Effects of the Environmental Temperature on Aedes aegypti and Aedes albopictus Mosquitoes: A Review


1
Department of Biochemistry, Virginia Polytechnic Institute and State University, Blacksburg, VA 24061, USA
2
Institut de Recherche sur la Biologie de l’Insecte, UMR CNRS 7261, Université de Tours, 37200 Tours, France
*
Author to whom correspondence should be addressed.
Insects 2018, 9(4), 158; https://0-doi-org.brum.beds.ac.uk/10.3390/insects9040158
Submission received: 30 September 2018
/
Revised: 25 October 2018
/
Accepted: 31 October 2018
/
Published: 6 November 2018
(This article belongs to the Special Issue Integrative Mosquito Biology: From Molecules to Ecosystems)
{kind=link}
Abstract
:The temperature of the environment is one of the most important abiotic factors affecting the life of insects. As poikilotherms, their body temperature is not constant, and they rely on various strategies to minimize the risk of thermal stress. They have been thus able to colonize a large spectrum of habitats. Mosquitoes, such as Ae. aegypti and Ae. albopictus, vector many pathogens, including dengue, chikungunya, and Zika viruses. The spread of these diseases has become a major global health concern, and it is predicted that climate change will affect the mosquitoes’ distribution, which will allow these insects to bring new pathogens to naïve populations. We synthesize here the current knowledge on the impact of temperature on the mosquito flight activity and host-seeking behavior (1); ecology and dispersion (2); as well as its potential effect on the pathogens themselves and how climate can affect the transmission of some of these pathogens (3).
1. Introduction
The environmental temperature (Ta) is one of the most important abiotic factors influencing the physiology, behavior, ecology, and, by extension, the survival of insects [1,2,3]. Indeed, as poikilotherms, the insect internal temperature varies and depends on the temperature of its surrounding environment. Due to local, daily, and seasonal thermal variations, insects have to face risks including desiccation, changes in metabolism, and even losing the ability to move. However, through evolutionary times, insects have developed various strategies to cope with these thermal variations and avoid thermal stress. Whether they synthesize heat shock proteins, thermoregulate, or adjust their behavioral activity, insects are able to maintain cellular integrity to optimize their fitness and survival [2,4,5].
For each single task (e.g., flying, feeding), the performance of a given insect species is maximal at a given temperature [3]. However, insects can only perform within a range of temperatures; beyond critical minimum and maximum points, their activity is impossible, and the risk of death increases. Several endogenous factors can affect the range of performance, including the physiological state or the age of the insect. The activity range is species-specific, and some have a large spectrum of temperatures at which they can perform (i.e., thermal generalists) while other species have a much narrower window for maintaining their activity (i.e., thermal specialists) [3,6]. In response to the thermal heterogeneity of their environment, in both time and space, insects have developed several methods of adaptation to protect themselves from thermal stress.
These adaptations have allowed insects to colonize almost all kinds of different habitats. For instance, mosquitoes are found worldwide (except in Antarctica) and can live in a large and eclectic array of ecosystems, from tropical forests to urban areas and tundra. These insects can transmit several important diseases to humans and other animals, including malaria, yellow fever, Zika, dengue, and chikungunya, and are responsible for the death of an estimated one million people each year [7]. Among mosquitoes, two species—Aedes aegypti (Linnaeus, 1762), the yellow fever mosquito, and Aedes albopictus (Skuse, 1894), the Asian tiger mosquito—have received a lot of attention in the past decade. For these species, integrated vector management, which is a combination of various method, including vector control and education of the public in order to optimize cost, efficiency, and sustainability [8], is extremely important. Indeed, vaccines have not been developed for several of the viruses transmitted by Ae. aegypti and Ae. albopictus, and vector control remains an essential component of vector management, consisting mainly of space treatment using fogging equipment for these species [9]. Moreover, not only have these species been responsible for the transmission of several pathogens, but their ecological fitting, sensu Janzen [10], has made them extremely invasive across the globe.
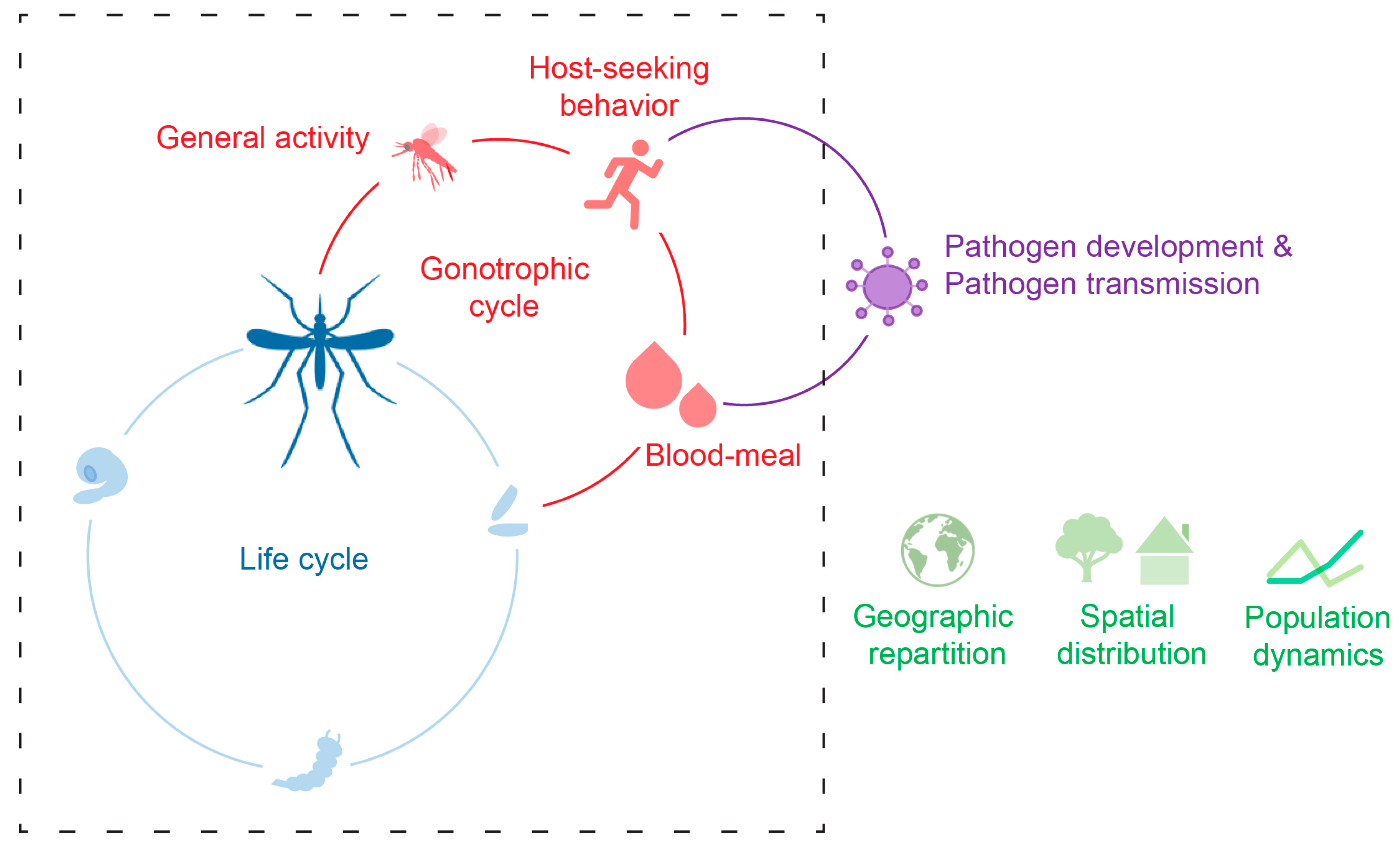
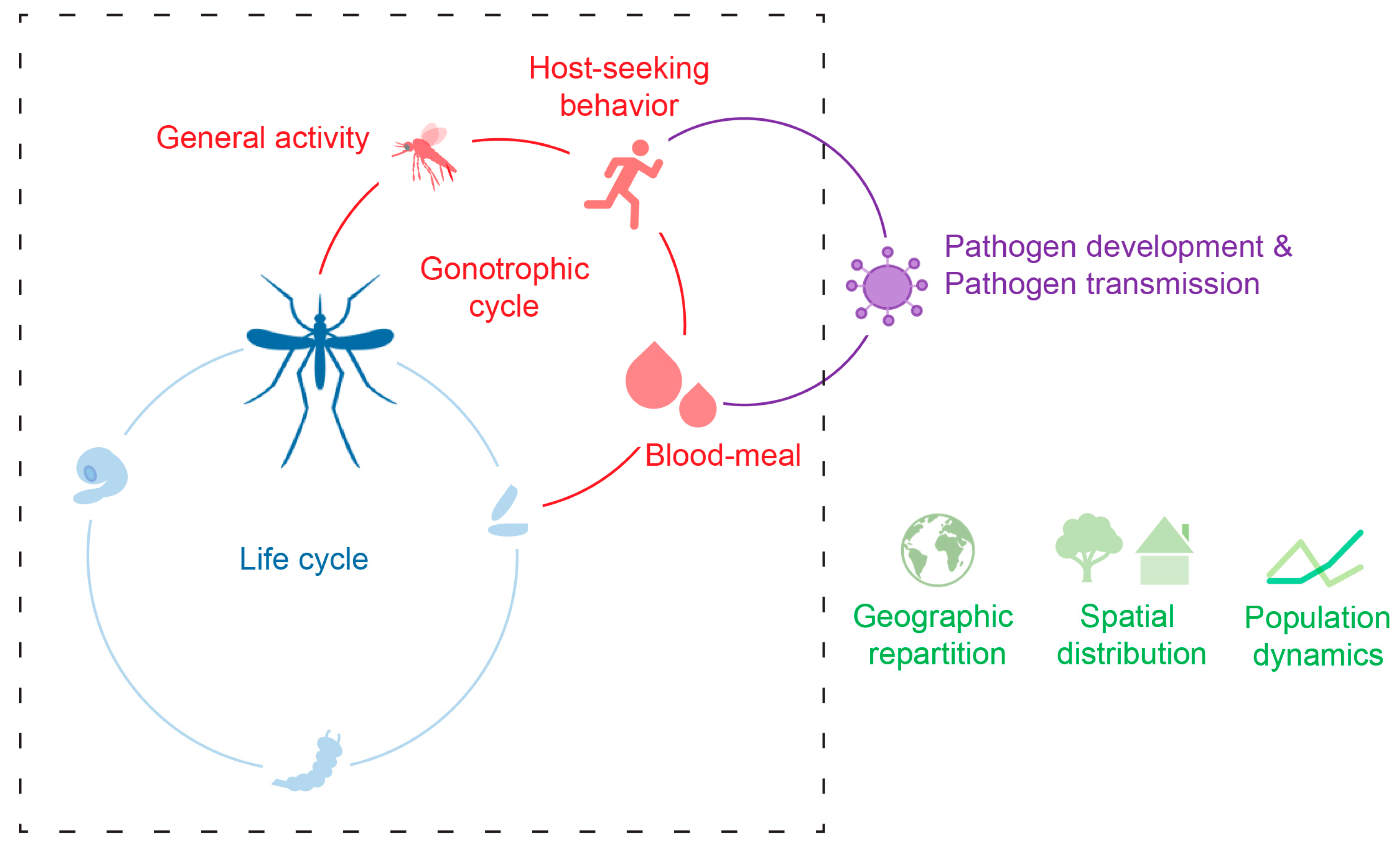
In an environmental context of climate change and global warming [11], which is likely to contribute to the spread of these two species, having a better understanding of how the environmental temperature affects mosquito biology appears essential for decrypting the factors driving the ability of these species to invade new areas, where they could potentially transmit pathogens. The present review compiles the current knowledge on the effect of environmental temperature on Ae. aegypti and Ae. albopictus mosquitoes with a focus on their host-seeking behavior and ecology, including dispersion and vector relevance (Figure 1).
2. Fight Activity, Host-Seeking, and Blood-Feeding
Ae. aegypti and Ae. albopictus are anthropophilic and live in close association with humans, developing preferentially in urban and suburban areas where human hosts are easily available [12,13,14]. Both species are day-biting mosquitoes, exhibiting two main peaks of activity: one in the early morning and the other in the late afternoon [12,13]. Ae. aegypti is endophilic (i.e., taking shelter inside houses) and endophagic (i.e., blood-feeding inside houses), but also moves between indoor and outdoor spaces. Ae. albopictus also exhibits endophilic activity but is considered exophagic (i.e., most biting occurs outside human dwellings), and is an opportunistic feeder, biting a wide range of hosts, from cold-blooded to warm-blooded animals [13].
Since mosquitoes move from inside to outside (or the opposite) shelters and encounter daily and seasonal thermal fluctuations, they might experience an important range of temperatures that can affect their behavior, including host-seeking. The lower temperature limit for Ae. aegypti is around 10 °C, a temperature below which mosquitoes become torpid and unable to move [12,15,16]. Rowley and Graham [17] found that tethered Ae. aegypti females were able to sustainably fly between 15 °C and 32 °C, while flight was possible but only for short periods of time at extreme temperatures such as 10 °C and 35 °C. The optimal flight temperature, in terms of duration and distance flown was determined to be at 21 °C, but overall, the flight performance of Ae. aegypti tethered females was better below 27 °C. The maximum flight speed (34.1 m/min) was recorded at 32 °C/50% humidity. It is important to highlight that the mosquitoes were able to fly at 10 °C (8.9 ± 0.96 m/min) and at 35 °C (18.2 ± 1.98 m/min). The authors argue that flying at lower temperatures allows the species to be active at the cooler hours of the day (i.e., early morning and late afternoon). Christophers [12] also found that female Ae. aegypti wing beat frequency is affected by Ta, measuring 367 beat/s at 18 °C vs. 427 beat/s at 25 °C. Mosquitoes use a wide range of different cues to locate their hosts, including thermal, visual, and chemical information (reviewed in Reference [18]), and host-seeking behavior is tightly linked to their activity (i.e., ability to move and fly). Besides affecting mosquitoes’ general activity and host-seeking behavior, to initiate probing and blood-feeding, a difference between Ta and the host temperature is required [19,20]. Bishop and Gilchrist [21] report a higher percentage of Ae. aegypti females imbibing blood at 42 °C when the difference between Ta and the blood-meal was 14 °C (71%) than when the fluid and Ta were at the same temperature (24 °C: 6%; 28 °C: 8%; 37 °C: 19%). The lower temperature limit at which Ae. aegypti has been found to cease biting is 15 °C, both in the field and experimentally in the lab [22,23,24]. Connor [22] stated that Ae. aegypti is most active at 28 °C. Marchoux et al. [24] found that females fed faster between 26 °C and 35 °C compared to temperatures between 19 °C and 25 °C. The upper temperature limit for blood-feeding is above 36 °C, with the death point being set at 40 °C in Ae. aegypti [12]. Interestingly, comparatively less is known regarding the impact of temperature on the flight activity and host-seeking behavior in Ae. albopictus.
Unlike numerous other mosquito species, Ae. aegypti generally takes several meals during a single gonotrophic cycle [25]. Scott et al. [26] noticed that the frequency of blood intake in this species was positively correlated with Ta (i.e., females fed more often in a warmer environment than in a relatively colder one). However, the authors highlight that this might be due to indirect effects of Ta on the mosquito development, energy storage, and rate of blood-meal digestion and not necessarily due to direct effects on blood-feeding itself [26,27]. Christophers [12] reports that mosquitoes will bite at any temperature at which they are active. In Ae. albopictus, it has been shown that the shortest gonotrophic cycle occurred at 30 °C (3.5 days). At this temperature, the authors also recorded the highest number of cycles (3.9) during the female’s life. However, the total number of eggs laid by females during each gonotrophic cycle does not seem to be different at 20 °C, 25 °C, 30 °C, or 35 °C [28]. In this study, Ae. albopictus was reared at a constant temperature. Studying the impact of fluctuating temperatures on the development of Ae. albopictus, Löwenberg Neto and Navarro-Silva [29] found that when reared under a 27 °C/20 °C thermal regime, the average duration of the females gonotrophic cycle was 11.2 days, with an average of 33.1 eggs laid during the first cycle. In Ae. aegypti, Carrington et al. [30] found that the length of the gonotrophic cycle was reduced with increasing mean temperatures, and the shortest was observed at 26 °C and 30 °C. In Ae. aegypti, a large Diurnal Temperature Range (DTR) (i.e., the range of Ta that mosquitoes experience over a 24-h period) decreases female fecundity, while a small DTR (8 °C) increases female reproduction [31].
3. Ecology and Dispersion
3.1. Thermal Optimum and Lower/Upper Zero Developmental and Survival Temperatures
The temperature of the environment alters mosquito population dynamics by affecting the development of the immature stages (i.e., eggs, larvae, and pupae) as well as reproduction (reviewed by Reference [32]). The lower temperature threshold for Ae. aegypti to develop is 16 °C, while 34 °C is the upper limit [12]. At a lower temperature (i.e., 8 °C), larvae are motionless and die within a couple of days. Couret et al. [33] also showed that food availability as well as density are factors to consider for affecting the larval development rate and survival in this species, in combination with temperature. The development time from hatching to adult emergence was shorter at higher temperatures (30 °C vs. 21 °C) and correlated with density and food availability. Bar-Zeev [34] found that the time taken by larvae to complete their development was optimal at 32 °C and that mortality was significant at 14 °C and 38 °C. The highest temperature at which development fully occurred was 36 °C. The same author reported 40 °C to be the thermal upper limit for Ae. aegypti females, while immature stages were found to survive short exposure to Ta up to 45 °C [35]. Ae. albopictus can develop and survive in a wider range of Ta. Indeed, the lower developmental zero temperature is 10.4 °C, with an optimum of 29.7 °C [28]. This species was shown to fully develop between 15 °C and 35 °C and to survive longer at lower temperatures (15 °C vs. 35 °C) in both females and males. It is interesting to mention that different results have been found for this species across studies, depending on the region of origin of the tested populations and their tolerance to cold. For example, Teng and Apperson [36] showed that the larval development temperature threshold was around 9 °C in the same species.
3.2. Phenology and Population Dynamics
Ae. albopictus occurs in both temperate and tropical areas and has the ability to overwinter as an egg [37] or an adult. Moreover, in sub-tropical regions, they can also lay eggs that will hatch and not enter diapause [38]. Hawley et al. [39] tested the overwintering survival capacity of different strains of Ae. aegypti and Ae. albopictus in Indiana (USA). While none of the Ae. aegypti strains survived, strains of Ae. albopictus acclimated to North American and Asian conditions had a higher survival rate compared to tropical strains. Moreover, within the USA, strains from the south had a lower ability to overwinter than those from northern parts of the country. This highlights the strong capacity of Ae. albopictus to adapt rapidly to new thermal conditions and spread to colder regions [40]. Ae. aegypti is primarily found in tropical and sub-tropical regions, but it is not completely bound by outside temperatures, being the most cosmopolitan species among insect vectors [41]. In this species, continuous breeding is tightly linked to higher Ta, and these mosquitoes will typically not survive winter as an adult under cold climatic conditions but rather overwinter at the egg stage [12,42]. However, an overwintering population of Ae. aegypti adults was recently located near Washington D.C. (USA), which has an average winter temperature lower than what has previously shown to be the limit for the development of this species [43]. The authors argued that Ae. aegypti is taking advantage of a subterranean habitat to survive cold temperatures. In tropical regions, both mosquito species can be found year-round, especially in urban areas. Tsunoda et al. [44] reported that the lowest water temperature (i.e., 14 °C) in artificial containers was above the lower thermal limit for Ae. aegypti and Ae. albopictus development, thus contributing to a yearly occurrence of these mosquitoes and some of the viruses they transmit, such as dengue. Both species’ eggs are resistant to desiccation, allowing them to survive unfavorable conditions and contributing to their ability to spread to new areas via the national and international transportation of materials (e.g., tires, plant pots) containing viable but dormant eggs [41]. Importantly, diurnal and seasonal Ta variations affect the development, density, and dispersion of both species. Carrington et al. [31] used a dynamic model to estimate the thermal limit under which an Ae. aegypti mosquito population may persist and found that both small and large DTRs can affect the population dynamics. Seasonal variations in Ta can greatly impact Ae. aegypti and large DTR (20 °C) negatively impacts survival [45]. Soper [41] highlights that seasonal fluctuations in terms of range and density are observed, especially at the periphery of the species regional distribution. While Ta has an important effect on the population dynamics, rainfall and drought also impact mosquito density and dispersion in both temperate and tropical regions [46]. In Japan, Suwonkerd et al. [47] found that relatively cooler Ta may explain the decline of Ae. albopictus in the dry season. Ae. aegypti populations decreased earlier in the season compared to Ae. albopictus, but this was not associated with the co-occurrence of the two species at the different sites. Mogi [48] studied several mosquito populations in the same country and highlighted that warmer temperatures are likely to affect overwintering strategies and pathogen transmission by modifying both mosquito biting activity and distribution as well as pathogen development. Beyond responding to changes of thermal conditions, mosquito population dynamics are affected by several other factors including photoperiod (i.e., short days inducing diapause). Tsunoda et al. [49] showed that day length affects egg hatching in Ae. albopictus and suggested that under milder (i.e., tropical) conditions, overwintering is not as prevalent in populations in temperate areas.
3.3. Spatial Distribution and Ta Variability
Ae. albopictus is capable of surviving in much colder climates than Ae. aegypti, and this species has also adapted well to urban environments. Tsuda et al. [50] showed that Ae. aegypti was more commonly found in urban areas while Ae. albopictus was found more in rural areas in Thailand, but the pattern of precipitation during their two field seasons was not different between the two sites. If no mention is made regarding Ta, it can be hypothesized that an urban habitat has a higher Ta compared to a rural area, where vegetation is more abundant. Ae. albopictus is also mainly exophagic, which means that air conditioning in the summer would not have as much of an effect on it, as it would on Ae. aegypti [51]. Ae. aegypti can be endophilic and endophagic, allowing this species to reap the benefits of controlled indoor temperatures in extreme heat and cold as well as utilizing man-made structures filled with rain water for oviposition, which allows this species to thrive in urban environments [52]. This highlights that Ae. albopictus has developed several strategies to cope with a wider range of Ta and adapt to local thermal conditions. Moreover, it has been shown that environmental variability can actually promote the occurrence of the sudden increased abundance of Ae. aegypti [53,54]. With higher Ta in the summer, the yearly pattern of distribution of Ae. aegypti in temperate and sub-tropical areas is indeed greatly affected [42]. Applying Schmalhausen’s law [55] to explain both the ecological and evolutionary consequences of a fluctuating environment, in particular in Ae. albopictus, Chaves [56] highlighted that perturbations within the normal thermal range have little effect under abnormal or stress conditions, but small perturbations have a stronger effect on individuals. In the same species, Chaves [57] showed that spatial and temporal abundance patterns are most affected by temperature and precipitation, respectively, and that co-occurrence with other species, in this case Ae. japonicus and Ae. flavopictus, also has major consequences on population dynamics and distribution. Higa et al. [58], in their study on the geographical distribution of these mosquitoes along a north-south transect in Vietnam, showed that Ta affects, in combination with rainfall, the density of both mosquito species. They also mention that in regions where both species were found, climatic conditions are considered milder and habitat heterogeneity becomes a much more important factor explaining spatial distribution.
3.4. Vector Distribution in the Context of Climate Change
A recent study by Kraemer et al. [59] reported that the global distribution of both vectors was never considered as important in the past as it is today. Ae. aegypti and Ae. albopictus have been able to greatly expand their geographical distribution worldwide in the past 30 years, and several studies have shown that climate change is likely to impact the range of these two species [60,61]. For example, Kearney et al. [62] studied the impact of climate change on the distribution and abundance of Ae. aegypti in Australia using biophysical models and evolutionary theory and found that water availability, egg desiccation, and tolerance to colder Ta are important factors with the potential to drive the establishment of these mosquitoes in new regions in the south and north of the country. Alto and Juiliano [63] investigated how temperature and precipitation regime affect Ae. albopictus. A relatively high Ta (i.e., 30 °C) was more favorable to adult development, both in terms of total number of individuals completing their development and time to adult emergence. This study also showed that in a mild environment (i.e., 22 °C), this species is still able to produce offspring and develop; this could contribute to the colonization of northern (i.e., cooler) regions of the USA. Yang et al. [64] used mathematical modeling (i.e., population dynamics theory) to determine that 29.2 °C was the optimal temperature to produce the highest amount of offspring in Ae. aegypti mosquitoes and maintain a viable mosquito population.
Several studies have also highlighted the impact of environmental conditions on the co-occurrence and inter-species competition between Ae. aegypti and Ae. albopictus. Since its arrival in the USA, Ae. albopictus has led to the decline of Ae. aegypti populations in several regions [65]. Lounibos et al. [66] compared the development and survival rates of both species when maintained at different Ta, in combination with other factors including food availability, and found that larval competition outcome was limited between 24 °C and 30 °C and that survival rate was unaffected. Kobayashi et al. [67] used a geographical information system (GIS) and larval survey to analyze the distribution of Ae. albopictus in Japan and predict its future expansion both in Japan and the USA. They showed an important correlation between the annual mean temperature and the density of Ae. albopictus and revealed that this species is moving further north. They estimated that its current range is also correlated with an average annual Ta of 11 °C in both countries, and that global warming will allow Ae. albopictus to expand northward. Mogi and Tuno [68] studied Ae. albopictus in its native geographical range using a retrospective approach, including local thermal conditions that have allowed this species to change its pattern of distribution in Japan. They showed that higher Ta during the winter was one important factor that had driven the expansion of the geographical range of Ae. albopictus.
Climate change is expected to result in major shifts in vector distribution and/or in the expansion of geographical ranges of both mosquito species with a potential health impact on local populations of humans and other animals due to an enhanced transmission rate of pathogens, including dengue and Zika [46,69,70,71,72]. Rochlin et al. [73] estimated that in the context of climate change, regions with suitable environmental conditions for the development of Ae. albopictus will increase by 50% by the end of the century, placing another 30 million people at risk.
4. Pathogen Transmission
The feeding and living habits of Ae. aegypti and Ae. albopictus make these species efficient vectors for human diseases. An arthropod is considered to be a competent vector of an arbovirus or other pathogens if the species demonstrates that it can naturally acquire, be infected by, and transmit a pathogen [74]. This is true for Ae. aegypti and Ae. albopictus, since they meet these criteria for several arboviruses [75,76]. While Ae. aegypti is generally considered a more competent vector for several arboviruses, such as dengue, chikungunya, Zika, yellow fever, and Mayaro, Ae. albopictus can vector many of the same viruses with less competence but with the ability to expand its range into colder regions [77]. Ae. albopictus also transmits diseases such as West Nile virus, eastern equine encephalitis, and dirofilariasis. The pathogens these species transmit overlap, which could allow some diseases to spread beyond the range of one vector with the help of the other. The ability of these species to vector these pathogens depends on two main factors: whether the pathogen can infect the midgut cells and whether the pathogen can disseminate in order for the vector to transmit it to a new host. Temperature can affect both of these factors, in addition to the development of the mosquito and the time between infection by the pathogen and its transmission (extrinsic incubation period) [57]. The effects of climate change allow these species to develop the pathogens more quickly and spread into new areas, carrying pathogens with them [78,79,80].
4.1. Dengue Virus Complex
The dengue virus complex, which refers to the four serotypes that cause dengue fever, has become a major worldwide concern due to its rapid spread and increased intensity in endemic areas [81]. According to the World Health Organization, this is the fastest spreading arbovirus; estimates show that 390 million people are currently infected per year, while 3.9 billion are considered at risk [82,83]. The serotypes can cause different severities of the virus in different people, ranging from asymptomatic to hemorrhagic fever to severe dengue, the latter usually occurring after infection of multiple serotypes, which can lead to coma or death [81]. Vaccines to prevent dengue infections are being developed, with one currently on the market, but only individuals who are seropositive are recommended to receive the vaccine. Because of the limited use of this vaccine, vector control is still the main method to prevent the spread of this virus. Eradication was achieved in the Americas in the 1950’s and 1960’s in an attempt to reduce yellow fever outbreaks, but Ae. aegypti returned in the 1970’s, leading to endemic outbreaks of dengue [52]. Since the 1970’s, dengue presence has expanded from nine countries to being endemic in over 100 [81].
Ae. aegypti is currently the primary vector, though Ae. albopictus can also act as a vector, especially in regions where Ae. aegypti is not present [75]. Lambrechts et al. [45] showed that a higher DTR (20 °C)—which would be seen in temperate climates in spring and autumn—has a negative effect on the vector competence for dengue and the survival of Ae. aegypti compared to a low DTR (10 °C), such as the temperature ranges typically experienced in summer. As discussed previously, temperature has an effect on the blood-feeding habits and survival of Ae. aegypti and potentially Ae. albopictus, which would both inherently impact pathogen transmission. Ae. aegypti is endophilic and endophagic, rendering this species capable of expanding into regions outside its typical temperature range, meaning it can bring dengue along to new regions, making predictions of dengue dispersion a difficult task [52]. The effect of temperature on the development of Ae. aegypti was studied by Couret and Benedict [32], who showed that increased temperatures had a positive impact on the rate of development. Global temperature increases would allow for a greater distribution range, as well as an increase in virus transmission. Watts et al. [84] showed that the extrinsic incubation period (EIP)—the amount of time between the mosquito taking an infected blood-meal to transmitting the disease to the next vertebrate host—for DEN2 in Ae. aegypti decreases with higher temperatures (32–35 °C) by several days, potentially allowing for faster transmission as one of the consequences of global warming. An experiment was conducted by Rohani et al. [85] for DEN2 and DEN4 serotypes with similar results. Ae. aegypti was also shown to be less likely to disseminate dengue at lower temperatures [86].
Because of the ability of Ae. albopictus to survive in colder climates, carry pathogens over the winter due to diapause, and have exophagic tendencies, Ae. albopictus does have some abilities for transmitting pathogens that Ae. aegypti does not [51,75]. Brady et al. [77] highlighted that the longer lifespan of Ae. albopictus causes its vector potential to exceed that of Ae. aegypti. However, Whitehorn et al. [87] found that Ae. albopictus is as susceptible to dengue as Ae. aegypti and can even carry a higher viral burden, but DEN2 and DEN4 are not found in high numbers in the saliva, indicating a lower transmission potential for these serotypes. Ae. albopictus is not usually found in nature with dengue in areas where Ae. aegypti is present, though it has been shown to transmit it in the lab as well as in regions where Ae. aegypti is not present. Ae. albopictus is thought to mainly be a secondary vector to maintain dengue in rural areas [65]. For this reason, Ae. albopictus is considered to be less important as a vector for dengue, as compared to Ae aegypti.
Mathematical modeling has shown a clear dependence on seasonal variation of dengue incidence [64]. This seasonality is often assumed to be due to only vector presence, which is typically worse during warm and rainy seasons. However, Thu et al. [88] found that the propagation of dengue within Ae. aegypti increased with humidity, with a preferred humidity over 60% and preferred a Ta range of around 24–31 °C. This shows that the amplification of the virus in the mosquito seems to be optimal in conditions that are also favorable to mosquitoes.
4.2. Chikungunya Virus
Chikungunya is an alphavirus that is typically found in Africa and Asia, although it has been spreading and has caused outbreaks in over 60 countries in recent years [89]. Though cases of chikungunya are relatively low compared to dengue, outbreaks in naïve populations can be devastating. The disease can be difficult to track, since it is often mistaken for dengue due to their shared symptoms, vectors, and distribution. Both chikungunya and dengue cause flu-like symptoms, such as fever, headache, and nausea, but chikungunya can cause severe joint pain and rarely causes death, with the exception of a recent outbreak in La Réunion [90]. The primary vector is Ae. aegypti and the secondary is Ae. albopictus for both chikungunya and dengue, which means that both viruses occur in areas where Ae. aegypti and/or Ae. albopictus are present [89]. It is thought that most of the spread from the original endemic regions in Asia and Africa is caused by Ae. albopictus. A major outbreak in La Réunion, an island off the coast of Madagascar, brought chikungunya to the attention of the world in 2005 and 2006. The number of reported cases exceeded 47,000 in a population of about 776,000, which included over 50 cases of materno-neonatal transmission and chikungunya playing a role in the death of over 200 people [90]. These numbers only include those that expressed clinical symptoms, which could only account for as low as 20% of the actual cases [91], and some protective immunity could occur in endemic regions [90]. The first instance of chikungunya in Europe was in Italy in 2007, shortly following the invasion of Ae. albopictus [92]. The 2007 outbreak revealed a new strain of chikungunya virus that had adapted to being vectored by Ae. albopictus. This mutation (E1-A226 V) gives the virus an advantage for both replication and transmission in Ae. albopictus [93]. However, according to Zouache et al. [94], the mutation allowing Ae. albopictus to transmit chikungunya may only be advantageous in different environments. This is due to the interaction between the mosquito population, the genetics of the virus, as well as environmental temperature, which means that temperature plays a large role in transmission of chikungunya. The outbreak in 2007 also showed that Ae. albopictus has the potential to bring diseases with it to any area to which it can expand [51], and there is a possibility that transovarial and venereal transmission in mosquitoes may lead to a resurgence of the disease [95,96,97]. The first case of chikungunya in the Americas occurred in 2013 on the Caribbean island of St. Martin, and the virus spread to Central and South America and the southern USA in 2014 [98,99]. Local transmission has not been reported in the USA since 2015, but imported cases still occur in several states each year [100]. No vaccine is currently available to prevent chikungunya, so vector control and personal protection from mosquito bites are the main methods for avoiding disease transmission. The spread of the disease over the last decade to new and higher latitudes is at present a cause for concern [89].
4.3. Zika Virus
Zika virus also belongs to the family flavivirus and resembles dengue so much that it could almost be referred to as the “fifth dengue serotype”, since serotypes are based on the cross-neutralization of antibodies, and Zika has the potential to neutralize dengue antibodies [101]. Zika is vectored mainly by Ae. aegypti in the Americas, as well as other Aedes species [102,103], but it can also be transmitted through sexual intercourse and from mother to fetus. The disease has gained interest over the last several years when it was linked to microcephaly and other complications in infants born to mothers who were infected by Zika during pregnancy [102]. Typically, only one out of every five people who are infected with Zika displays symptoms, which include fever, rash, muscle and joint pain, sinus congestion, and other flu-like symptoms [102]. Zika was first seen in humans in Uganda in 1952, and, until 2007, it was primarily endemic in countries of Africa and Asia. The first instance outside of the endemic areas occurred on the Island of Yap [104], and this strain continued to spread from there into French Polynesia, causing a large outbreak in 2013, and moved to Brazil in 2015 [105,106]. Shortly after Zika began spreading in Brazil, large increases in microcephaly in newborns and Guillain-Barré syndrome in adults were observed, being quickly connected to Zika. It has since spread across South America, and autochthonous transmission cases have been reported in more than 80 countries worldwide [102,107]. There is unfortunately no evidence about how Zika progresses in different temperatures, which means the main way to track its possible impact and spread is through understanding its vectors [108]. Though some studies have shown that other Aedes species, such as Ae. vexans, can vector Zika [103,109,110], there is still little evidence for the vector competence of species other than Ae. aegypti, such as Ae. albopictus, which could significantly change the possible range of Zika’s impact [111,112]. There is currently no vaccine or treatment for Zika virus, so vector control remains critical [102].
4.4. West Nile Virus
West Nile virus is another member of the flavivirus family, though it is more closely related to Japanese encephalitis and St. Louis encephalitis than to dengue and Zika [51]. West Nile virus can be very severe and even fatal to those who display symptoms, though only about 20% of people that are infected will show any signs. The severe symptoms are referred to as West Nile fever, which is a febrile disease with nausea, vomiting, body aches, and headaches, sometimes with a rash. Severe West Nile disease occurs in about 0.67% of those infected and contains the symptoms of West Nile fever with additional neurological problems, which could lead to coma and death [113]. West Nile virus mainly affects humans and horses, with birds and other non-mammals, such as alligators, acting as reservoirs and amplifiers for the disease [113,114]. The virus was first seen in Uganda in 1937 and has been circulating in Africa, the Middle East, Europe, and Australia for years. A strain of West Nile virus from Israel and Tunisia was brought into the USA via New York in 1999 and was picked up by native mosquitoes, causing a large outbreak [115]. The virus is now also considered endemic to North America [113]. West Nile virus can be vectored by several different types of mosquitoes, including species in the genera Culex (with Cx. pipiens as the main vector), Ochlerotatus, and Aedes, including Ae. albopictus [113,116]. Ae. albopictus has been shown to be susceptible to infection and easily disseminated West Nile virus, while Ae. aegypti did not show susceptibility [116]. Ae. albopictus has also proven to be a natural vector in the wild and could possibly act as a bridge vector from enzootic cycles to urban cycles due to its opportunistic feeding habits [75,117]. Like most arboviruses, seasonality has been observed in West Nile virus, which is in part due to vector density increasing during rainy and warm months, but it is also related to the development of the pathogen in the mosquito. Similar to dengue, infection and dissemination rates (tested in Cx. pipiens quinquefasciatus) increased with temperature, recording the lowest rates at 25 °C and the highest at 30 °C [118]. Dohm et al. [119] also found that dissemination rates were less than 30% for mosquitoes tested with an extrinsic incubation temperature of 18 °C, with the highest dissemination rate at 30 °C. These studies showed that relatively higher temperatures positively affect the amplification of West Nile virus in the vector, allowing for higher transmission rates in warmer environments.
4.5. Yellow Fever Virus
Yellow fever is another arbovirus in the family of flaviviruses. This virus does not always cause symptoms, but when it does, they can present in two stages. The first stage is moderate, which is characterized by fever, muscle pain, and vomiting, and the second is more severe and includes jaundice, renal failure, and bleeding. The more severe forms may be fatal within about a week. Because there is no cure for the disease, and it can cause such serious cases, it is important to control yellow fever virus as much as possible [120]. Yellow fever virus is now endemic to several countries in Africa and South America, but it caused widespread epidemics when it reached the Americas and Europe in the late 17th century. It circulates in nature in two cycles: a sylvatic in monkeys and an urban cycle in humans. Ae. aegypti is the main vector for the urban cycle, and Ae. albopictus is thought to bridge the gaps between the sylvatic and urban cycles due to its peri-domestic, opportunistic feeding habits [121]. The existence of the sylvatic cycle makes control of the disease more difficult, as does transovarial vertical transmission in mosquitoes, since it is maintained in different primates and different mosquitoes, such as Ae. africanus, Haemagogus spp., and Sabethes spp. [121,122,123]. Because the disease is maintained outside of the urban cycle, bridge vectors like Ae. albopictus can carry the disease back into the urban cycle, which can lead to outbreaks. Yellow fever virus was almost completely eradicated from the Americas in the mid-1920’s, but the disease re-emerged from the sylvatic cycle [124]. There is a highly efficient vaccine available for endemic areas, but this virus can be devastating to naïve populations, and outbreaks can still occur when the coverage of vaccination is not wide or quick enough, or if vector control is not implemented [120,121].
The spread of yellow fever virus is known to be affected by temperature. Davis [125] found that the EIP in Ae. aegypti decreased with an increase in temperature. The mosquitoes were able to transmit the disease in as short as four days after infection at 37 °C, while they could not transmit it at 18 °C or less, unless the temperature later increased. These data show that yellow fever virus can be transmitted after a shorter time in warmer weather. Although yellow fever virus has caused epidemics in typically temperate climates, these outbreaks would be seasonal in nature due to the effect temperature has on the virus. It is therefore important to focus on tropical and sub-tropical areas where yellow fever is endemic with strategies for both vector control and widespread use of vaccines.
4.6. Other Pathogens
In addition to the above-named viruses, Ae. aegypti and Ae. albopictus act as vectors for several other diseases. Ae. albopictus has been shown to be able to experimentally vector over 20 viruses, though there is no field evidence supporting the natural transmission of all of them [126].
Aedes albopictus is a potential bridge vector for eastern equine encephalitis (EEE), which is a virus that affects both humans and horses. EEE has been isolated from Ae. albopictus in Florida, USA [127]. Komar et al. [128] found that, when experimentally infecting Aedes mosquitoes with EEE, 100% of blood-fed Ae. albopictus were infected, compared to 40% of Ae. aegypti. Although climate change would have an effect on the distribution of the vector, Chamberlain and Sudia [129] found that temperature had little effect on the development and amplification of the virus in the vector, another Aedes species (Ae. triseriatus), though lower temperatures caused lower transmission rates and short exposures to high temperatures elongated the incubation period and decreased the rate of transmission.
Mayaro is a virus very similar to chikungunya, which exists in a sylvatic cycle in South America. However, cases have been seen closer to urban areas, and Ae. aegypti has been shown to experimentally transmit the virus [51]. A case of a young boy in Haiti who was found to be positive for both Mayaro and dengue showed that Mayaro may have serious public health implications [130]. da Costa Carvalho and Fournier [131] showed that temperature has an effect on the virus when Mayaro virus is subjected to heat stress (37 °C) in Ae. albopictus cells, and the virus halts translation of its own proteins, allowing the host cell to synthesize heat shock proteins. According to Hotez and Murray [132], Mayaro may have the potential to spread through the Americas, much like dengue and chikungunya.
Some strains of Ae. albopictus and Ae. aegypti also have the ability to transmit parasitic diseases, such as dirofilariasis and elephantiasis, with Ae. albopictus being the more competent vector [75,133,134]. The most common dirofilarial worm transmitted by these mosquitoes is Dirofilaria immitis, otherwise known as “heartworm”. This disease can be fatal to canines if left untreated, and immiticide treatments are unpleasant and expensive. The best option is preventing this disease using prophylactic treatments and vector control. Humans can also be infected with dirofilaria, which is mainly endemic to southern Europe, but is spreading north [135]. The seasonality for dirofilariasis, as well as its geographic range, is expanding due to rising global temperatures, allowing for a longer transmission season [136]. Ledesma and Harrington [137] also show that D. immitis L3s are significantly affected by temperature fluctuations during the EIP, developing more quickly under fluctuating temperatures than constant temperature within a certain range. Dirofilariids show little preference for vector species compared to some other pathogens, meaning that these worms could be easily introduced to a new region if a competent vector is present. The presence of invasive competent vectors, such as Ae. albopictus, in endemic areas can increase the transmission of the disease [138]. Therefore, it is important to understand to what extent changing temperatures would allow the expansion of Ae. albopictus and Ae. aegypti distribution range, in order to control for this and other diseases.
5. Conclusions, Future Directions and Knowledge Gaps
As Ae. albopictus and Ae. aegypti continue to expand their distribution range, and in the context of global warming, it appears more important than ever to have a good understanding of their biology and ecology. If much is known about the effects of Ta on Ae. aegypti, comparatively less information is available for Ae. albopictus. Focusing on this knowledge gap, in particular regarding the effects on the activity and host-seeking behavior of this species of mosquito, is however critical to implement accurate data in population dynamics models to determine future distribution of this disease vector species. Extensive work has been conducted in Anopheles mosquitoes and the pathogens they transmit, including Plasmodium falciparum, the agent of malaria (e.g., [139,140,141]). However, little is known regarding the effects of temperature on several viruses including chikungunya and Zika viruses (independent of vector or within vector), and these data would allow predictions to be more precise and realistic. It would also be important to know the vectors of diseases, especially emerging infectious diseases like Zika, in order to understand the potential range and build accurate models [111]. More information becoming available for these mosquitoes and the pathogens they vector will allow for a better understanding of what can be done to prevent the spread of disease.
Author Contributions
Writing—original draft preparation, J.M.R., C.R.L. and C.L.; Writing—review and editing, J.M.R., C.R.L. and C.L.; Supervision—C.L.
Funding
Biochemistry Department, Virginia Tech (USA); University of Tours and CNRS (France).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Denlinger, D.L.; Yocum, G.D. Physiology of heat sensitivity. In Temperature Sensitivity in Insects and Application in Integrated Pest Management; Hallman, G.J., Denlinger, D.L., Eds.; Westview Press: Boulder, CO, USA; Oxford, UK, 1998; pp. 7–53. [Google Scholar]
- Heinrich, B. The Hot-Blooded Insects: Strategies and Mechanisms of Thermoregulation; Harvard University Press: Cambridge, MA, USA, 1993; p. 600. [Google Scholar]
- Huey, R.B.; Stevenson, R.D. Integrating thermal physiology and ecology of ectotherms: A discussion of approaches. Am. Zool. 1979, 19, 357–366. [Google Scholar] [CrossRef]
- Benoit, J.B.; Lopez-Martinez, G.; Patrick, K.R.; Phillips, Z.P.; Krause, T.B.; Denlinger, D.L. Drinking a hot blood meal elicits a protective heat shock response in mosquitoes. Proc. Natl. Acad. Sci. USA 2011. [Google Scholar] [CrossRef] [PubMed]
- Lahondère, C.; Lazzari, C.R. Mosquitoes cool down during blood feeding to avoid overheating. Curr. Biol. 2012, 22, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Angilletta, M.J. Thermal Adaptation: A Theoretical and Empirical Synthesis; Oxford University Press: New York, NY, USA, 2009; p. 304. [Google Scholar]
- World Health Statistics (WHO). Monitoring Health for the SDGs, Sustainable Development Goals; Licence: CC BY-NC-SA 3.0 IGO; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- World Health Organization. Handbook for Integrated Vector Management; World Health Organization: Geneva, Switzerland, 2012. [Google Scholar]
- Matthews, G. Integrated Vector Management: Controlling Vectors of Malaria and Other Insect Vector Borne Diseases; John Wiley & Sons: Hoboken, NJ, USA, 2011. [Google Scholar]
- Janzen, D.H. On ecological fitting. Oikos 1985, 45, 308–310. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2014: Synthesis Report; Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014; 151p. [Google Scholar]
- Christophers, S.R. Aedes aegypti (L.) the Yellow Fever Mosquito; Cambridge University Press: London, UK, 1960. [Google Scholar]
- Delatte, H.; Desvars, A.; Bouétard, A.; Bord, S.; Gimonneau, G.; Vourc’h, G.; Fontenille, D. Blood-feeding behavior of Aedes albopictus, a vector of Chikungunya on La Réunion. Vector-Borne Zoonotic 2010, 10, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Harrington, L.C.; Edman, J.D.; Scott, T.W. Why do female Aedes aegypti (Diptera: Culicidae) feed preferentially and frequently on human blood? J. Med. Entomol. 2001, 38, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.J. Observations on Aedes aegypti, L. (Dipt. Culic.) under controlled atmospheric conditions. B Entomol. Res. 1933, 24, 363–372. [Google Scholar] [CrossRef]
- Otto, M.; Neumann, R.O. Studien über Gelbfieber in Brasilien. Z. Hyg. InfektKr. 1905, 51, 357–506. [Google Scholar] [CrossRef]
- Rowley, W.A.; Graham, C.L. The effect of temperature and relative humidity on the flight performance of female Aedes aegypti. J. Insect Physiol. 1968, 14, 1251–1257. [Google Scholar] [CrossRef]
- Bowen, M.F. The sensory physiology of host-seeking behavior in mosquitoes. Annu. Rev. Entomol. 1991, 36, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Corfas, R.A.; Vosshall, L.B. The cation channel TRPA1 tunes mosquito thermotaxis to host temperatures. eLife 2015, 4, e11750. [Google Scholar] [CrossRef] [PubMed]
- Zermoglio, P.F.; Robuchon, E.; Leonardi, M.S.; Chandre, F.; Lazzari, C.R. What does heat tell a mosquito? Characterization of the orientation behaviour of Aedes aegypti towards heat sources. J. Insect Physiol. 2017, 100, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Bishop, A.; Gilchrist, B.M. Experiments upon the feeding of Aedes aegypti through animal membranes with a view to applying this method to the chemotherapy of malaria. Parasitology 1946, 37, 85–100. [Google Scholar] [CrossRef] [PubMed]
- Connor, M.E. Suggestions for Developing a Campaign to Control Yellow Fever. Am. J. Trop. Med. Hyg. 1924, 1, 277–307. [Google Scholar] [CrossRef]
- Cossio, V. Observations sobre al Aedes aegypti (Stegomyia) mosquito de la febbre amarilla en Montevideo. Bol. Cons. Nat. Hig. Uruguay 1931, 23, 1664. [Google Scholar]
- Marchoux, E.; Salimbeni, A.T.; Simond, P.L. La Fièvre Jaune: Rapport de la Mission Française; Annales de l’Institut Pasteur: Paris, France, 1903. [Google Scholar]
- Scott, T.W.; Clark, G.G.; Amerasinghe, P.H.; Lorenz, L.H.; Reiter, P.; Edman, J.D. Detection of multiple blood feeding patterns in Aedes aegypti (Diptera: Culicidae) during a single gonotrophic cycle using a histological technique. J. Med. Entomol. 1993, 30, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Scott, T.W.; Amerasinghe, P.H.; Morrison, A.C.; Lorenz, L.H.; Clark, G.G.; Strickman, D.; Kittayapong, P.; Edman, J.D. Longitudinal studies of Aedes aegypti (Diptera: Culicidae) in Thailand and Puerto Rico: Blood feeding frequency. J. Med. Entomol. 2000, 37, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Yasuno, M.; Pant, C. Seasonal changes in biting and larval infestation rates of Aedes aegypti in Bangkok, Thailand in 1969. Bull. WHO 1970, 43, 319–325. [Google Scholar] [PubMed]
- Delatte, H.; Gimonneau, G.; Triboire, A.; Fontenille, D. Influence of temperature on immature development, survival, longevity, fecundity, and gonotrophic cycles of Aedes albopictus, vector of chikungunya and dengue in the Indian Ocean. J. Med. Entomol. 2009, 46, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Löwenberg Neto, P.; Navarro-Silva, M.A. Development, longevity, gonotrophic cycle and oviposition of Aedes albopictus Skuse (Diptera: Culicidae) under cyclic temperatures. Neotrop. Entomol. 2004, 33, 29–33. [Google Scholar] [CrossRef]
- Carrington, L.B.; Armijos, M.V.; Lambrechts, L.; Barker, C.M.; Scott, T.W. Effects of fluctuating daily temperatures at critical thermal extremes on Aedes aegypti life-history traits. PLoS ONE 2013, 8, e58824. [Google Scholar] [CrossRef] [PubMed]
- Carrington, L.B.; Seifert, S.N.; Willits, N.H.; Lambrechts, L.; Scott, T.W. Large diurnal temperature fluctuations negatively influence Aedes aegypti (Diptera: Culicidae) life history traits. J. Med. Entomol. 2013, 50, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Couret, J.; Benedict, M.Q. A meta-analysis of the factors influencing development rate variation in Aedes aegypti (Diptera: Culicidae). BMC Ecol. 2014, 14, 3. [Google Scholar] [CrossRef] [PubMed]
- Couret, J.; Dotson, E.; Benedict, M.Q. Temperature, larval diet, and density effects on development rate and survival of Aedes aegypti (Diptera: Culicidae). PLoS ONE 2014, 9, e87468. [Google Scholar] [CrossRef] [PubMed]
- Bar-Zeev, M. The effect of temperature on the growth rate and survival of the immature stages of Aedes aegypti (L.). Bull. Entomol. Res. 1958, 49, 157–163. [Google Scholar] [CrossRef]
- Bar-Zeev, M. The effect of extreme temperatures on different stages of Aedes aegypti (L.). Bull. Entomol. Res. 1957, 48, 593–599. [Google Scholar] [CrossRef]
- Teng, H.J.; Apperson, C.S. Development and survival of immature Aedes albopictus and Aedes triseriatus (Diptera: Culicidae) in the laboratory: Effects of density, food, and competition on response to temperature. J. Med. Entomol. 2000, 37, 40–52. [Google Scholar] [CrossRef] [PubMed]
- Mori, A.; Oda, T.; Wada, Y. Studies on the egg diapause and overwintering of Aedes albopictus in Nagasaki. Trop. Med. 1981, 23, 79–90. [Google Scholar]
- Higa, Y.; Toma, T.; Araki, Y.; Onodera, I.; Miyagi, I. Seasonal changes in oviposition activity, hatching and embryonation rates of eggs of Aedes albopictus (Diptera: Culicidae) on three islands of the Ryukyu Archipelago, Japan. Med. Entomol. Zool. 2007, 58, 1–10. [Google Scholar] [CrossRef]
- Hawley, W.A.; Pumpuni, C.B.; Brady, R.H.; Craig, G.B., Jr. Overwintering survival of Aedes albopictus (Diptera: Culicidae) eggs in Indiana. J. Med. Entomol. 1989, 26, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Hanson, S.M.; Craig, G.B., Jr. Cold acclimation, diapause, and geographic origin affect cold hardiness in eggs of Aedes albopictus (Diptera: Culicidae). J. Med. Entomol. 1994, 31, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Soper, F.L. Dynamics of Aedes aegypti distribution and density. Seasonal fluctuations in the Americas. Bull. WHO 1967, 36, 536. [Google Scholar] [PubMed]
- Rozeboom, L.E. Overwintering of Aedes aegypti in Stillwater. Proc. Okla. Acad. Sci. 1938, 19, 81–82. [Google Scholar]
- Lima, A.; Lovin, D.D.; Hickner, P.V.; Severson, D.W. Evidence for an overwintering population of Aedes aegypti in Capitol Hill neighborhood, Washington, DC. Am. J. Trop. Med. Hyg. 2016, 94, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Tsunoda, T.; Cuong, T.C.; Dong, T.D.; Yen, N.T.; Le, N.H.; Phong, T.V.; Minakawa, N. Winter refuge for Aedes aegypti and Ae. albopictus mosquitoes in Hanoi during Winter. PLoS ONE 2014, 9, e95606. [Google Scholar] [CrossRef] [PubMed]
- Lambrechts, L.; Paaijmans, K.P.; Fansiri, T.; Carrington, L.B.; Kramer, L.D.; Thomas, M.B.; Scott, T.W. Impact of daily temperature fluctuations on dengue virus transmission by Aedes aegypti. Proc. Natl. Acad. Sci. USA 2011, 108, 7460–7465. [Google Scholar] [CrossRef] [PubMed]
- Elbers, A.R.W.; Koenraadt, C.J.; Meiswinkel, R. Mosquitoes and Culicoides biting midges: Vector range and the influence of climate change. Rev. Sci. Tech. Off. Int. Epizoot. 2015, 34, 123–137. [Google Scholar] [CrossRef]
- Suwonkerd, W.; Tsuda, Y.; Takagi, M.; Wada, Y. Seasonal occurrence of Aedes aegypti and Ae. albopictus in used tires in 1992–1994, Chiangmai, Thailand. Trop. Med. 1997, 38, 101–105. [Google Scholar]
- Mogi, M. Overwintering strategies of mosquitoes (Diptera: Culicidae) on warmer islands may predict impact of global warming on Kyushu, Japan. J. Med. Entomol. 1996, 33, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Tsunoda, T.; Chaves, L.F.; Nguyen, G.T.T.; Nguyen, Y.T.; Takagi, M. Winter Activity and Diapause of Aedes albopictus (Diptera: Culicidae) in Hanoi, Northern Vietnam. J. Med. Entomol. 2015, 52, 1203–1212. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, Y.; Suwonkerd, W.; Chawprom, S.; Prajakwong, S.; Takagi, M. Different spatial distribution of Aedes aegypti and Aedes albopictus along an urban–rural gradient and the relating environmental factors examined in three villages in northern Thailand. J. Am. Mosq. Control 2006, 22, 222–228. [Google Scholar] [CrossRef]
- Weaver, S.C.; Reisen, W.K. Present and future arboviral threats. Antivir. Res. 2010, 85, 328–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, C.C.; Beebe, N.W. The dengue vector Aedes aegypti: What comes next. Microbes Infect. 2010, 12, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Chaves, L.F.; Morrison, A.C.; Kitron, U.D.; Scott, T.W. Nonlinear impacts of climatic variability on the density-dependent regulation of an insect vector of disease. Glob. Chang. Biol. 2012, 18, 457–468. [Google Scholar] [CrossRef]
- Chaves, L.F.; Scott, T.W.; Morrison, A.C.; Takada, T. Hot temperatures can force delayed mosquito outbreaks via sequential changes in Aedes aegypti demographic parameters in autocorrelated environments. Acta Trop. 2014, 129, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Lewontin, R.; Levins, R. Schmalhausen’s law. Capital. Natl. Soc. 2000, 11, 103–108. [Google Scholar] [CrossRef]
- Chaves, L.F. Globally invasive, withdrawing at home: Aedes albopictus and Aedes japonicus facing the rise of Aedes flavopictus. Int. J. Biometeorol. 2016, 60, 1727–1738. [Google Scholar] [CrossRef] [PubMed]
- Chaves, L.F. Climate change and the biology of insect vectors of human pathogens. Glob. Clim. Chang. Terr. Invertebr. 2017, 126–147. [Google Scholar] [CrossRef]
- Higa, Y.; Thi Yen, N.; Kawada, H.; Hai Son, T.; Thuy Hoa, N.; Takagi, M. Geographic distribution of Aedes aegypti and Aedes albopictus collected from used tires in Vietnam. J. Am. Mosq. Control 2010, 26, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, M.U.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. eLife 2015, 4, e08347. [Google Scholar] [CrossRef] [PubMed]
- Romi, R.; Di Luca, M.; Marjori, G. Current status of Aedes albopictus and Aedes atropalpus in Italy. J. Am. Mosq. Control 1999, 15, 425–427. [Google Scholar]
- Benedict, M.Q.; Levine, R.S.; Hawley, W.A.; Lounibos, L.P. Spread of the tiger: Global risk of invasion by the mosquito Aedes albopictus. Vector-Borne Zoonotic 2007, 7, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Kearney, M.; Porter, W.P.; Williams, C.; Ritchie, S.; Hoffmann, A.A. Integrating biophysical models and evolutionary theory to predict climatic impacts on species’ ranges: The dengue mosquito Aedes aegypti in Australia. Funct. Ecol. 2009, 23, 528–538. [Google Scholar] [CrossRef]
- Alto, B.W.; Juliano, S.A. Precipitation and temperature effects on populations of Aedes albopictus (Diptera: Culicidae): Implications for range expansion. J. Med. Entomol. 2001, 38, 646–656. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.M.; Macoris, M.D.L.D.G.; Galvani, K.C.; Andrighetti, M.T.M.; Wanderley, D.M.V. Assessing the effects of temperature on the population of Aedes aegypti, the vector of dengue. Epidemiol. Infect. 2009, 137, 1188–1202. [Google Scholar] [CrossRef] [PubMed]
- O’meara, G.F.; Evans, L.F., Jr.; Gettman, A.D.; Cuda, J.P. Spread of Aedes albopictus and decline of Ae. aegypti (Diptera: Culicidae) in Florida. J. Med. Entomol. 1995, 32, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Lounibos, L.P.; Suárez, S.; Menéndez, Z.; Nishimura, N.; Escher, R.L.; OConnell, S.M.; Rey, J.R. Does temperature affect the outcome of larval competition between Aedes aegypti and Aedes albopictus? J. Vector Ecol. 2002, 27, 86–95. [Google Scholar] [PubMed]
- Kobayashi, M.; Nihei, N.; Kurihara, T. Analysis of northern distribution of Aedes albopictus (Diptera: Culicidae) in Japan by geographical information system. J. Med. Entomol. 2002, 39, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Mogi, M.; Tuno, N. Impact of climate change on the distribution of Aedes albopictus (Diptera: Culicidae) in northern Japan: Retrospective analyses. J. Med. Entomol. 2014, 51, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Epstein, P.R.; Diaz, H.F.; Elias, S.; Grabherr, G.; Graham, N.E.; Martens, W.J.; MosIey-Thompson, E.; Susskind, J. Biological and physical signs of climate change: Focus on mosquito-borne diseases. Bull. Am. Meteorol. Soc. 1998, 79, 409–417. [Google Scholar] [CrossRef]
- Ostfeld, R.S. Climate change and the distribution and intensity of infectious diseases. Ecology 2009, 90, 903–905. [Google Scholar] [CrossRef] [PubMed]
- Patz, J.A.; Martens, W.J.; Focks, D.A.; Jetten, T.H. Dengue fever epidemic potential as projected by general circulation models of global climate change. Environ. Health Perspect. 1998, 106, 147. [Google Scholar] [CrossRef] [PubMed]
- Sutherst, R.W. Implications of global change and climate variability for vector-borne diseases: Generic approaches to impact assessments. Int. J. Parasitol. 1998, 28, 935–945. [Google Scholar] [CrossRef]
- Rochlin, I.; Ninivaggi, D.V.; Hutchinson, M.L.; Farajollahi, A. Climate change and range expansion of the Asian tiger mosquito (Aedes albopictus) in Northeastern USA: Implications for public health practitioners. PLoS ONE 2013, 8, e60874. [Google Scholar] [CrossRef] [PubMed]
- Kramer, L.D.; Ebel, G.D. Dynamics of flavivirus infection in mosquitoes. Adv. Virus Res. 2003, 60, 187–232. [Google Scholar] [PubMed]
- Gratz, N.G. Critical review of the vector status of Aedes albopictus. Med. Vet. Entomol. 2004, 18, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Vega-Rúa, A.; Zouache, K.; Girod, R.; Failloux, A.B.; Lourenço-de-Oliveira, R. High vector competence of Aedes aegypti and Aedes albopictus from ten American countries as a crucial factor of the spread of Chikungunya. J. Virol. 2014, JVI-00370. [Google Scholar] [CrossRef] [PubMed]
- Brady, O.J.; Golding, N.; Pigott, D.M.; Kraemer, M.U.; Messina, J.P.; Reiner, R.C., Jr.; Scott, T.W.; Smith, D.L.; Gething, P.W.; Hay, S.I. Global temperature constraints on Aedes aegypti and Aedes albopictus persistence and competence for dengue virus transmission. Parasite Vector 2014, 7, 338. [Google Scholar] [CrossRef] [PubMed]
- Patz, J.A.; Githeko, A.K.; McCarty, J.P.; Hussein, S.; Confalonieri, U.; De Wet, N. Climate change and infectious diseases. Clim. Chang. Hum. Health Risks Responses 2003, 6, 103–137. [Google Scholar]
- Lafferty, K.D. The ecology of climate change and infectious diseases. Ecology 2009, 90, 888–900. [Google Scholar] [CrossRef] [PubMed]
- McMichael, A.J.; Woodruff, R.E. Climate change and infectious diseases. Soc. Ecol. Infect. Dis. 2008, 378–407. [Google Scholar] [CrossRef]
- World Health Organization. Dengue and Severe Dengue. Available online: http://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue (accessed on 13 September 2018).
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O. The global distribution and burden of dengue. Nature 2013, 496, 504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brady, O.J.; Gething, P.W.; Bhatt, S.; Messina, J.P.; Brownstein, J.S.; Hoen, A.G.; Moyes, C.L.; Farlow, A.W.; Scott, T.W.; Hay, S.I. Refining the global spatial limits of dengue virus transmission by evidence-based consensus. PLoS NTDs 2012, 6, e1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watts, D.M.; Burke, D.S.; Harrison, B.A.; Whitmire, R.E.; Nisalak, A. Effect of temperature on the vector efficiency of Aedes aegypti for dengue 2 virus. Am. J. Trop. Med. Hyg. 1987, 36, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Rohani, A.; Wong, Y.C.; Zamre, I.; Lee, H.L.; Zurainee, M.N. The effect of extrinsic incubation temperature on development of dengue serotype 2 and 4 viruses in Aedes aegypti (L.). SE Asian J. Trop. Med. 2009, 40, 942. [Google Scholar]
- Alto, B.W.; Bettinardo, D. Temperature and dengue virus infection in mosquitoes: Independent effects of the immature and adult stages. Am. J. Trop. Med. Hyg. 2013, 88, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Whitehorn, J.; Kien, D.T.; Nguyen, N.M.; Nguyen, H.L.; Kyrylos, P.P.; Carrington, L.B.; Tran, C.N.; Quyen, N.T.; Thi, L.V.; Le Thi, D.; et al. Comparative susceptibility of Aedes albopictus and Aedes aegypti to dengue virus infection after feeding on blood of viremic humans: Implications for public health. J. Infect. Dis. 2015, 212, 1182–1190. [Google Scholar] [CrossRef] [PubMed]
- Thu, H.M.; Aye, K.M.; Thein, S. The effect of temperature and humidity on dengue virus propagation in Aedes aegypti mosquitoes. Southeast Asian J. Trop. Med. Public Health 1998, 29, 280–284. [Google Scholar] [PubMed]
- World Health Organization. Chikungunya. Available online: http://www.who.int/news-room/fact-sheets/detail/chikungunya (accessed on 12 April 2017).
- Renault, P.; Solet, J.L.; Sissoko, D.; Balleydier, E.; Larrieu, S.; Filleul, L.; Lassalle, C.; Thiria, J.; Rachou, E.; de Valk, H.; et al. A major epidemic of chikungunya virus infection on Reunion Island, France, 2005–2006. Am. J. Trop. Med. Hyg. 2007, 77, 727–731. [Google Scholar] [CrossRef] [PubMed]
- Yoon, I.K.; Alera, M.T.; Lago, C.B.; Tac-An, I.A.; Villa, D.; Fernandez, S.; Thaisomboonsuk, B.; Klungthong, C.; Levy, J.W.; Velasco, J.M.; et al. High rate of subclinical chikungunya virus infection and association of neutralizing antibody with protection in a prospective cohort in the Philippines. PLoS NTDs 2015, 9, e0003764. [Google Scholar] [CrossRef] [PubMed]
- Becker, N. Influence of climate change on mosquito development and mosquito-borne diseases in Europe. Parasitol. Res. 2008, 103, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Paupy, C.; Delatte, H.; Bagny, L.; Corbel, V.; Fontenille, D. Aedes albopictus, an arbovirus vector: From the darkness to the light. Microbes Infect. 2009, 11, 1177–1185. [Google Scholar] [CrossRef] [PubMed]
- Zouache, K.; Fontaine, A.; Vega-Rua, A.; Mousson, L.; Thiberge, J.M.; Lourenco-De-Oliveira, R.; Caro, V.; Lambrechts, L.; Failloux, A.B. Three-way interactions between mosquito population, viral strain and temperature underlying chikungunya virus transmission potential. Proc. R. Soc. Lond. B Biol. Sci. 2014, 281, 20141078. [Google Scholar] [CrossRef] [PubMed]
- Li, C.X.; Guo, X.X.; Deng, Y.Q.; Xing, D.; Sun, A.J.; Liu, Q.M.; Wu, Q.; Zhang, Y.M.; Zhang, H.D.; Cao, W.C.; et al. Vector competence and transovarial transmission of two Aedes aegypti strains to Zika virus. Emerg. Microbes Infec. 2017, 6, e23. [Google Scholar] [CrossRef] [PubMed]
- Niyas, K.P.; Abraham, R.; Unnikrishnan, R.N.; Mathew, T.; Nair, S.; Manakkadan, A.; Issac, A.; Sreekumar, E. Molecular characterization of Chikungunya virus isolates from clinical samples and adult Aedes albopictus mosquitoes emerged from larvae from Kerala, South India. Virol. J. 2010, 7, 189. [Google Scholar] [CrossRef] [PubMed]
- Mavale, M.; Parashar, D.; Sudeep, A.; Gokhale, M.; Ghodke, Y.; Geevarghese, G.; Arankalle, V.; Mishra, A.C. Venereal transmission of chikungunya virus by Aedes aegypti mosquitoes (Diptera: Culicidae). Am. J. Trop. Med. Hyg. 2010, 83, 1242–1244. [Google Scholar] [CrossRef] [PubMed]
- Cauchemez, S.; Ledrans, M.; Poletto, C.; Quenel, P.D.; De Valk, H.; Colizza, V.; Boëlle, P.Y. Local and regional spread of chikungunya fever in the Americas. Euro Surveillance: Bulletin Europeen sur les Maladies Transmissibles=Eur. Commun. Dis. Bull. 2014, 19, 20854. [Google Scholar] [CrossRef] [Green Version]
- Kendrick, K.; Stanek, D.; Blackmore, C.; Centers for Disease Control and Prevention (CDC). Notes from the field: Transmission of chikungunya virus in the continental United States—Florida, 2014. MMWR Morb. Mortal. Wkly. Rep. 2014, 63, 1137. [Google Scholar] [PubMed]
- Center for Disease Control and Prevention. Chikungunya Virus Home: Geographic Distribution. Available online: https://www.cdc.gov/chikungunya/geo/index.html (accessed on 29 May 2018).
- Barba-Spaeth, G.; Dejnirattisai, W.; Rouvinski, A.; Vaney, M.C.; Medits, I.; Sharma, A.; Simon-Lorière, E.; Sakuntabhai, A.; Cao-Lormeau, V.M.; Haouz, A.; et al. Structural basis of potent Zika–dengue virus antibody cross-neutralization. Nature 2016, 536, 48. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Zika Virus. Available online: http://www.who.int/news-room/fact-sheets/detail/zika-virus (accessed on 20 July 2018).
- Benelli, G.; Romano, D. Mosquito vectors of Zika virus. Entomol. Gen. 2017, 36, 309–318. [Google Scholar] [CrossRef]
- Hayes, E.B. Zika virus outside Africa. Emerg. Infect. Dis. 2009, 15, 1347. [Google Scholar] [CrossRef] [PubMed]
- Haddow, A.D.; Schuh, A.J.; Yasuda, C.Y.; Kasper, M.R.; Heang, V.; Huy, R.; Guzman, H.; Tesh, R.B.; Weaver, S.C. Genetic characterization of Zika virus strains: Geographic expansion of the Asian lineage. PLoS NTDs 2012, 6, e1477. [Google Scholar] [CrossRef] [PubMed]
- Petersen, L.R.; Jamieson, D.J.; Powers, A.M.; Honein, M.A. Zika virus. New Engl. J. Med. 2016, 374, 1552–1563. [Google Scholar] [CrossRef] [PubMed]
- Haddow, A.D.; Schuh, A.J.; Yasuda, C.Y.; Kasper, M.R.; Heang, V.; Huy, R.; Guzman, H.; Tesh, R.B.; Weaver, S.C. On the seasonal occurrence and abundance of the Zika virus vector mosquito Aedes aegypti in the contiguous United States. PLoS Curr. 2016, 8. [Google Scholar] [CrossRef]
- Mordecai, E.A.; Cohen, J.M.; Evans, M.V.; Gudapati, P.; Johnson, L.R.; Lippi, C.A.; Miazgowicz, K.; Murdock, C.C.; Rohr, J.R.; Ryan, S.J.; et al. Detecting the impact of temperature on transmission of Zika, dengue, and chikungunya using mechanistic models. PLoS NTDs 2017, 11, e0005568. [Google Scholar] [CrossRef] [PubMed]
- Azar, S.R.; Roundy, C.M.; Rossi, S.L.; Huang, J.H.; Leal, G.; Yun, R.; Fernandez-Salas, I.; Vitek, C.J.; Paploski, I.A.; Stark, P.M.; et al. Differential vector competency of Aedes albopictus populations from the Americas for Zika virus. Am. J. Trop. Med. Hyg. 2017, 97, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Gendernalik, A.; Weger-Lucarelli, J.; Luna, S.M.G.; Fauver, J.R.; Rückert, C.; Murrieta, R.A.; Burgren, N.; Samaras, D.; Nguyen, C.; Kading, R.C.; Ebel, G.D. American Aedes vexans mosquitoes are competent vectors of Zika virus. Am. J. Trop. Med. Hyg. 2017, 96, 1338–1340. [Google Scholar] [CrossRef] [PubMed]
- Ayres, C.F. Identification of Zika virus vectors and implications for control. Lancet Infect. Dis. 2016, 16, 278–279. [Google Scholar] [CrossRef]
- Gardner, L.M.; Chen, N.; Sarkar, S. Global risk of Zika virus depends critically on vector status of Aedes albopictus. Lancet Infect. Dis. 2016, 16, 522–523. [Google Scholar] [CrossRef]
- World Health Organization. West Nile Virus. Available online: http://www.who.int/news-room/fact-sheets/detail/west-nile-virus (accessed on 3 October 2017).
- Klenk, K.; Snow, J.; Morgan, K.; Bowen, R.; Stephens, M.; Foster, F.; Gordy, P.; Beckett, S.; Komar, N.; Gubler, D.; et al. Alligators as West Nile virus amplifiers. Emerg. Infect. Dis. 2004, 10, 2150. [Google Scholar] [CrossRef] [PubMed]
- Campbell, G.L.; Marfin, A.A.; Lanciotti, R.S.; Gubler, D.J. West Nile virus. Lancet Infect. Dis. 2002, 2, 519–529. [Google Scholar] [CrossRef]
- Turell, M.J.; O’Guinn, M.L.; Dohm, D.J.; Jones, J.W. Vector competence of North American mosquitoes (diptera: Culicidae) for West Nile virus. J. Med. Entomol. 2001, 38, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Turell, M.J.; Dohm, D.J.; Sardelis, M.R.; O’guinn, M.L.; Andreadis, T.G.; Blow, J.A. An update on the potential of North American mosquitoes (Diptera: Culicidae) to transmit West Nile virus. J. Med. Entomol. 2005, 42, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Richards, S.L.; Mores, C.N.; Lord, C.C.; Tabachnick, W.J. Impact of extrinsic incubation temperature and virus exposure on vector competence of Culex pipiens quinquefasciatus Say (Diptera: Culicidae) for West Nile virus. Vector-Borne Zoonotic 2007, 7, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Dohm, D.J.; O’Guinn, M.L.; Turell, M.J. Effect of environmental temperature on the ability of Culex pipiens (Diptera: Culicidae) to transmit West Nile virus. J. Med. Entomol. 2002, 39, 221–225. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Yellow Fever Virus. Available online: http://www.who.int/news-room/fact-sheets/detail/yellow-fever (accessed on 1 May 2018).
- Rogers, D.J.; Wilson, A.J.; Hay, S.I.; Graham, A.J. The global distribution of yellow fever and dengue. Adv. Parasit. 2006, 62, 181–220. [Google Scholar]
- Aitken, T.H.; Tesh, R.B.; Beaty, B.J.; Rosen, L. Transovarial transmission of yellow fever virus by mosquitoes (Aedes aegypti). Am. J. Trop. Med. Hyg. 1979, 28, 119–121. [Google Scholar] [CrossRef] [PubMed]
- Center for Disease Control and Prevention. Transmission of Yellow Fever Virus. Available online: https://www.cdc.gov/yellowfever/transmission/index.html (accessed on 13 August 2015).
- Gubler, D.J. The changing epidemiology of yellow fever and dengue, 1900 to 2003: Full circle? Comp. Immunol. Microb. 2004, 27, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Davis, N.C. The Effect of Various Temperatures in modifying the Extrinsic Incubation Period of the Yellow Fever Virus in Aedes aegypti. Am. J. Hyg. 1932, 16, 163–176. [Google Scholar] [CrossRef]
- Moore, C.G.; Mitchell, C.J. Aedes albopictus in the United States: Ten-year presence and public health implications. Emerg. Infect. Dis. 1997, 3, 329. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.J.; Niebylski, M.L.; Smith, G.C.; Karabatsos, N.; Martin, D.; Mutebi, J.P.; Craig, G.B.; Mahler, M.J. Isolation of eastern equine encephalitis virus from Aedes albopictus in Florida. Science 1992, 257, 526–527. [Google Scholar] [CrossRef] [PubMed]
- Komar, N.; Dohm, D.J.; Turell, M.J.; Spielman, A. Eastern equine encephalitis virus in birds: Relative competence of European starlings (Sturnus vulgaris). Am. J. Trop. Med. Hyg. 1999, 60, 387–391. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, R.; Sudia, W. The Effects of Temperature upon the Extrinsic Incubation of Eastern Equine Encephalitis in Mosquitoes. Am. J. Hyg. 1955, 62, 295–305. [Google Scholar] [PubMed]
- Lednicky, J.; De Rochars, V.M.B.; Loeb, J.; Telisma, T.; Chavannes, S.; Anilis, G.; Cella, E. Ciccozzi, M.; Okech, B.; Salemi, M.; et al. Mayaro virus in child with acute febrile illness, Haiti, 2015. Emerg. Infect. Dis. 2016, 22, 2000. [Google Scholar] [CrossRef] [PubMed]
- Da Costa Carvalho, M.D.G.; Fournier, M.V. Effect of heat shock on gene expression of Aedes albopictus cells infected with Mayaro virus. Res. Virol. 1991, 142, 25–31. [Google Scholar] [CrossRef]
- Hotez, P.J.; Murray, K.O. Dengue, West Nile virus, chikungunya, Zika—And now Mayaro? PLoS Negl. Trop. Dis. 2017, 11, e005462. [Google Scholar] [CrossRef] [PubMed]
- Kartman, L. Factors influencing infection of the mosquito with Dirofilaria immitis (Leidy, 1856). Exp. Parasitol. 1953, 2, 27–78. [Google Scholar] [CrossRef]
- McGreevy, P.B.; Kolstrup, N.; Tao, J.; McGreevy, M.M.; de C. Marshall, T.F. Ingestion and development of Wuchereria bancrofti in Culex quinquefasciatus, Anopheles gambiae and Aedes aegypti after feeding on humans with varying densities of microfilariae in Tanzania. Trans. R. Soc. Trop. Med. Hyg. 1982, 76, 288–296. [Google Scholar] [CrossRef]
- Simón, F.; López-Belmonte, J.; Marcos-Atxutegi, C.; Morchón, R.; Martín-Pacho, J.R. What is happening outside North America regarding human dirofilariasis? Vet. Parasitol. 2005, 133, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Ledesma, N.; Harrington, L. Mosquito vectors of dog heartworm in the United States: Vector status and factors influencing transmission efficiency. Top. Companion Anim. Med. 2011, 26, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Ledesma, N.; Harrington, L. Fine-scale temperature fluctuation and modulation of Dirofilaria immitis larval development in Aedes aegypti. Vet. Parasitol. 2015, 209, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Mas-Coma, S.; Valero, M.A.; Bargues, M.D. Effects of climate change on animal and zoonotic helminthiases. Rev. Sci. Tech. 2008, 27, 443–457. [Google Scholar] [CrossRef] [PubMed]
- Beck-Johnson, L.M.; Nelson, W.A.; Paaijmans, K.P.; Read, A.F.; Thomas, M.B.; Bjørnstad, O.N. The effect of temperature on Anopheles mosquito population dynamics and the potential for malaria transmission. PLoS ONE 2013, 8, e79276. [Google Scholar] [CrossRef] [PubMed]
- Paaijmans, K.P.; Blanford, S.; Bell, A.S.; Blanford, J.I.; Read, A.F.; Thomas, M.B. Influence of climate on malaria transmission depends on daily temperature variation. Proc. Natl. Acad. Sci. USA 2010, 107, 15135–15139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanderberg, J.P.; Yoeli, M. Effects of temperature on sporogonic development of Plasmodium berghei. J. Parasitol. 1966, 52, 559–564. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The temperature of the environment (Ta) affects the mosquito development (blue), its activity including host-seeking and blood-meal intake (red), as well as pathogen development and transmission (purple). Consequently, Ta affects species geographic repartition, spatial distribution, and population dynamics (green). The dashed square represents the cycles related to mosquito biology.
Figure 1.
The temperature of the environment (Ta) affects the mosquito development (blue), its activity including host-seeking and blood-meal intake (red), as well as pathogen development and transmission (purple). Consequently, Ta affects species geographic repartition, spatial distribution, and population dynamics (green). The dashed square represents the cycles related to mosquito biology.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Reinhold, J.M.; Lazzari, C.R.; Lahondère, C. Effects of the Environmental Temperature on Aedes aegypti and Aedes albopictus Mosquitoes: A Review. Insects 2018, 9, 158. https://0-doi-org.brum.beds.ac.uk/10.3390/insects9040158
AMA Style
Reinhold JM, Lazzari CR, Lahondère C. Effects of the Environmental Temperature on Aedes aegypti and Aedes albopictus Mosquitoes: A Review. Insects. 2018; 9(4):158. https://0-doi-org.brum.beds.ac.uk/10.3390/insects9040158
Chicago/Turabian StyleReinhold, Joanna M., Claudio R. Lazzari, and Chloé Lahondère. 2018. "Effects of the Environmental Temperature on Aedes aegypti and Aedes albopictus Mosquitoes: A Review" Insects 9, no. 4: 158. https://0-doi-org.brum.beds.ac.uk/10.3390/insects9040158
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.