
Determination of Moisture and Protein Content in Living Mealworm Larvae (Tenebrio molitor L.) Using Near-Infrared Reflectance Spectroscopy (NIRS)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Samples and Feeding Groups
2.2. Spectra Collection
2.3. Multivariate Analysis
2.4. Proximate Analysis
2.5. Statistical Analysis
3. Results
3.1. Growth Rate and Feed Conversion Efficiency
3.2. Nutritional Analysis
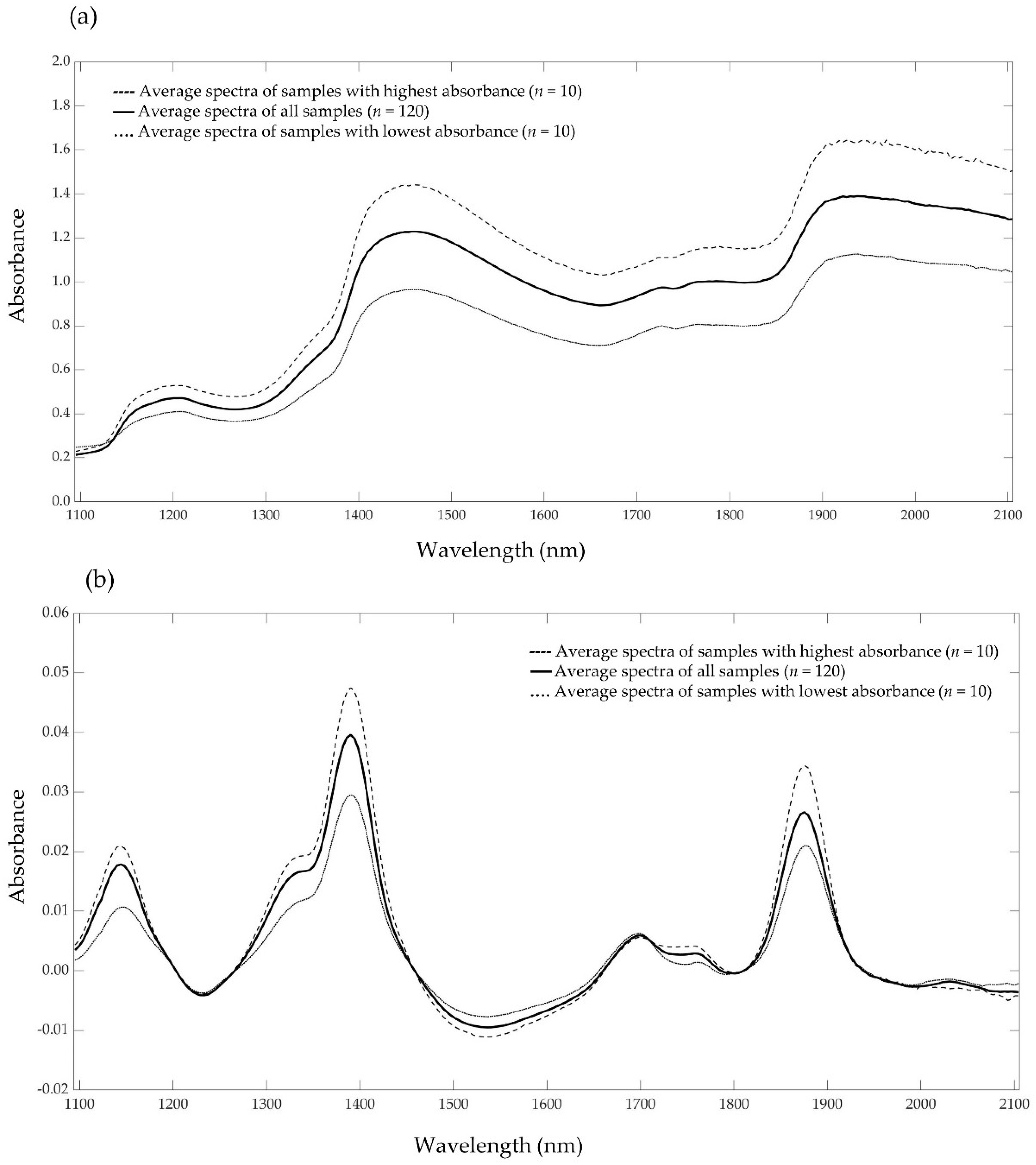
3.3. NIR Spectra and Reference Data
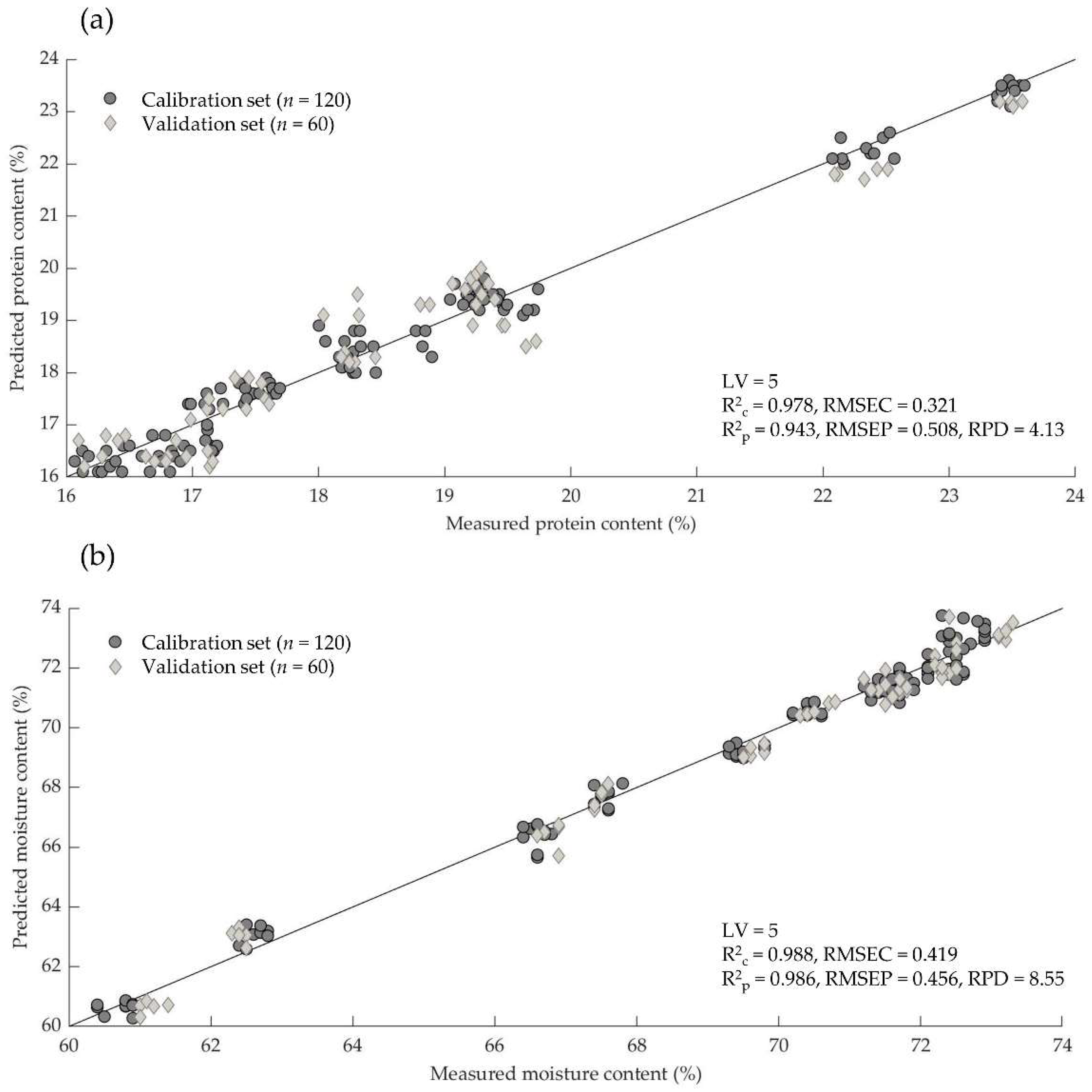
3.4. Prediction of Moisture and Protein Content
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grafton, R.Q.; Daugbjerg, C.; Qureshi, M.E. Towards food security by 2050. Food Secur. 2015, 7, 179–183. [Google Scholar] [CrossRef]
- Belluco, S.; Losasso, C.; Maggioletti, M.; Alonzi, C.C.; Paoletti, M.G.; Ricci, A. Edible Insects in a Food Safety and Nutritional Perspective: A Critical Review. Compr. Rev. Food Sci. Food Saf. 2013, 12, 296–313. [Google Scholar] [CrossRef]
- Dobermann, D.; Swift, J.A.; Field, L.M. Opportunities and hurdles of edible insects for food and feed. Nutr. Bull. 2017, 42, 293–308. [Google Scholar] [CrossRef] [Green Version]
- van Huis, A.; Oonincx, D.G.A.B. The environmental sustainability of insects as food and feed. A review. Agron. Sustain. Dev. 2017, 37, 293–308. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.; Suleria, H.A.R.; Rauf, A. Edible insects as innovative foods: Nutritional and functional assessments. Trends Food Sci. Technol. 2019, 86, 352–359. [Google Scholar] [CrossRef]
- Nikkhah, A.; van Haute, S.; Jovanovic, V.; Jung, H.; Dewulf, J.; Cirkovic Velickovic, T.; Ghnimi, S. Life cycle assessment of edible insects (Protaetia brevitarsis seulensis larvae) as a future protein and fat source. Sci. Rep. 2021, 11, 14030. [Google Scholar] [CrossRef] [PubMed]
- Finke, M.D. Nutrient Content of Insects. In Encyclopedia of Entomology; Capinera, J.L., Ed.; Kluwer Academic Publ: Dordrecht, The Netherlands, 2004; pp. 1563–1575. [Google Scholar]
- Finke, M.D. Complete nutrient content of four species of feeder insects. Zoo Biol. 2013, 32, 27–36. [Google Scholar] [CrossRef]
- Hong, J.; Han, T.; Kim, Y.Y. Mealworm (Tenebrio molitor Larvae) as an Alternative Protein Source for Monogastric Animal: A Review. Anim. Open Access J. MDPI 2020, 10, 2068. [Google Scholar] [CrossRef]
- Stull, V.J.; Kersten, M.; Bergmans, R.S.; Patz, J.A.; Paskewitz, S. Crude Protein, Amino Acid, and Iron Content of Tenebrio molitor (Coleoptera, Tenebrionidae) Reared on an Agricultural Byproduct from Maize Production: An Exploratory Study. Ann. Entomol. Soc. Am. 2019, 112, 533–543. [Google Scholar] [CrossRef]
- Mariod, A.A. Nutrient Composition of Mealworm (Tenebrio molitor). In African Edible Insects as Alternative Source of Food, Oil, Protein and Bioactive Components; Adam Mariod, A., Ed.; Springer eBook Collection, 1st ed.; Springer International Publishing: Cham, Germany, 2020; pp. 275–280. [Google Scholar]
- Bukkens, S.G. The nutritional value of edible insects. Ecol. Food Nutr. 1997, 36, 287–319. [Google Scholar] [CrossRef]
- Adámková, A.; Adámek, M.; Mlček, J.; Borkovcová, M.; Bednářová, M.; Kouřimská, L.; Skácel, J.; Ví tová, E. Welfare of the mealworm (Tenebrio molitor) breeding with regard to nutrition value and food safety. Potravin. Slovak J. Food Sci. 2017, 11, 460–465. [Google Scholar] [CrossRef] [Green Version]
- Murray, D.R.P. The importance of water on the normal growth of larvae of Tenebrio molitor. Entomol. Exp. Et Appl. 1968, 11, 149–168. [Google Scholar] [CrossRef]
- Rumpold, B.A.; Schlüter, O.K. Potential and challenges of insects as an innovative source for food and feed production. Innov. Food Sci. Emerg. Technol. 2013, 17, 1–11. [Google Scholar] [CrossRef]
- Sánchez-Muros, M.-J.; Barroso, F.G.; Manzano-Agugliaro, F. Insect meal as renewable source of food for animal feeding: A review. J. Clean. Prod. 2014, 65, 16–27. [Google Scholar] [CrossRef]
- Payne, C.L.; Scarborough, P.; Rayner, M.; Nonaka, K. A systematic review of nutrient composition data available for twelve commercially available edible insects, and comparison with reference values. Trends Food Sci. Technol. 2016, 47, 69–77. [Google Scholar] [CrossRef]
- Dreassi, E.; Cito, A.; Zanfini, A.; Materozzi, L.; Botta, M.; Francardi, V. Dietary fatty acids influence the growth and fatty acid composition of the yellow mealworm Tenebrio molitor (Coleoptera: Tenebrionidae). Lipids 2017, 52, 285–294. [Google Scholar] [CrossRef]
- Oonincx, D.G.A.B.; van Broekhoven, S.; van Huis, A.; van Loon, J.J.A. Feed Conversion, Survival and Development, and Composition of Four Insect Species on Diets Composed of Food By-Products. PLoS ONE 2015, 10, e0144601. [Google Scholar] [CrossRef] [Green Version]
- Mirzaeva, D.A.; Khujamshukurov, N.A.; Zokirov, B.; Soxibov, B.O.; Kuchkarova, D.K. Influence of Temperature and Humidity on the Development of Tenebrio molitor L. Int. J. Curr. Microbiol. Appl. Sci. 2020, 9, 3544–3559. [Google Scholar] [CrossRef]
- Johnsen, N.S.; Andersen, J.L.; Offenberg, J. The effect of relative humidity on the survival and growth rate of the yellow mealworm larvae (Tenebrio molitor, Linnaeus 1758). J. Insects Food Feed. 2021, 7, 311–318. [Google Scholar] [CrossRef]
- Prieto, N.; Andrés, S.; Giráldez, F.J.; Mantecón, A.R.; Lavín, P. Ability of near infrared reflectance spectroscopy (NIRS) to estimate physical parameters of adult steers (oxen) and young cattle meat samples. Meat Sci. 2008, 79, 692–699. [Google Scholar] [CrossRef]
- Fernández Pierna, J.A.; Vermeulen, P.; Amand, O.; Tossens, A.; Dardenne, P.; Baeten, V. NIR hyperspectral imaging spectroscopy and chemometrics for the detection of undesirable substances in food and feed. Chemom. Intell. Lab. Syst. 2012, 117, 233–239. [Google Scholar] [CrossRef]
- Kobayashi, K.-I.; Mori, M.; Nishino, K.; Toyota, T.; Nakauchi, S. Visualisation of Fat and Fatty Acid Distribution in Beef Using a Set of Filters Based on near Infrared Spectroscopy. J. Near Infrared Spectrosc. 2012, 20, 509–519. [Google Scholar] [CrossRef]
- Burger, J.; Geladi, P. Hyperspectral NIR imaging for calibration and prediction: A comparison between image and spectrometer data for studying organic and biological samples. Analyst 2006, 131, 1152–1160. [Google Scholar] [CrossRef] [PubMed]
- Osborne, B.G.; Fearn, T.; Hindle, P.H. Practical NIR Spectroscopy with Applications in Food and Beverage Analysis, 2nd ed.; Longman Food Technology; Longman Scientific & Technical: Harlow, UK, 1993. [Google Scholar]
- Prieto, N.; Roehe, R.; Lavín, P.; Batten, G.; Andrés, S. Application of near infrared reflectance spectroscopy to predict meat and meat products quality: A review. Meat Sci. 2009, 83, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Isaksson, T.; Nilsen, B.N.; Tøgersen, G.; Hammond, R.P.; Hildrum, K.I. On-line, proximate analysis of ground beef directly at a meat grinder outlet. Meat Sci. 1996, 43, 245–253. [Google Scholar] [CrossRef]
- Prieto, N.; Andrés, S.; Giráldez, F.J.; Mantecón, A.R.; Lavín, P. Potential use of near infrared reflectance spectroscopy (NIRS) for the estimation of chemical composition of oxen meat samples. Meat Sci. 2006, 74, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Bruno-Soares, A.M.; Murray, I.; Paterson, R.M.; Abreu, J.M. Use of near infrared reflectance spectroscopy (NIRS) for the prediction of the chemical composition and nutritional attributes of green crop cereals. Anim. Feed. Sci. Technol. 1998, 75, 15–25. [Google Scholar] [CrossRef]
- Caporaso, N.; Whitworth, M.B.; Fisk, I.D. Near-Infrared spectroscopy and hyperspectral imaging for non-destructive quality assessment of cereal grains. Appl. Spectrosc. Rev. 2018, 53, 667–687. [Google Scholar] [CrossRef] [Green Version]
- Dong, W.; Ni, Y.; Kokot, S. A near-infrared reflectance spectroscopy method for direct analysis of several chemical components and properties of fruit, for example, Chinese hawthorn. J. Agric. Food Chem. 2013, 61, 540–546. [Google Scholar] [CrossRef]
- Yi, J.; Sun, Y.; Zhu, Z.; Liu, N.; Lu, J. Near-infrared reflectance spectroscopy for the prediction of chemical composition in walnut kernel. Int. J. Food Prop. 2017, 20, 1633–1642. [Google Scholar] [CrossRef]
- Frank, J.F.; Birth, G.S. Application of Near Infrared Reflectance Spectroscopy to Cheese Analysis. J. Dairy Sci. 1982, 65, 1110–1116. [Google Scholar] [CrossRef]
- Margolies, B.J.; Barbano, D.M. Determination of fat, protein, moisture, and salt content of Cheddar cheese using mid-infrared transmittance spectroscopy. J. Dairy Sci. 2018, 101, 924–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowell, F.E.; Throne, J.E.; Wang, D.; Baker, J.E. Identifying Stored-Grain Insects Using Near-Infrared Spectroscopy. J. Econ. Entomol. 1999, 92, 165–169. [Google Scholar] [CrossRef]
- Johnson, J.B. An overview of near-infrared spectroscopy (NIRS) for the detection of insect pests in stored grains. J. Stored Prod. Res. 2020, 86, 101558. [Google Scholar] [CrossRef]
- Biancolillo, A.; Firmani, P.; Bucci, R.; Magrì, A.; Marini, F. Determination of insect infestation on stored rice by near infrared (NIR) spectroscopy. Microchem. J. 2019, 145, 252–258. [Google Scholar] [CrossRef]
- Beć, K.B.; Grabska, J.; Plewka, N.; Huck, C.W. Insect Protein Content Analysis in Handcrafted Fitness Bars by NIR Spectroscopy. Gaussian Process Regression and Data Fusion for Performance Enhancement of Miniaturized Cost-Effective Consumer-Grade Sensors. Molecules 2021, 26, 6390. [Google Scholar] [CrossRef]
- Mellado-Carretero, J.; García-Gutiérrez, N.; Ferrando, M.; Güell, C.; García-Gonzalo, D.; Lamo-Castellví, S. de Rapid discrimination and classification of edible insect powders using ATR-FTIR spectroscopy combined with multivariate analysis. J. Insects Food Feed. 2020, 6, 141–148. [Google Scholar] [CrossRef]
- Waldbauer, G.P. The Consumption and Utilization of Food by Insects. In Advances in Insect Physiology; Academic Press: London, UK, 1968; Volume 5, pp. 229–288. [Google Scholar]
- Barlocco, N.; Vadell, A.; Ballesteros, F.; Galietta, G.; Cozzolino, D. Predicting intramuscular fat, moisture and Warner-Bratzler shear force in pork muscle using near infrared reflectance spectroscopy. Anim. Sci. 2006, 82, 111–116. [Google Scholar] [CrossRef]
- Chang, C.-W.; Laird, D.A. Near-infrared reflectance spectroscopic analyis of soil C and N. Soil Sci. 2002, 167, 110–116. [Google Scholar] [CrossRef]
- Kröncke, N.; Grebenteuch, S.; Keil, C.; Demtröder, S.; Kroh, L.; Thünemann, A.F.; Benning, R.; Haase, H. Effect of Different Drying Methods on Nutrient Quality of the Yellow Mealworm (Tenebrio molitor L.). Insects 2019, 10, 84. [Google Scholar] [CrossRef] [Green Version]
- Verband Deutscher Landwirtschaftlicher Untersuchungs-und Forschungsanstalten. VDLUFA methodenbuch III. In VDLUFA-Verlag (Vol Ed), Band III-Die Chemische Untersuchung von Futtermitteln; VDLUFA-Verlag: Bonn, Germany, 2013; p. 2190. [Google Scholar]
- Rødbotten, R.; Nilsen, B.; Hildrum, K. Prediction of beef quality attributes from early post mortem near infrared reflectance spectra. Food Chem. 2000, 69, 427–436. [Google Scholar] [CrossRef]
- Leroy, B.; Lambotte, S.; Dotreppe, O.; Lecocq, H.; Istasse, L.; Clinquart, A. Prediction of technological and organoleptic properties of beef Longissimus thoracis from near-infrared reflectance and transmission spectra. Meat Sci. 2004, 66, 45–54. [Google Scholar] [CrossRef]
- Fourty, T.; Baret, F.; Jacquemoud, S.; Schmuck, G.; Verdebout, J. Leaf optical properties with explicit description of its biochemical composition: Direct and inverse problems. Remote Sens. Environ. 1996, 56, 104–117. [Google Scholar] [CrossRef]
- van Huis, A.; Rumpold, B.; Maya, C.; Roos, N. Nutritional Qualities and Enhancement of Edible Insects. Annu. Rev. Nutr. 2021, 41, 551–576. [Google Scholar] [CrossRef] [PubMed]
- Rumpold, B.A.; Schlüter, O.K. Nutritional composition and safety aspects of edible insects. Mol. Nutr. Food Res. 2013, 57, 802–823. [Google Scholar] [CrossRef]
- Urs, K.; Hopkins, T.L. Effect of moisture on growth rate and development of two strains of Tenebrio molitor L. (Coleoptera, Tenebrionidae). J. Stored Prod. Res. 1973, 8, 291–297. [Google Scholar] [CrossRef]
- Machin, J. Water balance in Tenebrio molitor, L. Larvae; the effect of atmospheric water absorption. J. Comp. Physiol. B 1975, 101, 121–132. [Google Scholar] [CrossRef]
- Mellanby, K. Humidity and Insect Metabolism. Nature 1936, 138, 124–125. [Google Scholar] [CrossRef]
- Mellanby, K. The effect of atmospheric humidity on the metabolism of the fasting mealworm (Tenebrio molitor L., Coleoptera). Proc. R. Soc. London. Ser. B, Boil. Sci. 1932, 111, 376–390. [Google Scholar] [CrossRef] [Green Version]
- Viljoen, M.; Hoffman, L.C.; Brand, T.S. Prediction of the chemical composition of mutton with near infrared reflectance spectroscopy. Small Rumin. Res. 2007, 69, 88–94. [Google Scholar] [CrossRef] [Green Version]
- Cozzolino, D.; Murray, I. Effect of Sample Presentation and Animal Muscle Species on the Analysis of Meat by near Infrared Reflectance Spectroscopy. J. Near Infrared Spectrosc. 2002, 10, 37–44. [Google Scholar] [CrossRef]
- Berzaghi, P.; Dalle Zotte, A.; Jansson, L.M.; Andrighetto, I. Near-infrared reflectance spectroscopy as a method to predict chemical composition of breast meat and discriminate between different n-3 feeding sources. Poult. Sci. 2005, 84, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Viljoen, M.; Hoffman, L.C.; Brand, T.S. Prediction of the chemical composition of freeze dried ostrich meat with near infrared reflectance spectroscopy. Meat Sci. 2005, 69, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Alomar, D.; Gallo, C.; Castañeda, M.; Fuchslocher, R. Chemical and discriminant analysis of bovine meat by near infrared reflectance spectroscopy (NIRS). Meat Sci. 2003, 63, 441–450. [Google Scholar] [CrossRef]
- Tøgersen, G.; Arnesen, J.; Nilsen, B.; Hildrum, K. On-line prediction of chemical composition of semi-frozen ground beef by non-invasive NIR spectroscopy. Meat Sci. 2003, 63, 515–523. [Google Scholar] [CrossRef]
- McDevitt, R.M.; Gavin, A.J.; Andrés, S.; Murray, I. The Ability of Visible and near Infrared Reflectance Spectroscopy to Predict the Chemical Composition of Ground Chicken Carcasses and to Discriminate between Carcasses from Different Genotypes. J. Near Infrared Spectrosc. 2005, 13, 109–117. [Google Scholar] [CrossRef]
- Cozzolino, D.; Murray, I.; Scaife, J.R.; Paterson, R. Study of dissected lamb muscles by visible and near infrared reflectance spectroscopy for composition assessment. Anim. Sci. 2000, 70, 417–423. [Google Scholar] [CrossRef]
- Lanza, E. Determination of Moisture, Protein, Fat, and Calories in Raw Pork and Beef By Near Infrared Spectroscopy. J. Food Sci. 1983, 48, 471–474. [Google Scholar] [CrossRef]
- Song, Y.-S.; Kim, M.-W.; Moon, C.; Seo, D.-J.; Han, Y.S.; Jo, Y.H.; Noh, M.Y.; Park, Y.-K.; Kim, S.-A.; Kim, Y.W.; et al. Extraction of chitin and chitosan from larval exuvium and whole body of edible mealworm, Tenebrio molitor. Entomol. Res. 2018, 48, 227–233. [Google Scholar] [CrossRef]
- Janssen, R.H.; Vincken, J.-P.; van den Broek, L.A.M.; Fogliano, V.; Lakemond, C.M.M. Nitrogen-to-Protein Conversion Factors for Three Edible Insects: Tenebrio molitor, Alphitobius diaperinus, and Hermetia illucens. J. Agric. Food Chem. 2017, 65, 2275–2278. [Google Scholar] [CrossRef]
- Song, H.-S.; Lee, K.-T.; Park, S.-M.; Kang, O.-J.; Cheong, H.-S. Measurement of Deproteinization and Deacetylation of Chitin and Chitosan by Near Infrared Spectroscopy. Korean J. Fish. Aquat. Sci. 2003, 36, 88–93. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Tsai, C.-C.; Chen, R.L.; Yang, I.-C.; Hsiao, H.-Y.; Chen, C.-T.; Yang, C.-W. Deacetylation of Chitinous Materials Using Near Infrared Spectroscopy. Eng. Agric. Environ. Food 2008, 1, 33–38. [Google Scholar] [CrossRef]
- Benes, E.; Biró, B.; Fodor, M.; Gere, A. Analysis of wheat flour-insect powder mixtures based on their near infrared spectra. Food Chem. X 2022, 13, 100266. [Google Scholar] [CrossRef] [PubMed]
- Brøndum, J.; Munck, L.; Henckel, P.; Karlsson, A.; Tornberg, E.; Engelsen, S.B. Prediction of water-holding capacity and composition of porcine meat by comparative spectroscopy. Meat Sci. 2000, 55, 177–185. [Google Scholar] [CrossRef]
- Thennadil, S.N.; Martin, E.B. Empirical preprocessing methods and their impact on NIR calibrations: A simulation study. J. Chemom. 2005, 19, 77–89. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Group | Relative Humidity (%) | Water Source | Watering Interval |
|---|---|---|---|
| A | 75 | None | - |
| B | 75 | Pure water (2.6 g) | 1 × per day |
| C | 50 | None | - |
| D | 50 | Pure water (2.6 g) | 1 × per day |
| E | 50 | Carrots (3.0 g) | 1 × per week |
| F | 75 | Pure water (2.6 g) | Every two days |
| G | 75 | Carrots (3.0 g) | 1 × per week |
| H | 75 | Carrots (3.0 g) | Every two days |
| I | 75 | Carrots (3.0 g) | 1 × per day |
| J | 75 | Carrots (10.0 g) | 1 × per week |
| K | 75 | Pure water (2.6 g) and carrots (3.0 g) | 1 × per week and 1 × per day |
| L | 75 | Pure water (2.6 g) and carrots (10.0 g) | 1 × per week and 1 × per day |
| Group | LWGpL (mg) | FCR (-) | ECI (%) | SGR (% per Day) |
|---|---|---|---|---|
| A | 16.0 ± 0.1 g | 5.8 ± 0.2 b | 17.3 ± 0.7 e | 2.4 ± 0.1 e |
| B | 60.5 ± 0.1 a | 2.3 ± 0.0 e | 44.4 ± 0.9 b | 5.1 ± 0.0 a |
| C | 8.5 ± 0.0 h | 8.9 ± 0.2 a | 11.3 ± 0.3 f | 1.5 ± 0.1 f |
| D | 51.9 ± 0.2 c | 2.3 ± 0.0 e | 42.8 ± 0.4 b | 4.8 ± 0.1 a |
| E | 31.0 ± 0.1 f | 2.7 ± 0.1 c | 36.8 ± 0.9 d | 3.6 ± 0.1 d |
| F | 43.8 ± 0.1 d | 2.3 ± 0.1 e | 43.8 ± 1.0 b | 4.4 ± 0.0 b |
| G | 38.7 ± 0.1 e | 2.5 ± 0.1 d | 39.9 ± 1.0 c | 4.1 ± 0.0 c |
| H | 56.2 ± 0.1 b | 1.8 ± 0.0 f | 56.1 ± 1.0 a | 4.9 ± 0.0 a |
| I | 55.4 ± 0.4 b | 1.8 ± 0.1 f | 55.2 ± 3.9 a | 4.9 ± 0.2 a |
| J | 44.7 ± 0.0 d | 2.2 ± 0.0 e | 44.6 ± 0.5 b | 4.4 ± 0.0 b |
| K | 60.1 ± 0.1 a | 2.3 ± 0.1 e | 42.9 ± 1.0 b | 5.1 ± 0.0 a |
| L | 56.6 ± 0.3 b | 2.5 ± 0.1 d | 40.2 ± 1.7 b | 5.0 ± 0.1 a |
| Group | Moisture (%) | Protein (% FW) |
|---|---|---|
| A | 62.7 ± 0.7 c | 22.1 ± 0.1 b |
| B | 72.6 ± 0.6 a | 16.5 ± 0.3 e |
| C | 60.8 ± 0.1 d | 23.5 ± 0.1 a |
| D | 71.5 ± 0.6 a | 17.6 ± 0.6 d |
| E | 67.3 ± 0.5 b | 19.2 ± 0.3 c |
| F | 72.6 ± 2.1 a | 19.4 ± 0.5 c |
| G | 66.5 ± 1.2 b | 18.9 ± 0.5 c |
| H | 71.6 ± 0.7 a | 16.0 ± 0.6 e |
| I | 72.9 ± 0.5 a | 16.5 ± 0.1 e |
| J | 69.2 ± 2.3 ab | 19.5 ± 0.2 c |
| K | 70.4 ± 0.6 a | 18.0 ± 0.2 d |
| L | 71.5 ± 1.2 a | 17.4 ± 0.4 d |
| Statistics | Calibration Set (%) | Validation Set (%) | ||
|---|---|---|---|---|
| Moisture | Protein | Moisture | Protein | |
| n | 120 | 120 | 60 | 60 |
| Mean | 69.1 | 18.7 | 69.2 | 18.8 |
| Minimum | 60.4 | 16.1 | 61.0 | 16.3 |
| Maximum | 72.9 | 23.6 | 73.3 | 23.2 |
| SD | 3.8 | 2.2 | 3.9 | 2.1 |
| Item | Mathematical Treatment | No. of Latent Variables | Calibration Set | Validation Set | |||
|---|---|---|---|---|---|---|---|
| R2c | RMSEC | R2p | RMSEP | RPD | |||
| Moisture | None | 7 | 0.922 | 1.084 | 0.910 | 1.171 | 3.33 |
| MC | 7 | 0.849 | 1.539 | 0.834 | 1.618 | 2.41 | |
| MSC | 5 | 0.966 | 0.701 | 0.966 | 0.711 | 5.49 | |
| 1D | 5 | 0.988 | 0.419 | 0.986 | 0.456 | 8.55 | |
| 2D | 6 | 0.865 | 1.456 | 0.867 | 1.445 | 2.70 | |
| MC + 1D | 4 | 0.953 | 0.829 | 0.953 | 0.832 | 4.69 | |
| MSC + 1D | 4 | 0.952 | 0.837 | 0.949 | 0.868 | 4.49 | |
| Protein | None | 7 | 0.931 | 0.565 | 0.895 | 0.688 | 3.05 |
| MC | 7 | 0.856 | 0.821 | 0.808 | 0.932 | 2.25 | |
| MSC | 5 | 0.944 | 0.506 | 0.915 | 0.611 | 3.44 | |
| 1D | 5 | 0.978 | 0.321 | 0.943 | 0.508 | 4.13 | |
| 2D | 6 | 0.898 | 0.690 | 0.871 | 0.763 | 2.75 | |
| MC + 1D | 4 | 0.940 | 0.525 | 0.916 | 0.607 | 3.46 | |
| MSC + 1D | 4 | 0.913 | 0.631 | 0.892 | 0.683 | 3.08 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kröncke, N.; Benning, R. Determination of Moisture and Protein Content in Living Mealworm Larvae (Tenebrio molitor L.) Using Near-Infrared Reflectance Spectroscopy (NIRS). Insects 2022, 13, 560. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13060560
Kröncke N, Benning R. Determination of Moisture and Protein Content in Living Mealworm Larvae (Tenebrio molitor L.) Using Near-Infrared Reflectance Spectroscopy (NIRS). Insects. 2022; 13(6):560. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13060560
Chicago/Turabian StyleKröncke, Nina, and Rainer Benning. 2022. "Determination of Moisture and Protein Content in Living Mealworm Larvae (Tenebrio molitor L.) Using Near-Infrared Reflectance Spectroscopy (NIRS)" Insects 13, no. 6: 560. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13060560