Sequential Infection of Aedes aegypti Mosquitoes with Chikungunya Virus and Zika Virus Enhances Early Zika Virus Transmission

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mosquitoes and Viruses
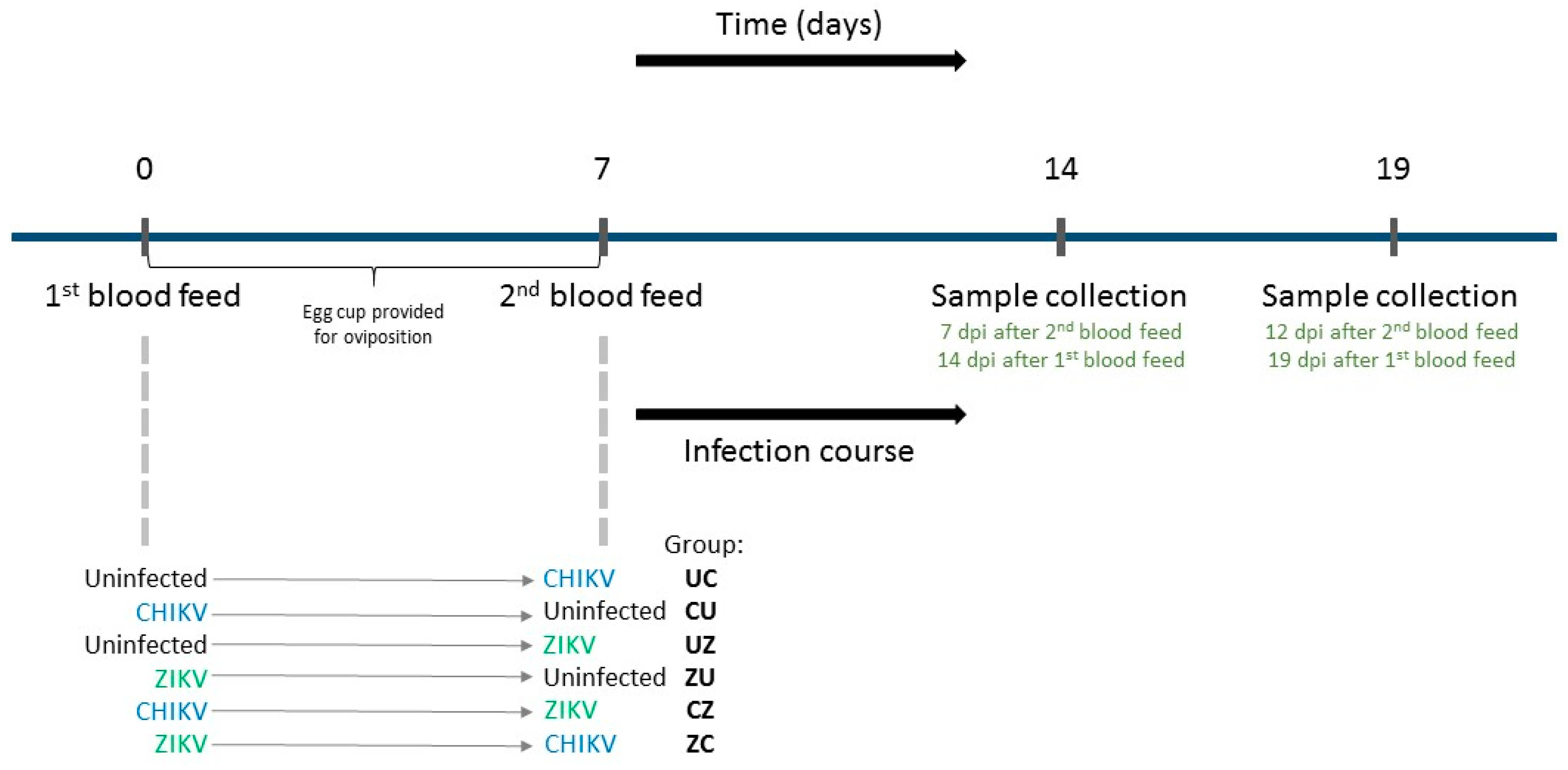
2.2. Sequential Infections with CHIKV and ZIKV, and Sample Collection
2.3. Viral RNA Extraction and Quantitative Real-Time Reverse Transcriptase PCR (qRT-PCR)
2.4. Statistical Analysis
3. Results
3.1. Infection and Dissemination Rates
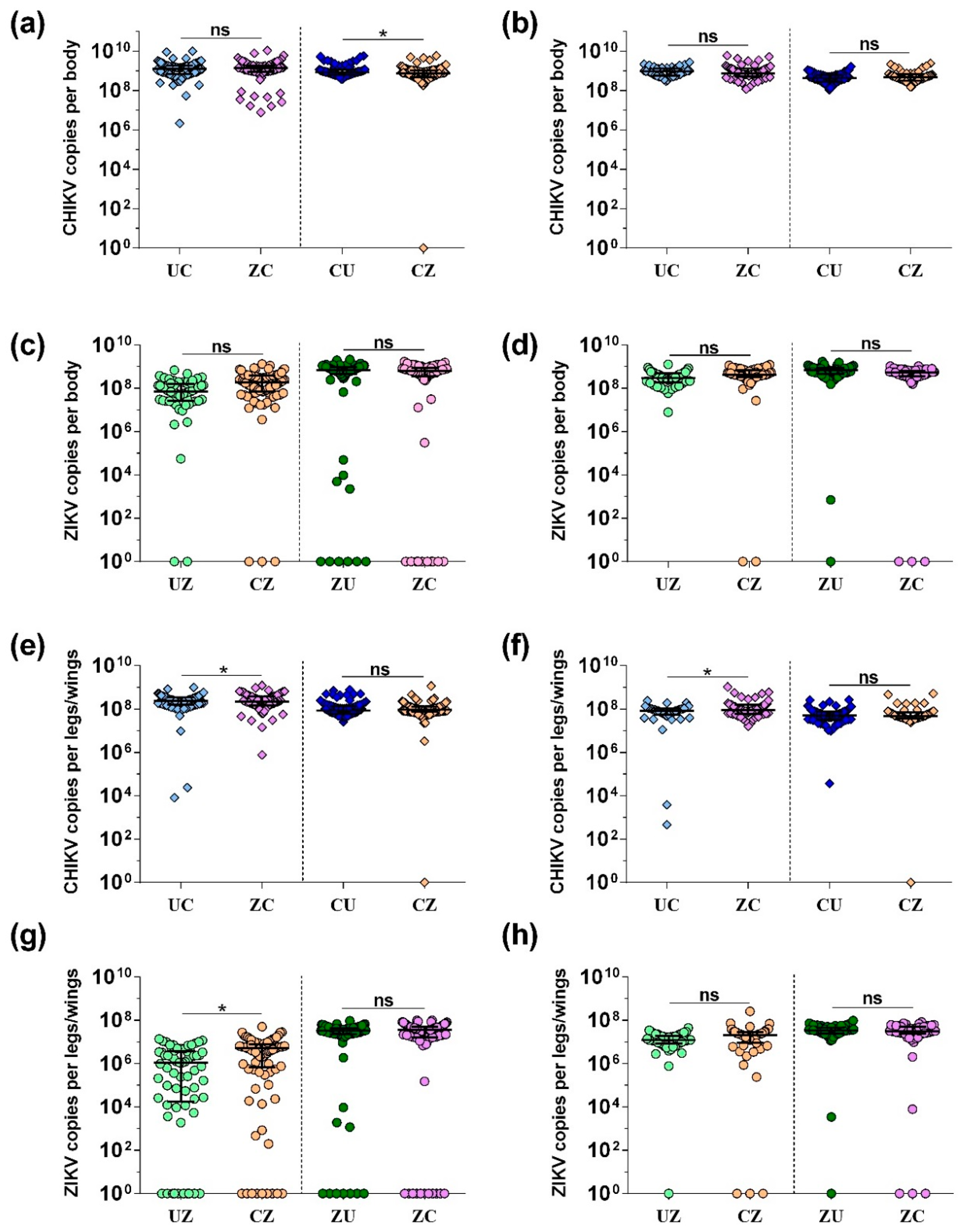
3.2. CHIKV and ZIKV Load in Bodies and Legs/Wings
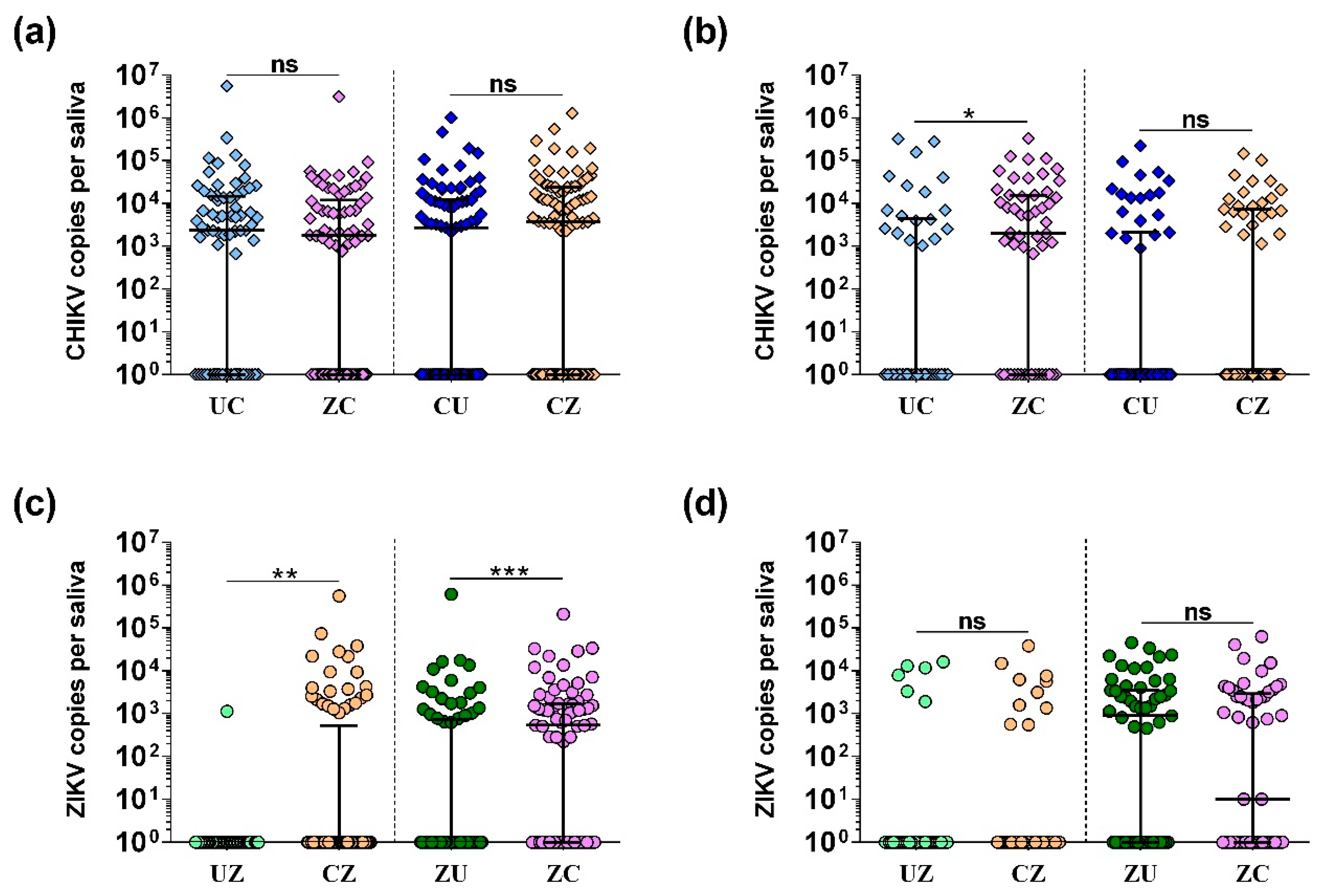
3.3. Transmission Rates and Viral Load in Saliva
3.4. Ae. aegypti Can Co-Transmit CHIKV and ZIKV after Sequential Exposure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Morrison, T.E. Reemergence of chikungunya virus. J. Virol. 2014, 88, 11644–11647. [Google Scholar] [CrossRef] [PubMed]
- Grubaugh, N.D.; Faria, N.R.; Andersen, K.G.; Pybus, O.G. Genomic Insights into Zika Virus Emergence and Spread. Cell 2018, 172, 1160–1162. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Morales, A.J.; Villamil-Gomez, W.E.; Franco-Paredes, C. The arboviral burden of disease caused by co-circulation and co-infection of dengue, chikungunya and Zika in the Americas. Travel Med. Infect. Dis. 2016, 14, 177–179. [Google Scholar] [CrossRef] [PubMed]
- Doughty, C.T.; Yawetz, S.; Lyons, J. Emerging Causes of Arbovirus Encephalitis in North America: Powassan, Chikungunya, and Zika Viruses. Curr. Neurol. Neurosci. Rep. 2017, 17, 12. [Google Scholar] [CrossRef] [PubMed]
- Pierson, T.C.; Diamond, M.S. The emergence of Zika virus and its new clinical syndromes. Nature 2018, 560, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Edman, J.D. Fitness advantages in multiple blood-feeding: The Aedes aegypti example. In Ecological Aspects for Application of Genetically Modified Mosquitoes; Takken, W., Scott, T.W., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 63–74. [Google Scholar]
- Farjana, T.; Tuno, N. Multiple blood feeding and host-seeking behavior in Aedes aegypti and Aedes albopictus (Diptera: Culicidae). J. Med. Entomol. 2013, 50, 838–846. [Google Scholar] [CrossRef] [PubMed]
- White, S.K.; Mavian, C.; Elbadry, M.A.; Beau De Rochars, V.M.; Paisie, T.; Telisma, T.; Salemi, M.; Lednicky, J.A.; Morris, J.G. Detection and phylogenetic characterization of arbovirus dual-infections among persons during a chikungunya fever outbreak, Haiti 2014. PLoS Negl. Trop. Dis. 2018, 12, e0006505. [Google Scholar] [CrossRef] [PubMed]
- Sardi, S.I.; Somasekar, S.; Naccache, S.N.; Bandeira, A.C.; Tauro, L.B.; Campos, G. Coinfections of Zika and Chikungunya Viruses in Bahia, Brazil, Identified by Metagenomic Next-Generation Sequencing. J. Clin. Microbiol. 2016, 54, 2348–2353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waggoner, J.J.; Gresh, L.; Vargas, M.J.; Ballesteros, G.; Tellez, Y.; Soda, K.J.; Sahoo, M.K.; Nuñez, A.; Balmaseda, A.; Harris, E.; et al. Viremia and Clinical Presentation in Nicaraguan Patients Infected with Zika Virus, Chikungunya Virus, and Dengue Virus. Clin. Infect. Dis. 2016, 63, 1584–1590. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-Hernandez, M.Y.; Ruiz-Saenz, J.; Villamizar, L.J.; Gomez-Rangel, S.Y.; Martinez-Gutierrez, M. Co-circulation and simultaneous co-infection of dengue, chikungunya, and zika viruses in patients with febrile syndrome at the Colombian-Venezuelan border. BMC Infect. Dis. 2018, 18, 61. [Google Scholar] [CrossRef] [PubMed]
- Cherabuddi, K.; Iovine, N.M.; Shah, K.; White, S.K.; Paisie, T.; Salemi, M.; Morris, J.G.; Lednicky, J.A. Zika and Chikungunya virus co-infection in a traveller returning from Colombia, 2016, virus isolation and genetic analysis. JMM Case Rep. 2016, 3, e005072. [Google Scholar] [CrossRef] [PubMed]
- Norman, F.F.; Chamorro, S.; Vazquez, A.; Sanchez-Seco, M.P.; Perez-Molina, J.A.; Monge-Maillo, B.; Vivancos, M.J.; Rodríguez-Dominguez, M.; Galán, J.C.; de Ory, F.; et al. Sequential Chikungunya and Zika Virus Infections in a Traveler from Honduras. Am. J. Trop. Med. Hyg. 2016, 95, 1166–1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zambrano, H.; Waggoner, J.J.; Almeida, C.; Rivera, L.; Benjamin, J.Q.; Pinsky, B.A. Zika Virus and Chikungunya Virus CoInfections: A Series of Three Cases from a Single Center in Ecuador. Am. J. Trop. Med. Hyg. 2016, 95, 894–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acevedo, N.; Waggoner, J.; Rodriguez, M.; Rivera, L.; Landivar, J.; Pinsky, B.; Zambrano, H. Zika Virus, Chikungunya Virus, and Dengue Virus in Cerebrospinal Fluid from Adults with Neurological Manifestations, Guayaquil, Ecuador. Front. Microbiol. 2017, 8, 42. [Google Scholar] [CrossRef] [PubMed]
- Brito, C.A.A.; Azevedo, F.; Cordeiro, M.T.; Marques, E.T.A., Jr. Franca RFO Central and peripheral nervous system involvement caused by Zika and chikungunya coinfection. PLoS Negl. Trop. Dis. 2017, 11, e0005583. [Google Scholar] [CrossRef] [PubMed]
- Caron, M.; Paupy, C.; Grard, G.; Becquart, P.; Mombo, I.; Nso, B.B.; Kassa, F.; Nkoghe, D.; Leroy, E.M. Recent introduction and rapid dissemination of Chikungunya virus and Dengue virus serotype 2 associated with human and mosquito coinfections in Gabon, central Africa. Clin. Infect. Dis. 2012, 55, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Göertz, G.P.; Vogels, C.B.F.; Geertsema, C.; Koenraadt, C.J.M.; Pijlman, G.P. Mosquito co-infection with Zika and chikungunya virus allows simultaneous transmission without affecting vector competence of Aedes aegypti. PLoS Negl. Trop. Dis. 2017, 11, e0005654. [Google Scholar] [CrossRef] [PubMed]
- Rückert, C.; Weger-Lucarelli, J.; Garcia-Luna, S.M.; Young, M.C.; Byas, A.D.; Murrieta, R.A.; Fauver, J.R.; Ebel, G.D. Impact of simultaneous exposure to arboviruses on infection and transmission by Aedes aegypti mosquitoes. Nat. Commun. 2017, 8, 15412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samuel, G.H.; Adelman, Z.N.; Myles, K.M. Antiviral Immunity and Virus-Mediated Antagonism in Disease Vector Mosquitoes. Trends Microbiol. 2018, 26, 447–461. [Google Scholar] [CrossRef] [PubMed]
- DaPalma, T.; Doonan, B.P.; Trager, N.M.; Kasman, L.M. A systematic approach to virus-virus interactions. Virus Res. 2010, 149, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bara, J.J.; Muturi, E.J. Effect of mixed infections of Sindbis and La Crosse viruses on replication of each virus in vitro. Acta Trop. 2014, 130, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Potiwat, R.; Komalamisra, N.; Thavara, U.; Tawatsin, A.; Siriyasatien, P. Competitive suppression between chikungunya and dengue virus in Aedes albopictus c6/36 cell line. Southeast Asian J. Trop. Med. Public Health 2011, 42, 1388–1394. [Google Scholar] [PubMed]
- Pepin, K.M.; Lambeth, K.; Hanley, K.A. Asymmetric competitive suppression between strains of dengue virus. BMC Microbiol. 2008, 8, 28. [Google Scholar] [CrossRef] [PubMed]
- Abrao, E.P.; da Fonseca, B.A. Infection of Mosquito Cells (C6/36) by Dengue-2 Virus Interferes with Subsequent Infection by Yellow Fever Virus. Vector Borne Zoonotic Dis. 2016, 16, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Nuckols, J.T.; Huang, Y.J.; Higgs, S.; Miller, A.L.; Pyles, R.B.; Spratt, H.M.; Horne, K.M.; Vanlandingham, D.L. Evaluation of Simultaneous Transmission of Chikungunya Virus and Dengue Virus Type 2 in Infected Aedes aegypti and Aedes albopictus (Diptera: Culicidae). J. Med. Entomol. 2015, 52, 447–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Coupanec, A.; Tchankouo-Nguetcheu, S.; Roux, P.; Khun, H.; Huerre, M.; Morales-Vargas, R.; Enguehard, M.; Lavillette, D.; Missé, D.; Choumet, V. Co-Infection of Mosquitoes with Chikungunya and Dengue Viruses Reveals Modulation of the Replication of Both Viruses in Midguts and Salivary Glands of Aedes aegypti Mosquitoes. Int. J. Mol. Sci. 2017, 18, 1708. [Google Scholar] [CrossRef] [PubMed]
- Mourya, D.T.; Gokhale, M.D.; Majumdar, T.D.; Yadav, P.D.; Kumar, V.; Mavale, M.S. Experimental Zika virus infection in Aedes aegypti: Susceptibility, transmission & co-infection with dengue & chikungunya viruses. Indian J. Med. Res. 2018, 147, 88–96. [Google Scholar] [PubMed]
- Vera-Maloof, F.Z.; Saavedra-Rodriguez, K.; Elizondo-Quiroga, A.E.; Lozano-Fuentes, S.; Black Iv, W.C. Coevolution of the Ile1,016 and Cys1,534 Mutations in the Voltage Gated Sodium Channel Gene of Aedes aegypti in Mexico. PLoS Negl. Trop. Dis. 2015, 9, e0004263. [Google Scholar] [CrossRef] [PubMed]
- Vazeille, M.; Mousson, L.; Martin, E.; Failloux, A.B. Orally co-Infected Aedes albopictus from La Reunion Island, Indian Ocean, can deliver both dengue and chikungunya infectious viral particles in their saliva. PLoS Negl. Trop. Dis. 2010, 4, e706. [Google Scholar] [CrossRef] [PubMed]
- Ruckert, C.; Ebel, G.D. How Do Virus-Mosquito Interactions Lead to Viral Emergence? Trends Parasitol. 2018, 34, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Sirisena, P.; Kumar, A.; Sunil, S. Evaluation of Aedes aegypti (Diptera: Culicidae) Life Table Attributes Upon Chikungunya Virus Replication Reveals Impact on Egg-Laying Pathways. J. Med. Entomol. 2018, 55, 1580–1587. [Google Scholar] [CrossRef] [PubMed]
- Kantor, A.M.; Dong, S.; Held, N.L.; Ishimwe, E.; Passarelli, A.L.; Clem, R.J.; Franz, A.W. Identification and initial characterization of matrix metalloproteinases in the yellow fever mosquito, Aedes aegypti. Insect Mol. Biol. 2017, 26, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Balaraman, V.; Kantor, A.M.; Lin, J.; Grant, D.G.; Held, N.L.; Franz, A.W. Chikungunya virus dissemination from the midgut of Aedes aegypti is associated with temporal basal lamina degradation during bloodmeal digestion. PLoS Negl. Trop. Dis. 2017, 11, e0005976. [Google Scholar] [CrossRef] [PubMed]
- Blair, C.D.; Olson, K.E. The role of RNA interference (RNAi) in arbovirus-vector interactions. Viruses 2015, 7, 820–843. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.L.; Dodd, B.J.; Brackney, D.E.; Wilusz, C.J.; Ebel, G.D.; Wilusz, J. Flavivirus sfRNA suppresses antiviral RNA interference in cultured cells and mosquitoes and directly interacts with the RNAi machinery. Virology 2015, 485, 322–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McFarlane, M.; Arias-Goeta, C.; Martin, E.; O’Hara, Z.; Lulla, A.; Mousson, L.; Rainey, S.M.; Misbah, S.; Schnettler, E.; Donald, C.L.; et al. Characterization of Aedes aegypti innate-immune pathways that limit Chikungunya virus replication. PLoS Negl. Trop. Dis. 2014, 8, e2994. [Google Scholar] [CrossRef] [PubMed]
- Magalhaes, T.; Braga, C.; Cordeiro, M.T.; Oliveira, A.L.S.; Castanha, P.M.S.; Maciel, A.P.; Amancio, N.M.; Gouveia, P.N.; Peixoto-da-Silva, V.J.; Peixoto, T.F.; et al. Zika virus displacement by a chikungunya outbreak in Recife, Brazil. PLoS Negl. Trop. Dis. 2017, 11, e0006055. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| CHIKV | 7 dpi | 12 dpi | ||||
| Infection | Dissemination | Transmission | Infection | Dissemination | Transmission | |
| UC | 72/72 (100%) | 72/72 (100%) | 46/72 (63.9%) | 42/42 (100%) | 42/42 (100%) | 18/42 (42.9%) |
| CU | 83/83 (100%) | 82/83 (98.8%) | 43/83 (51.8%) | 59/59 (100%) | 59/59 (100%) | 19/59 (32.2%) |
| UZ | - | - | - | - | - | - |
| ZU | - | - | - | - | - | - |
| CZ | 92/93 (98.9%) | 91/93 (97.8%) | 55/93 (59.1%) | 52/52 (100%) | 51/52 (98.1%) | 23/52 (44.2%) |
| ZC | 74/74 (100%) | 70/74 (94.6%) | 45/74 (60.8%) | 53/53 (100%) | 53/53 (100%) | 36/53 (67.9%) |
| ZIKV | 7 dpi | 12 dpi | ||||
| Infection | Dissemination | Transmission | Infection | Dissemination | Transmission | |
| UC | - | - | - | - | - | - |
| CU | - | - | - | - | - | - |
| UZ | 63/65 (96.9%) | 55/65 (84.6%) | 1/65 (1.54%) | 44/44 (100%) | 43/44 (97.7%) | 9/44 (20.5%) |
| ZU | 74/80 (92.5%) | 73/80 (91.3%) | 22/80 (27.5%) | 54/55 (91.3%) | 54/55 (91.3%) | 32/55 (58.2%) |
| CZ | 90/93 (96.8%) | 83/93 (89.2%) | 23/93 (24.7%) B | 50/52 (96.2%) | 49/52 (94.2%) | 10/52 (19.2%) |
| ZC | 63/74 (86.5%) | 63/74 (85.1%) | 44/74 (59.5%) C | 50/53 (94.3%) | 50/53 (94.3%) | 27/53 (50.9%) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magalhaes, T.; Robison, A.; Young, M.C.; Black, W.C.; Foy, B.D.; Ebel, G.D.; Rückert, C. Sequential Infection of Aedes aegypti Mosquitoes with Chikungunya Virus and Zika Virus Enhances Early Zika Virus Transmission. Insects 2018, 9, 177. https://0-doi-org.brum.beds.ac.uk/10.3390/insects9040177
Magalhaes T, Robison A, Young MC, Black WC, Foy BD, Ebel GD, Rückert C. Sequential Infection of Aedes aegypti Mosquitoes with Chikungunya Virus and Zika Virus Enhances Early Zika Virus Transmission. Insects. 2018; 9(4):177. https://0-doi-org.brum.beds.ac.uk/10.3390/insects9040177
Chicago/Turabian StyleMagalhaes, Tereza, Alexis Robison, Michael C. Young, William C. Black, Brian D. Foy, Gregory D. Ebel, and Claudia Rückert. 2018. "Sequential Infection of Aedes aegypti Mosquitoes with Chikungunya Virus and Zika Virus Enhances Early Zika Virus Transmission" Insects 9, no. 4: 177. https://0-doi-org.brum.beds.ac.uk/10.3390/insects9040177