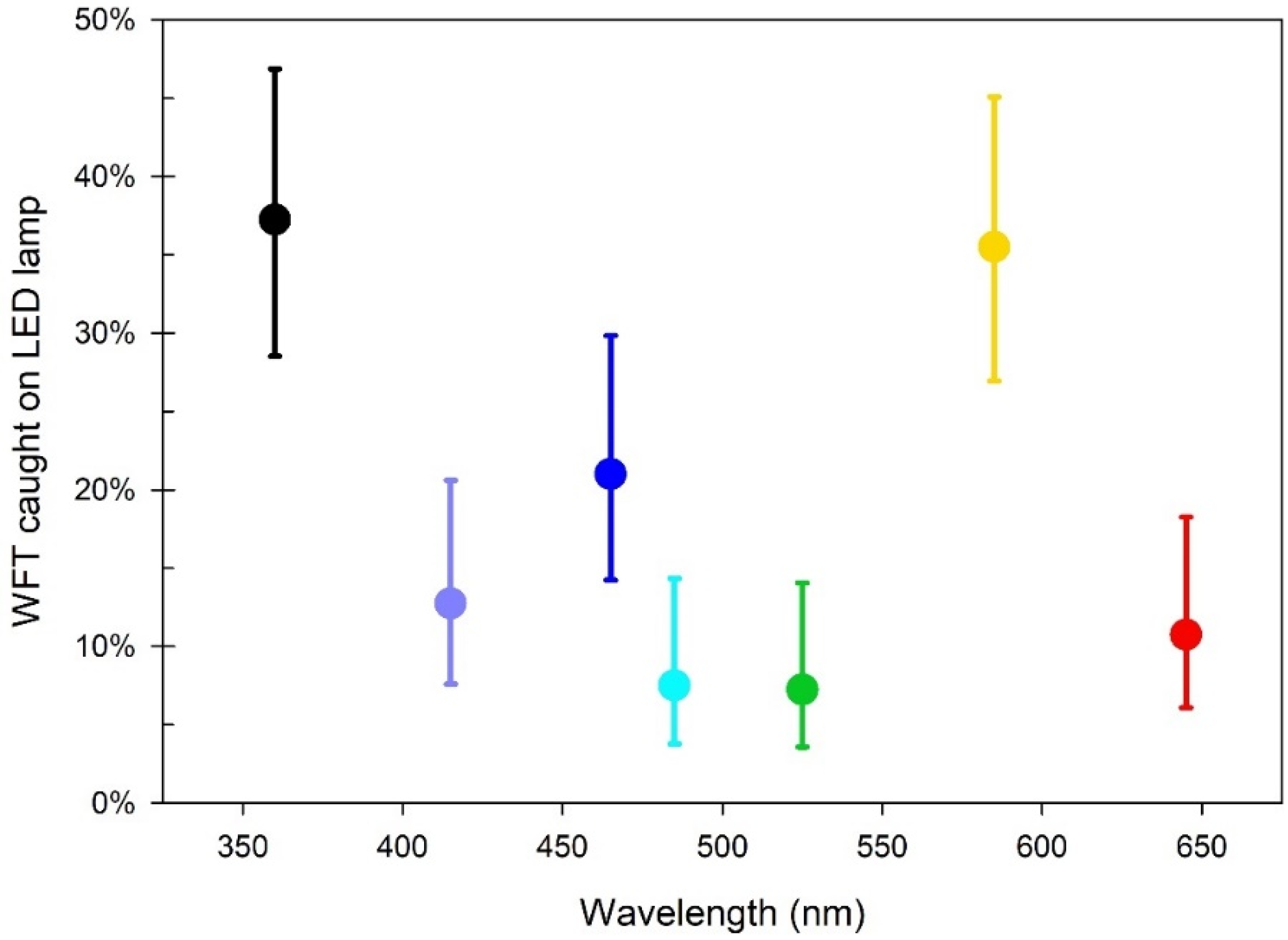
3.2. Experiment i: Response of NLc to Different LED Colours
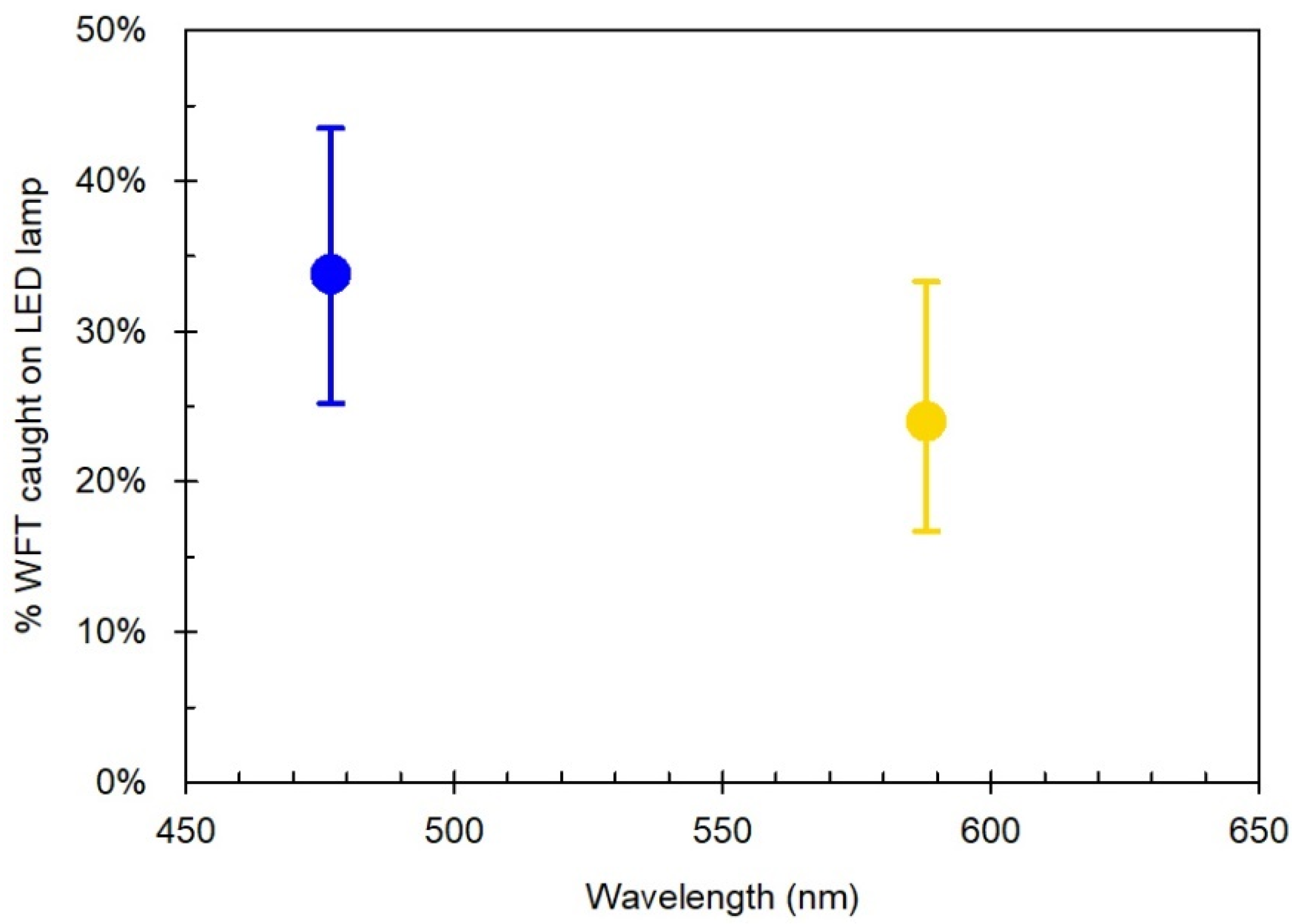
3.3. Experiment ii: Response of DEc to Blue and Yellow LEDs
3.4. Experiment iii: Blue and Yellow LED Choice Test of Dutch (NL) and German (DE) WFT from Different Host Plant Rearing Regimes
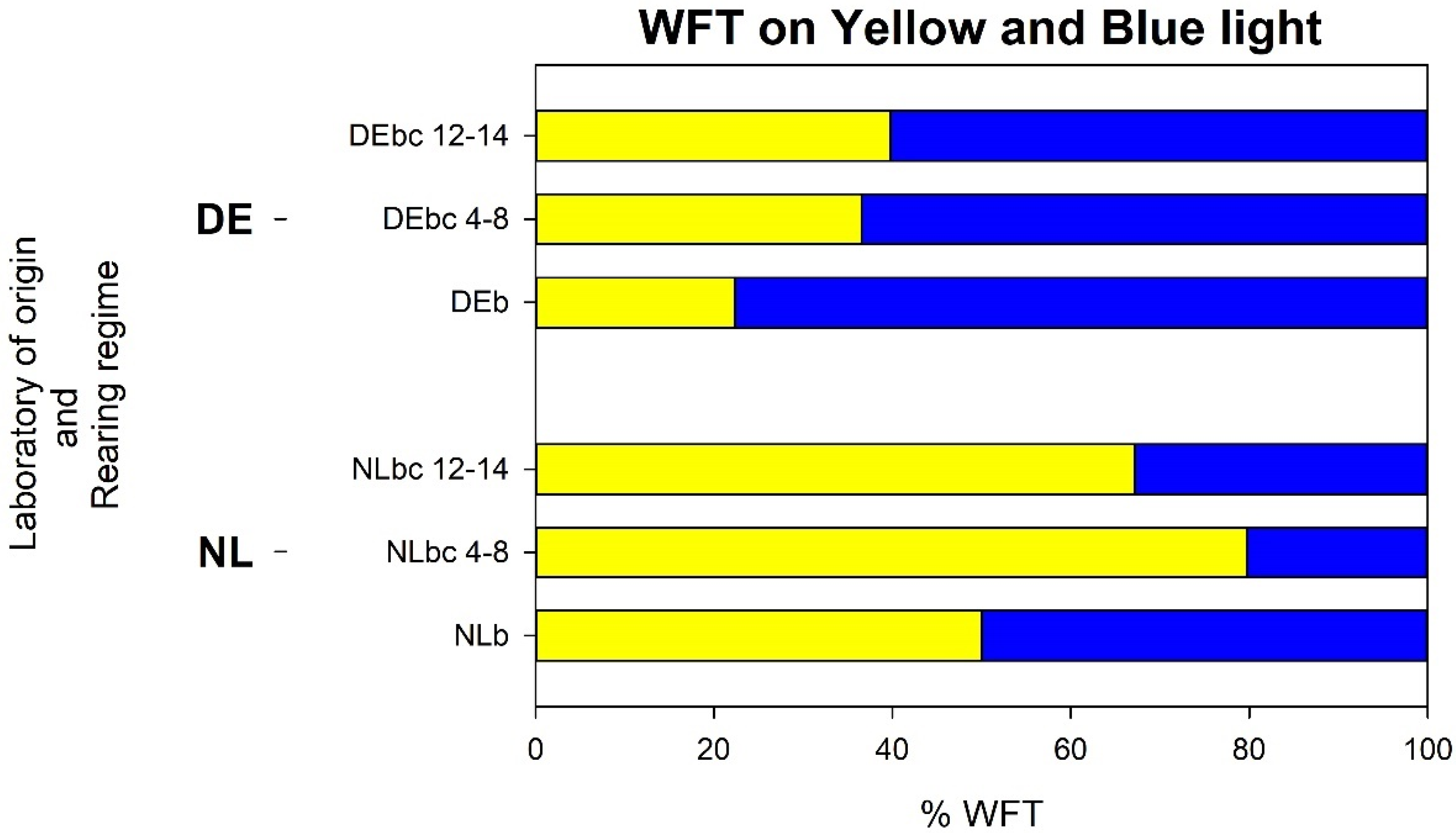
3.4.1. Analysis a: Response of WFT that Landed on Blue or Yellow on the Glass Plate of the LED Lamp
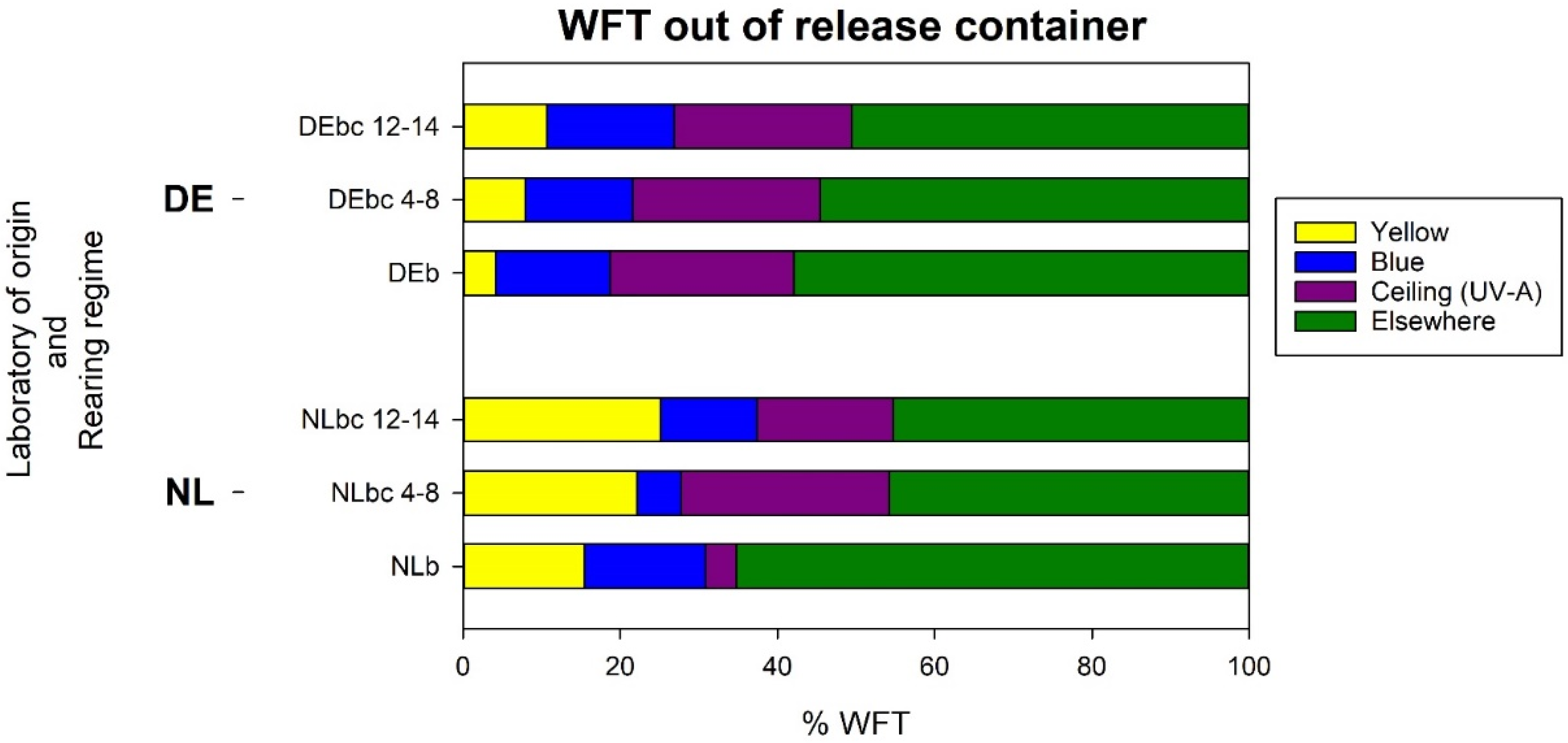
3.4.2. Analysis b: Response of WFT Relative to the Thrips That Left the Release Container
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gao, Y.-L.; Lei, Z.; Reitz, S.R. Western Flower Thrips Resistance to Insecticides: Detection, Mechanisms and Management Strategies. Pest Manag. Sci. 2012, 68, 1111–1121. [Google Scholar] [CrossRef] [PubMed]
- Jensen, S.E. Insecticide Resistance in the Western Flower Thrips, Frankliniella occidentalis. Integr. Pest Manag. Rev. 2000, 5, 131–146. [Google Scholar] [CrossRef]
- Li, D.; Shang, X.; Reitz, S.R.; Nauen, R.; Lei, Z.; Lee, S.H.; Gao, Y.-L. Field Resistance to Spinosad in Western Flower Thrips Frankliniella occidentalis (Thysanoptera: Thripidae). J. Integr. Agric. 2016, 15, 2803–2808. [Google Scholar] [CrossRef]
- Hillocks, R.J. Farming with Fewer Pesticides: EU Pesticide Review and Resulting Challenges for UK Agriculture. Crop Prot. 2012, 31, 85–93. [Google Scholar] [CrossRef]
- Karabelas, A.J.; Plakas, K.V.; Solomou, E.S.; Drossou, V.; Sarigiannis, D.A. Impact of European Legislation on Marketed Pesticides—A View from the Standpoint of Health Impact Assessment Studies. Environ. Int. 2009, 35, 1096–1107. [Google Scholar] [CrossRef]
- Lescourret, F. Toward a Reduced Use of Pesticides in European Farming Systems: An Introduction to the PURE Project. Crop Prot. 2017, 97, 7–9. [Google Scholar] [CrossRef]
- Lamichhane, J.R. Pesticide Use and Risk Reduction in European Farming Systems with IPM: An Introduction to the Special Issue. Crop Prot. 2017, 97, 1–6. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Arendse, W.; Dachbrodt-Saaydeh, S.; Kudsk, P.; Roman, J.C.; van Bijsterveldt-Gels, J.E.M.; Wick, M.; Messéan, A. Challenges and Opportunities for Integrated Pest Management in Europe: A Telling Example of Minor Uses. Crop Prot. 2015, 74, 42–47. [Google Scholar] [CrossRef]
- Jacobson, R.J. Integrated Pest Management (IPM) in Glasshouses. In Thrips as Crop Pests; CAB International: Wallingford, UK; New York, NY, USA, 1997; pp. 639–666. ISBN 0-85199-178-5. [Google Scholar]
- Lewis, T. Field and Laboratory Techniques. In Thrips as Crop Pests; CAB International: Wallingford, UK; New York, NY, USA, 1997; pp. 435–476. [Google Scholar]
- Kirk, W.D.J.; de Kogel, W.J.; Koschier, E.H.; Teulon, D.A. Semiochemicals for Thrips and Their Use in Pest Management. Annu. Rev. Entomol. 2021, 66, 101–119. [Google Scholar]
- van Tol, R.W.; Davidson, M.M.; Butler, R.C.; Teulon, D.A.; de Kogel, W.J. Visually and Olfactorily Enhanced Attractive Devices for Thrips Management. Entomol. Exp. Appl. 2020, 168, 665–677. [Google Scholar]
- Johansen, N.S.; Torp, T.; Solhaug, K.A. Phototactic Response of Frankliniella occidentalis to Sticky Traps with Blue Light Emitting Diodes in Herb and Alstroemeria Greenhouses. Crop Prot. 2018, 114, 120–128. [Google Scholar] [CrossRef]
- Makabe, T.; Futamura, T.; Noudomi, T.; Wakakuwa, M.; Arikawa, K. Phototaxis of Western Flower Thrips, Frankliniella occidentalis and Onion Thrips, Thrips Tabaci and the Possibility of Controlling Thrips Using Ultraviolet-Emitting Trap in the Greenhouse of Satsuma Mandarin (Citrus Unshiu). Jpn. J. Appl. Entomol. Zool. 2014, 58, 187–195. [Google Scholar] [CrossRef] [Green Version]
- Otieno, J.A.; Stukenberg, N.; Weller, J.; Poehling, H.-M. Efficacy of LED-Enhanced Blue Sticky Traps Combined with the Synthetic Lure Lurem-TR for Trapping of Western Flower Thrips (Frankliniella occidentalis). J. Pest Sci. 2018, 91, 1301–1314. [Google Scholar] [CrossRef]
- Yang, J.-Y.; Sung, B.-K.; Lee, H.-S. Phototactic Behavior 8: Phototactic Behavioral Responses of Western Flower Thrips, Frankliniella occidentalis Pergande (Thysanoptera: Thripidae), to Light-Emitting Diodes. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 359–363. [Google Scholar] [CrossRef]
- Lewis, T. (Ed.) Thrips as Crop Pests; CAB International: Wallingford, UK; New York, NY, USA, 1997; ISBN 0-85199-178-8. [Google Scholar]
- Brødsgaard, H.F. Coloured Sticky Traps for Frankliniella occidentalis (Pergande)(Thysanoptera, Thripidae) in Glasshouses. J. Appl. Entomol. 1989, 107, 136–140. [Google Scholar] [CrossRef]
- Matteson, N.A.; Terry, L.I. Response to Color by Male and Female Frankliniella occidentalis during Swarming and Non-swarming Behavior. Entomol. Exp. Appl. 1992, 63, 187–201. [Google Scholar] [CrossRef]
- Stukenberg, N.; Pietruska, M.; Waldherr, A.; Meyhöfer, R. Wavelength-Specific Behavior of the Western Flower Thrips (Frankliniella occidentalis): Evidence for a Blue-Green Chromatic Mechanism. Insects 2020, 11, 423. [Google Scholar] [CrossRef]
- Vernon, R.S.; Gillespie, D.R. Response of Frankliniella occidentalis (Thysanoptera: Thripidae) and Trialeurodes Vaporariorum (Homoptera: Aleyrodidae) to Fluorescent Traps in a Cucumber Greenhouse. J. Entomol. Soc. Br. Columbia 1990, 87, 38–41. [Google Scholar]
- Davidson, M.M.; Butler, R.C.; Teulon, D.A.J. Response of Female Frankliniella occidentalis (Pergande) to Visual Cues and Para-Anisaldehyde in a Flight Chamber. J. Insect Behav. 2012, 25, 297–307. [Google Scholar] [CrossRef]
- Prema, M.S.; Ganapathy, N.; Renukadevi, P.; Mohankumar, S.; Kennedy, J.S. Coloured Sticky Traps to Monitor Thrips Population in Cotton. J. Entomol. Zool. Stud. 2018, 6, 948–952. [Google Scholar]
- Ren, X.; Wu, S.; Xing, Z.; Xu, R.; Cai, W.; Lei, Z. Behavioral Responses of Western Flower Thrips (Frankliniella occidentalis) to Visual and Olfactory Cues at Short Distances. Insects 2020, 11, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smits, P.H.; van Deventer, P.; De Kogel, W.J. Western Flower Thrips: Reactions to Odours and Colours. West. Flower Thrips React. Odours Colours 2000, 11, 175–180. [Google Scholar]
- Cruz-Esteban, S.; Rojas, J.C.; Hernández-Ledesma, P. Trap Colour and Aggregation Pheromone Dose Affect the Catch of Western Flower Thrips in Blackberry Crops. J. Appl. Entomol. 2020, 144, 755–763. [Google Scholar] [CrossRef]
- Hoddle, M.S.; Robinson, L.; Morgan, D. Attraction of Thrips (Thysanoptera: Thripidae and Aeolothripidae) to Colored Sticky Cards in a California Avocado Orchard. Crop Prot. 2002, 21, 383–388. [Google Scholar] [CrossRef] [Green Version]
- Yudin, L.S.; Mitchell, W.C.; Cho, J.J. Color Preference of Thrips (Thysanoptera: Thripidae) with Reference to Aphids (Homoptera: Aphididae) and Leafminers in Hawaiian Lettuce Farms. J. Econ. Entomol. 1987, 80, 51–55. [Google Scholar] [CrossRef]
- Campan, R. Tactic Components in Orientation. EXS 1997, 84, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Jaenike, J. Host Specialization in Phytophagous Insects. Annu. Rev. Ecol. Syst. 1990, 21, 243–273. [Google Scholar] [CrossRef]
- Davidson, M.M.; Butler, R.C.; Teulon, D.A.J. Starvation Period and Age Affect the Response of Female Frankliniella occidentalis (Pergande)(Thysanoptera: Thripidae) to Odor and Visual Cues. J. Insect Physiol. 2006, 52, 729–736. [Google Scholar] [CrossRef]
- Fatnassi, H.; Pizzol, J.; Senoussi, R.; Biondi, A.; Desneux, N.; Poncet, C.; Boulard, T. Within-Crop Air Temperature and Humidity Outcomes on Spatio-Temporal Distribution of the Key Rose Pest Frankliniella occidentalis. PLoS ONE 2015, 10, e0126655. [Google Scholar] [CrossRef]
- Nielsen, M.-C. Factors Affecting the Response of Thrips to an Olfactory Cue. Ph.D. Dissertation, Lincoln University, Lincoln, UK, 2013. [Google Scholar]
- Teulon, D.A.J.; Hollister, B.; Butler, R.C.; Cameron, E.A. Colour and Odour Responses of Flying Western Flower Thrips: Wind Tunnel and Greenhouse Experiments. Entomol. Exp. Appl. 1999, 93, 9–19. [Google Scholar] [CrossRef]
- Van Tol, R.W.; Tom, J.; Roher, M.; Schreurs, A.; van Dooremalen, C. Haze of Glue Determines Preference of Western Flower Thrips (Frankliniella occidentalis) for Yellow or Blue Traps. Sci. Rep. 2021, 11, 6557. [Google Scholar] [CrossRef] [PubMed]
- Rugman-Jones, P.F.; Hoddle, M.S.; Stouthamer, R. Nuclear-Mitochondrial Barcoding Exposes the Global Pest Western Flower Thrips (Thysanoptera: Thripidae) as Two Sympatric Cryptic Species in Its Native California. J. Econ. Entomol. 2010, 103, 877–886. [Google Scholar] [CrossRef] [PubMed]
- OEPP/EPPO. PM 7/129 (1) DNA Barcoding as an Identification Tool for a Number of Regulated Pests. EPPO Bull 2016, 46, 501–537. [CrossRef] [Green Version]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA Primers for Amplification of Mitochondrial Cytochrome c Oxidase Subunit I from Diverse Metazoan Invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Kimura, M. A Simple Method for Estimating Evolutionary Rates of Base Substitutions through Comparative Studies of Nucleotide Sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- McCullagh, P.; Nelder, J.A. Generalized Linear Models; Chapman and Hall: London, UK, 1989; ISBN 0-203-75373-9. [Google Scholar]
- Payne, R.; Murray, D.; Baird, D. The Guide to the Genstat Command Language (Release 19); VSN International: Hemel Hempsted, UK, 2017. [Google Scholar]
- Kadirvel, P.; Srinivasan, R.; Hsu, Y.-C.; Su, F.-C.; De La Pena, R. Application of Cytochrome Oxidase I Sequences for Phylogenetic Analysis and Identification of Thrips Species Occurring on Vegetable Crops. J. Econ. Entomol. 2013, 106, 408–418. [Google Scholar] [CrossRef]
- Zhang, G.F.; Meng, X.Q.; Min, L.; Qiao, W.N.; Wan, F.H. Rapid Diagnosis of the Invasive Species, Frankliniella occidentalis (Pergande): A Species-specific COI Marker. J. Appl. Entomol. 2012, 136, 410–420. [Google Scholar] [CrossRef]
- Gunawardana, D.N.; Li, D.; Masumoto, M.; Mound, L.A.; O’Donnel, C.A.; Skarlinsky, T.L. Resolving the Confused Identity of Frankliniella Panamensis (Thysanoptera: Thripidae). Zootaxa 2017, 4323, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Mound, L.A.; Wheeler, G.S.; Williams, D.A. Resolving Cryptic Species with Morphology and DNA; Thrips as a Potential Biocontrol Agent of Brazilian Peppertree, with a New Species and Overview of Pseudophilothrips (Thysanoptera). Zootaxa 2010, 2432, 59–68. [Google Scholar] [CrossRef]
- Nielsen, M.-C.; Teulon, D.A.J.; Chapman, R.B.; Butler, R.C.; Drayton, G.M.; Philipsen, H. Comparison of Life History Parameters of Two Frankliniella occidentalis (Thysanoptera: Thripidae) Strains in New Zealand. Environ. Entomol. 2010, 39, 303–311. [Google Scholar] [CrossRef]
- Fang, J.; Kritzman, A.; Yonash, N.; Gera, A.; Pollak, N.; Lavi, U. Genetic Variation of Thrips Populations Assessed by Amplified Fragment Length Polymorphism (Thysanoptera: Thripidae). Ann. Entomol. Soc. Am. 2005, 98, 351–358. [Google Scholar] [CrossRef]
- Mirnezhad, M.; Schidlo, N.; Klinkhamer, P.G.; Leiss, K.A. Variation in Genetics and Performance of Dutch Western Flower Thrips Populations. J. Econ. Entomol. 2012, 105, 1816–1824. [Google Scholar] [CrossRef]
- Brunner, P.C.; Frey, J.E. Habitat-Specific Population Structure in Native Western Flower Thrips Frankliniella occidentalis (Insecta, Thysanoptera). J. Evol. Biol. 2010, 23, 797–804. [Google Scholar] [CrossRef]
- Chittka, L.; Ings, T.C.; Raine, N.E. Chance and Adaptation in the Evolution of Island Bumblebee Behaviour. Popul. Ecol. 2004, 46, 243–251. [Google Scholar] [CrossRef] [Green Version]
- Szentesi, Á.; Jermy, T. The Role of Experience in Host Plant Choice by Phytophagous Insects. In Insect-Plant Interactions; CRC Press: Boca Raton, FL, USA, 1990; ISBN 978-0-203-71173-6. [Google Scholar]
- Döring, T.F. How Aphids Find Their Host Plants, and How They Don’t. Ann. Appl. Biol. 2014, 165, 3–26. [Google Scholar] [CrossRef]
- Döring, T.F.; Chittka, L. Visual Ecology of Aphids—A Critical Review on the Role of Colours in Host Finding. Arthropod-Plant Interact. 2007, 1, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Kirk, W.D.J. Ecological Studies on Thrips Imaginis Bagnall (Thysanoptera) in Flowers of Echium Plantagineum L. in Australia. Aust. J. Ecol. 1984, 9, 9–18. [Google Scholar] [CrossRef]
- Prokopy, R.J.; Owens, E.D. Visual Detection of Plants by Herbivorous Insects. Annu. Rev. Entomol. 1983, 28, 337–364. [Google Scholar] [CrossRef]
- Reeves, J.L. Vision Should Not Be Overlooked as an Important Sensory Modality for Finding Host Plants. Environ. Entomol. 2011, 40, 855–863. [Google Scholar] [CrossRef]
- Rogge, S.A.; Meyhöfer, R. The Role of Plant Physiology and Cultivar of Chrysanthemum in the Resistance against Western Flower Thrips. Entomol. Exp. Appl. 2021, 169, 275–289. [Google Scholar] [CrossRef]
- Banschbach, V.S. Colour Association Influences Honey Bee Choice between Sucrose Concentrations. J. Comp. Physiol. A 1994, 175, 107–114. [Google Scholar] [CrossRef]
- Giurfa, M. Conditioning Procedure and Color Discrimination in the Honeybee Apis Mellifera. Naturwissenschaften 2004, 91, 228–231. [Google Scholar] [CrossRef] [PubMed]
- Muth, F.; Papaj, D.R.; Leonard, A.S. Bees Remember Flowers for More than One Reason: Pollen Mediates Associative Learning. Anim. Behav. 2016, 111, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Nicholls, E.; Hempel de Ibarra, N. Bees Associate Colour Cues with Differences in Pollen Rewards. J. Exp. Biol. 2014, 217, 2783–2788. [Google Scholar] [CrossRef] [Green Version]
- Nicholls, E.; Hempel de Ibarra, N. Assessment of Pollen Rewards by Foraging Bees. Funct. Ecol. 2017, 31, 76–87. [Google Scholar] [CrossRef] [Green Version]
- Wilmsen, S.; Gottlieb, R.; Junker, R.R.; Lunau, K. Bumblebees Require Visual Pollen Stimuli to Initiate and Multimodal Stimuli to Complete a Full Behavioral Sequence in Close-range Flower Orientation. Ecol. Evol. 2017, 7, 1384–1393. [Google Scholar] [CrossRef] [Green Version]
- Tommasini, M.G.; Maini, S. Frankliniella Occidentalis and Other Thrips Harmful to Vegetable and Ornamental Crops in Europe; Landbouwuniversiteit Wageningen (Wageningen Agricultural University): Wageningen, The Netherlands, 1995. [Google Scholar]
- Gerin, C.; Hance, T.H.; Van Impe, G. Impact of Flowers on the Demography of Western Flower Thrips Frankliniella occidentalis (Thysan., Thripidae). J. Appl. Entomol. 1999, 123, 569–574. [Google Scholar] [CrossRef]
- Mound, L.A. Thysanoptera: Diversity and Interactions. Annu. Rev. Entomol. 2005, 50, 247–269. [Google Scholar] [CrossRef]
- Pearsall, I.A. Flower Preference Behaviour of Western Flower Thrips in the Similkameen Valley, British Columbia, Canada. Entomol. Exp. Appl. 2000, 95, 303–313. [Google Scholar] [CrossRef]
- Blumthal, M.R.; Spomer, L.A.; Warnock, D.F.; Cloyd, R.A. Flower Color Preferences of Western Flower Thrips. HortTechnology 2005, 15, 846–853. [Google Scholar] [CrossRef] [Green Version]
- Kirk, W.D.J. Feeding. In Thrips as Crop Pests; CAB International: Wallingford, UK; New York, NY, USA, 1997; pp. 119–174. [Google Scholar]
- Broadbent, A.B.; Matteoni, J.A.; Allen, W.R. Feeding Preferences of the Western Flower Thrips, Frankliniella occidentalis (Pergande)(Thysanoptera: Thripidae), and Incidence of Tomato Spotted Wilt Virus among Cultivars of Florist’s Chrysanthemum. Can. Entomol. 1990, 122, 1111–1117. [Google Scholar] [CrossRef]
- De Jager, C.M.; Butôt, R.P.T.; De Jong, T.J.; Klinkhamer, P.G.L.; Van Der Meijden, E. Population Growth and Survival of Western Flower Thrips Frankliniella occidentalis Pergande (Thysanoptera, Thripidae) on Different Chrysanthemum Cultivars: Two Methods for Measuring Resistance. J. Appl. Entomol. 1993, 115, 519–525. [Google Scholar] [CrossRef]
- De Jager, C.M.; Butôt, R.P.T.; Klinkhamer, P.G.L.; De Jong, T.J.; Wolff, K.; Van Der Meijden, E. Genetic Variation in Chrysanthemum for Resistance to Frankliniella occidentalis. Entomol. Exp. Appl. 1995, 77, 277–287. [Google Scholar] [CrossRef]
- Kirk, W.D.J. Distribution, Abundance and Population Dynamics. In Thrips as Crop Pests; CAB International: Wallingford, UK; New York, NY, USA, 1997; pp. 217–257. [Google Scholar]
- Lewis, T. Appendix II: Major Crops Infested by Thrips with Main Symptoms and Predominant Injurious Species. In Thrips as Crop Pests; CAB International: Wallingford, UK; New York, NY, USA, 1997; pp. 675–709. [Google Scholar]
- Brodbeck, B.V.; Funderburk, J.; Stavisky, J.; Andersen, P.C.; Hulshof, J.; Marullo, R.; Mound, L.A. Recent Advances in the Nutritional Ecology of Thysanoptera, or the Lack Thereof. In Proceedings of the 7th International Symposium on Thysanoptera; Australian National Insect Collection: Canberra, Australia, 2002; Volume 7, pp. 145–153. [Google Scholar]
- Hulshof, J.; Vanninen, I. Western Flower Thrips Feeding on Pollen, and Its Implications for Control. In Proceedings of the 7th International Symposium on Thysanoptera; Australian National Insect Collection: Canberra, Australia, 2002; Volume 7, pp. 173–179. [Google Scholar]
- Kirk, W.D.J. Aggregation and Mating of Thrips in Flowers of Calystegia Sepium. Ecol. Entomol. 1985, 10, 433–440. [Google Scholar] [CrossRef]
- Kirk, W.D.J. Pollen-feeding and the Host Specificity and Fecundity of Flower Thrips (Thysanoptera). Ecol. Entomol. 1985, 10, 281–289. [Google Scholar] [CrossRef]
- Leskey, T.C.; Teulon, D.A.; Cameron, E.A. Effects of Temperature and Sugar Maple Pollen on Oviposition and Longevity of Pear Thrips (Thysanoptera: Thripidae). Environ. Entomol. 1997, 26, 566–571. [Google Scholar] [CrossRef]
- Yaku, A.; Walter, G.H.; Najar-Rodriguez, A.J. Thrips See Red–Flower Colour and the Host Relationships of a Polyphagous Anthophilic Thrips. Ecol. Entomol. 2007, 32, 527–535. [Google Scholar] [CrossRef]
- Milne, M.; Walter, G.H. Feeding and Breeding across Host Plants within a Locality by the Widespread Thrips Frankliniella Schultzei, and the Invasive Potential of Polyphagous Herbivores. Divers. Distrib. 2000, 6, 243–257. [Google Scholar] [CrossRef]
- Skorupski, P.; Döring, T.F.; Chittka, L. Photoreceptor Spectral Sensitivity in Island and Mainland Populations of the Bumblebee, Bombus Terrestris. J. Comp. Physiol. A 2007, 193, 485–494. [Google Scholar] [CrossRef]
- Hollis, K.L.; Guillette, L.M. What Associative Learning in Insects Tells Us about the Evolution of Learned and Fixed Behavior. Int. J. Comp. Psychol. 2015, 28, 1–18. [Google Scholar] [CrossRef]
- Polilov, A.A.; Makarova, A.A.; Kolesnikova, U.K. Cognitive Abilities with a Tiny Brain: Neuronal Structures and Associative Learning in the Minute Nephanes Titan (Coleoptera: Ptiliidae). Arthropod Struct. Dev. 2019, 48, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Hollis, K.L.; Guillette, L.M. Associative Learning in Insects: Evolutionary Models, Mushroom Bodies, and a Neuroscientific Conundrum. Comp. Cogn. Behav. Rev. 2011, 6, 24–45. [Google Scholar] [CrossRef] [Green Version]
- Egri, Á.; Farkas, P.; Bernáth, B.; Guerin, P.M.; Fail, J. Spectral sensitivity of L2 biotype in the Thrips tabaci cryptic species complex. J. Insect Physiol. 2020, 121, 103999. [Google Scholar] [CrossRef]
- Cronin, T.W.; Bok, M.J. Photoreception and Vision in the Ultraviolet. J. Exp. Biol. 2016, 219, 2790–2801. [Google Scholar] [CrossRef] [Green Version]
- Dáder, B.; Moreno, A.; Gwynn-Jones, D.; Winters, A.; Fereres, A. Aphid Orientation and Performance in Glasshouses under Different UV-A/UV-B Radiation Regimes. Entomol. Exp. Appl. 2017, 163, 344–353. [Google Scholar] [CrossRef]
- Doukas, D.; Payne, C.C. Greenhouse Whitefly (Homoptera: Aleyrodidae) Dispersal under Different UV-Light Environments. J. Econ. Entomol. 2007, 100, 389–397. [Google Scholar] [CrossRef]
- Stukenberg, N.; Gebauer, K.; Poehling, H.-M. Light Emitting Diode (LED)-based Trapping of the Greenhouse Whitefly (Trialeurodes Vaporariorum). J. Appl. Entomol. 2015, 139, 268–279. [Google Scholar] [CrossRef]
- Nguyen, T.H.N.; Borgemeister, C.; Max, J.; Poehling, H.-M. Manipulation of Ultraviolet Light Affects Immigration Behavior of Ceratothripoides Claratris (Thysanoptera: Thripidae). J. Econ. Entomol. 2009, 102, 1559–1566. [Google Scholar] [CrossRef]
- Goldsmith, T.H. Ultraviolet Receptors and Color Vision: Evolutionary Implications and a Dissonance of Paradigms. Vision Res. 1994, 34, 1479–1487. [Google Scholar] [CrossRef]
- Menzel, R. Spectral Sensitivity and Color Vision in Invertebrates. In Comparative Physiology and Evolution of Vision in Invertebrates; Springer: Berlin/Heidelberg, Germany, 1979; pp. 503–580. [Google Scholar]
- Gumbert, A. Color Choices by Bumble Bees (Bombus Terrestris): Innate Preferences and Generalization after Learning. Behav. Ecol. Sociobiol. 2000, 48, 36–43. [Google Scholar] [CrossRef]
- Kelber, A. Innate Preferences for Flower Features in the Hawkmoth Macroglossum Stellatarum. J. Exp. Biol. 1997, 200, 827–836. [Google Scholar] [CrossRef] [PubMed]
- Kuenzinger, W.; Kelber, A.; Weesner, J.; Travis, J.; Raguso, R.A.; Goyret, J. Innate Colour Preferences of a Hawkmoth Depend on Visual Context. Biol. Lett. 2019, 15, 20180886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lunau, K. Colour Saturation Triggers Innate Reactions to Flower Signals: Flower Dummy Experiments with Bumblebees. J. Comp. Physiol. A 1990, 166, 827–834. [Google Scholar] [CrossRef]
- Ostroverkhova, O.; Galindo, G.; Lande, C.; Kirby, J.; Scherr, M.; Hoffman, G.; Rao, S. Understanding Innate Preferences of Wild Bee Species: Responses to Wavelength-Dependent Selective Excitation of Blue and Green Photoreceptor Types. J. Comp. Physiol. A 2018, 204, 667–675. [Google Scholar] [CrossRef]
- Satoh, A.; Kinoshita, M.; Arikawa, K. Innate Preference and Learning of Colour in the Male Cotton Bollworm Moth, Helicoverpa Armigera. J. Exp. Biol. 2016, 219, 3857–3860. [Google Scholar]
- Yoshida, M.; Itoh, Y.; Ômura, H.; Arikawa, K.; Kinoshita, M. Plant Scents Modify Innate Colour Preference in Foraging Swallowtail Butterflies. Biol. Lett. 2015, 11, 20150390. [Google Scholar] [CrossRef] [Green Version]
- Stukenberg, N.; Poehling, H.-M. Blue–Green Opponency and Trichromatic Vision in the Greenhouse Whitefly (Trialeurodes Vaporariorum) Explored Using Light Emitting Diodes. Ann. Appl. Biol. 2019, 175, 146–163. [Google Scholar] [CrossRef]
- Farnier, K.; Dyer, A.G.; Steinbauer, M.J. Related but Not Alike: Not All Hemiptera are Attracted to Yellow. Front. Ecol. Evol. 2014, 2, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Otani, Y.; Wakakuwa, M.; Arikawa, K. Relationship between Action Spectrum and Spectral Sensitivity of Compound Eyes Relating Phototactic Behavior of the Western Flower Thrips, Frankliniella occidentalis. Jpn. J. Appl. Entomol. Zool. 2014, 58, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Virgilio, M.; Backeljau, T.; Nevado, B.; De Meyer, M. Comparative Performances of DNA Barcoding across Insect Orders. BMC Bioinform. 2010, 11, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | LED Colour a | Peak Wavelength (nm) |
|---|---|---|
| 1 | UV-A | 369 |
| 2 | Violet | 422 |
| 3 | Blue | 477 |
| 4 | Cyan | 502 |
| 5 | Green | 529 |
| 6 | Yellow | 588 |
| 7 | Red | 651 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopez-Reyes, K.; Armstrong, K.F.; Teulon, D.A.J.; Butler, R.C.; van Dooremalen, C.; Roher, M.; van Tol, R.W.H.M. Colour Response in Western Flower Thrips Varies Intraspecifically. Insects 2022, 13, 538. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13060538
Lopez-Reyes K, Armstrong KF, Teulon DAJ, Butler RC, van Dooremalen C, Roher M, van Tol RWHM. Colour Response in Western Flower Thrips Varies Intraspecifically. Insects. 2022; 13(6):538. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13060538
Chicago/Turabian StyleLopez-Reyes, Karla, Karen F. Armstrong, David A. J. Teulon, Ruth C. Butler, Coby van Dooremalen, Monika Roher, and Robert W. H. M. van Tol. 2022. "Colour Response in Western Flower Thrips Varies Intraspecifically" Insects 13, no. 6: 538. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13060538