
The Histone H3 K4me3, K27me3, and K27ac Genome-Wide Distributions Are Differently Influenced by Sex in Brain Cortexes and Gastrocnemius of the Alzheimer’s Disease PSAPP Mouse Model

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. ChIPSeq
2.3. Computational Pipeline
3. Results
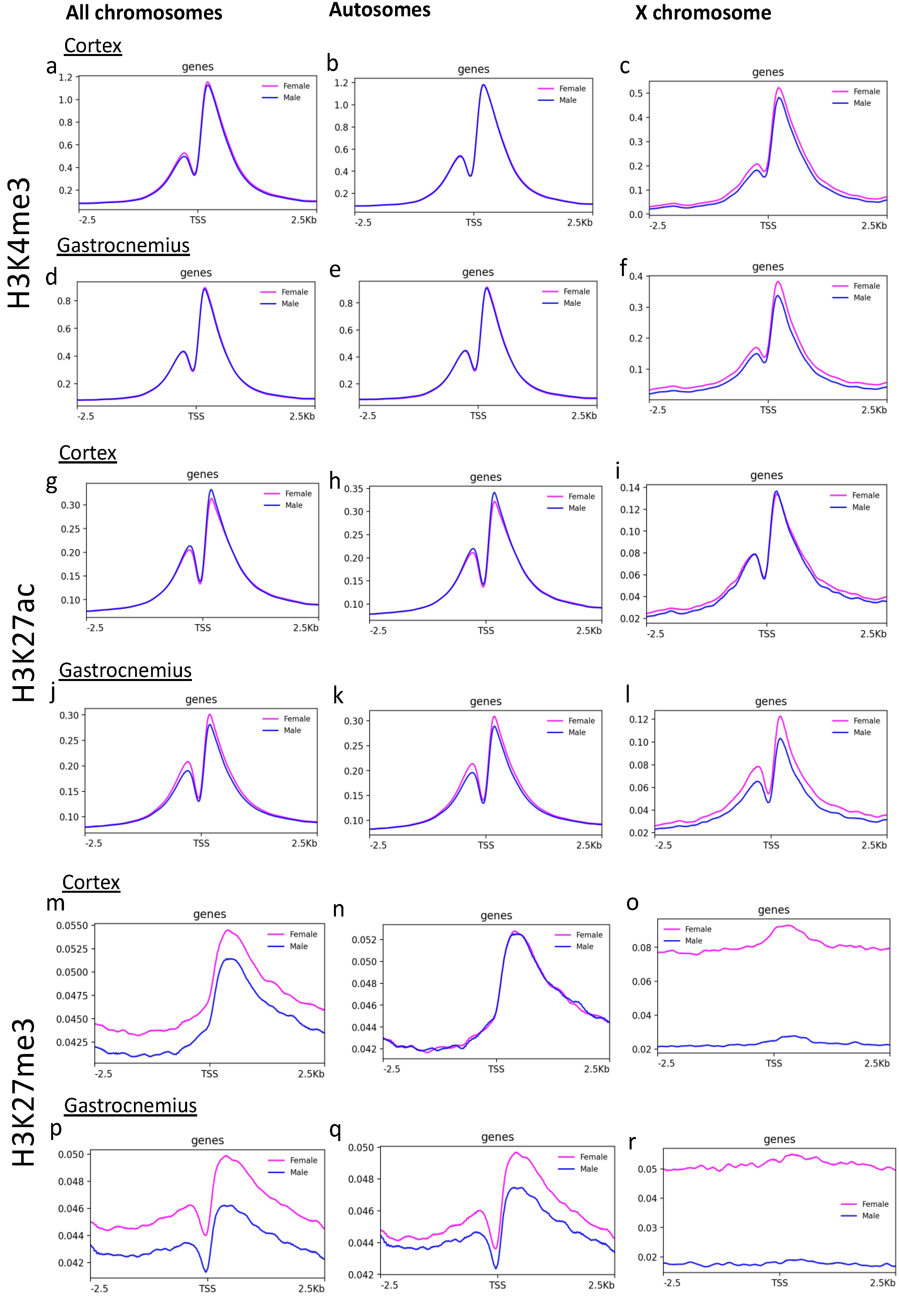
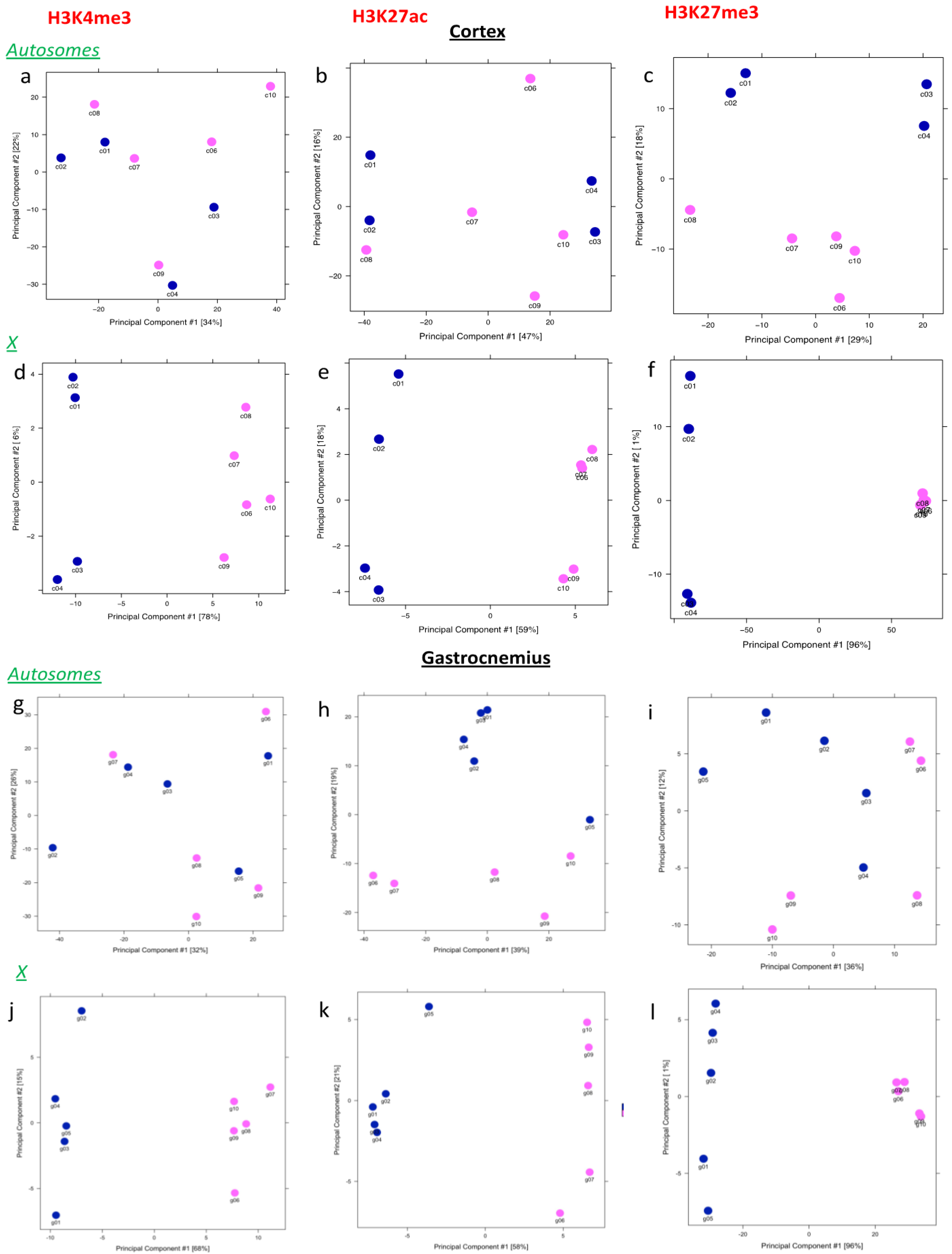
3.1. H3K4 and H3K27 me3 Analysis on X Chromosome Distinguishes Males and Females
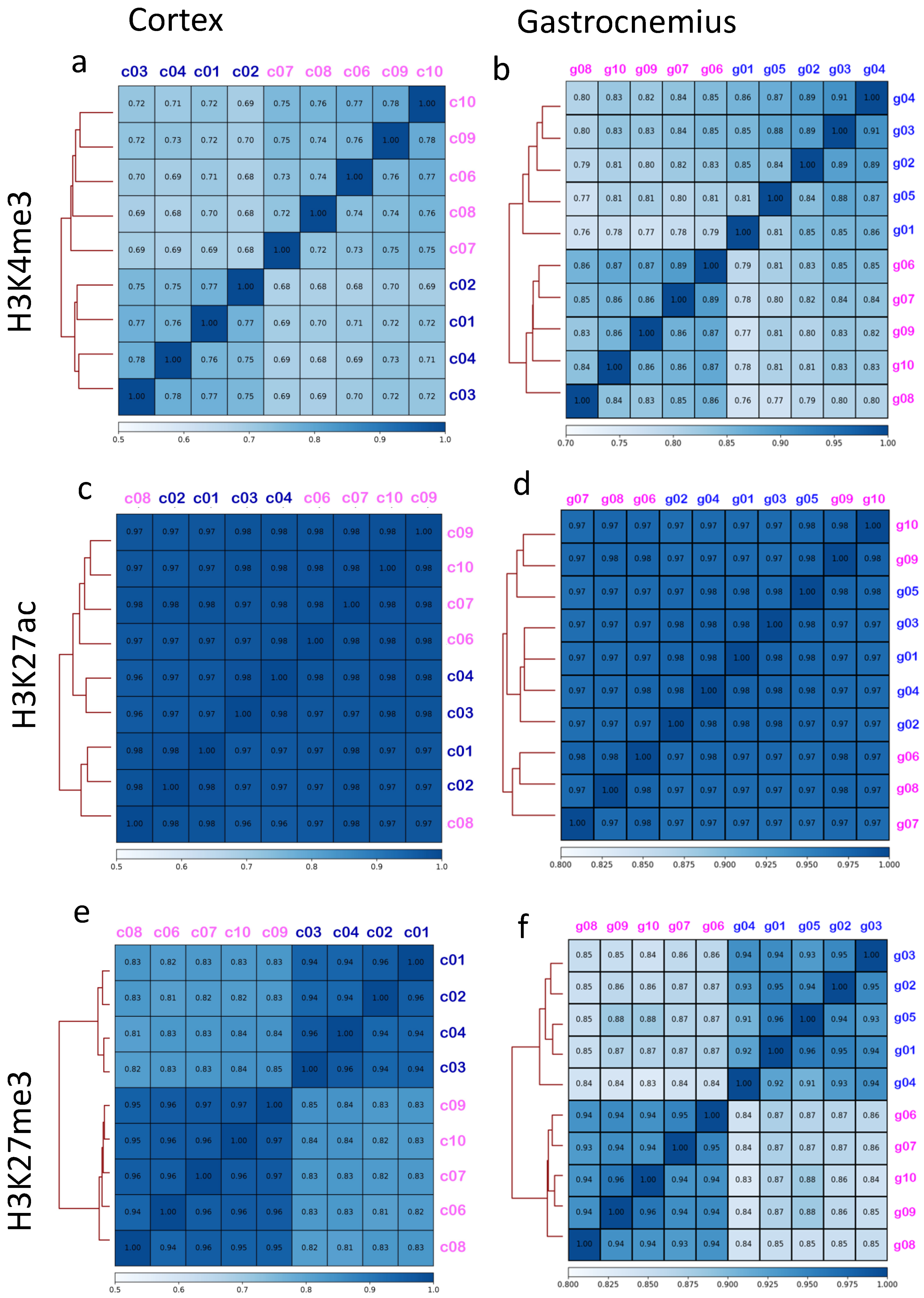
3.2. Brain Cortex Shows a Signal around the Transcription Starting Sites (TSS) That Is More Homogenous Than Gastrocnemius
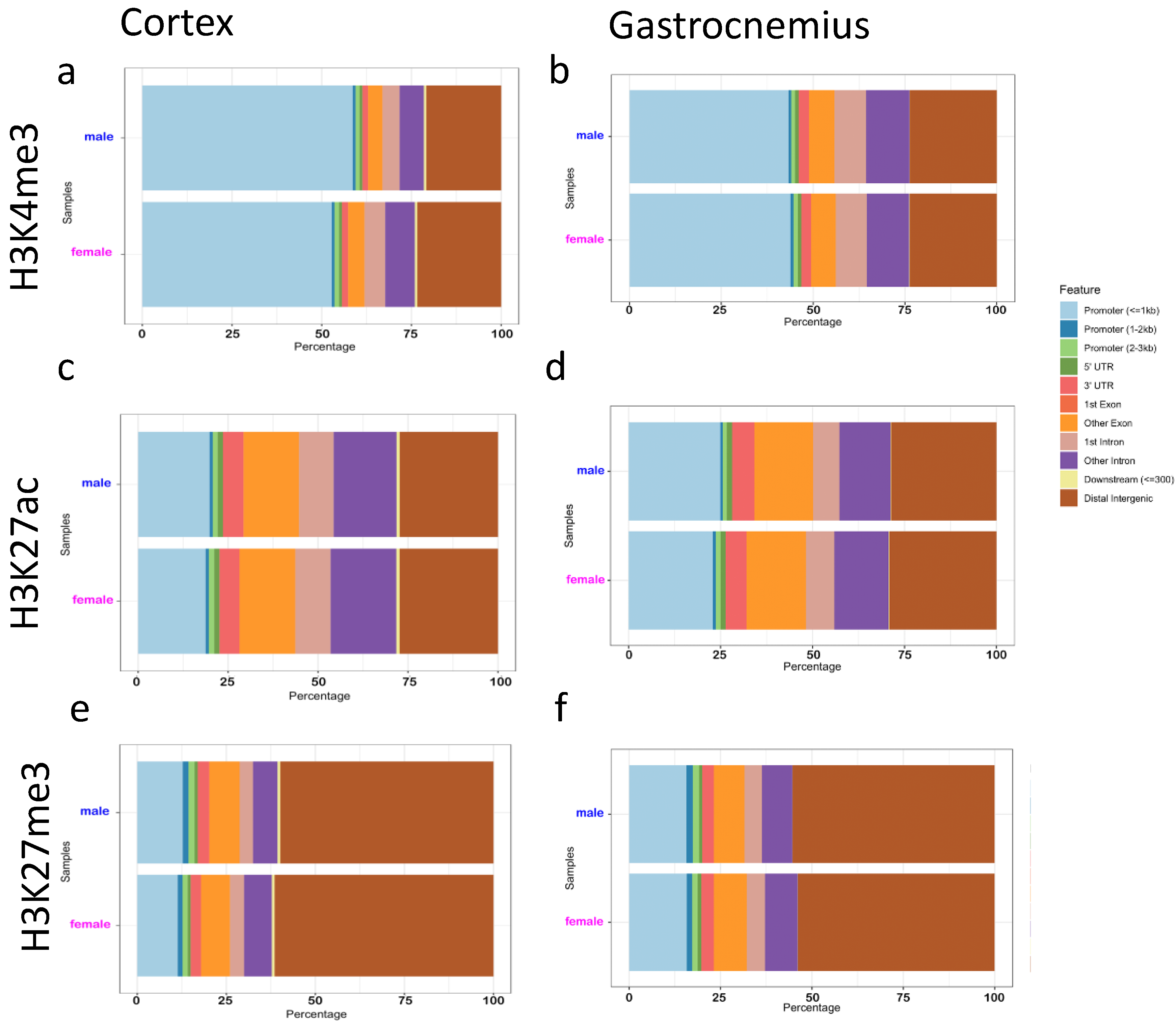
3.3. Binding Sites Are Differently Affected by Sex in Cortex and Gastrocnemius
3.4. Genes Involved in Cognitive Functions Show Different H3K27me3 Signal between Sex
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thies, W.; Bleiler, L. Alzheimer’s Association 2013 Alzheimer’s Disease Facts and Figures. Alzheimer’s Dement. 2013, 9, 208–245. [Google Scholar] [CrossRef]
- Gauthier, S.; Rosa-Neto, P.; Morais, J.; Webster, C. World Alzheimer Report 2021: Journey through the Diagnosis of Dementia; Alzheimer’s Disease International: London, UK.
- Beam, C.R.; Kaneshiro, C.; Jang, J.Y.; Reynolds, C.A.; Pedersen, N.L.; Gatz, M. Differences Between Women and Men in Incidence Rates of Dementia and Alzheimer’s Disease. J. Alzheimer’s Dis. 2018, 64, 1077–1083. [Google Scholar] [CrossRef]
- Andersen, K.; Launer, L.; Dewey, M.; Letenneur, L.; Ott, A.; Copeland, J.; Dartigues, J.; Kragh-Sorensen, P.; Baldereschi, M.; Brayne, C.; et al. Gender Differences in the Incidence of AD and Vascular Dementia: The EURODEM Studies. EURODEM Incidence Research Group. Neurology 1999, 53, 1992–1997. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.; Kienbacher, E.; Benke, T.; Dal-Bianco, P.; Delazer, M.; Ladurner, G.; Jellinger, K.; Marksteiner, J.; Ransmayr, G.; Schmidt, H.; et al. Sex Differences in Alzheimer’s Disease. Neuropsychiatry 2008, 22, 1–15. [Google Scholar]
- Tschanz, J.T.; Corcoran, C.D.; Schwartz, S.; Treiber, K.; Green, R.C.; Norton, M.C.; Mielke, M.M.; Piercy, K.; Steinberg, M.; Rabins, P.V.; et al. Progression of Cognitive, Functional and Neuropsychiatric Symptom Domains in a Population Cohort with Alzheimer’s Dementia The Cache County Dementia Progression Study. Am. J. Geriatr. Psychiatry 2011, 19, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-Q.; Mobley, W.C. Alzheimer Disease Pathogenesis: Insights From Molecular and Cellular Biology Studies of Oligomeric Aβ and Tau Species. Front. Neurosci. 2019, 13, 659. [Google Scholar] [CrossRef]
- Ordoñez-Gutierrez, L.; Fernandez-Perez, I.; Herrera, J.L.; Anton, M.; Benito-Cuesta, I.; Wandosell, F. AβPP/PS1 Transgenic Mice Show Sex Differences in the Cerebellum Associated with Aging. J. Alzheimer’s Dis. 2016, 54, 645–656. [Google Scholar] [CrossRef]
- Ogawa, Y.; Kaneko, Y.; Sato, T.; Shimizu, S.; Kanetaka, H.; Hanyu, H. Sarcopenia and Muscle Functions at Various Stages of Alzheimer Disease. Front. Neurol. 2018, 9, 710. [Google Scholar] [CrossRef] [PubMed]
- Gershoni, M.; Pietrokovski, S. The Landscape of Sex-Differential Transcriptome and Its Consequent Selection in Human Adults. BMC Biol. 2017, 15, 7. [Google Scholar] [CrossRef]
- Monteiro-Cardoso, V.; Castro, M.; Oliveira, M.M.; Moreira, P.; Peixoto, F.; Videira, R. Age-Dependent Biochemical Dysfunction in Skeletal Muscle of Triple-Transgenic Mouse Model of Alzheimer’s Disease. Curr. Alzheimer Res. 2015, 12, 100–115. [Google Scholar] [CrossRef] [PubMed]
- Schuh, R.A.; Jackson, K.C.; Schlappal, A.E.; Spangenburg, E.E.; Ward, C.W.; Park, J.H.; Dugger, N.; Shi, G.L.; Fishman, P.S. Mitochondrial Oxygen Consumption Deficits in Skeletal Muscle Isolated from an Alzheimer’s Disease-Relevant Murine Model. BMC Neurosci. 2014, 15, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Jiao, B.; Shen, L. The Epigenetics of Alzheimer’s Disease: Factors and Therapeutic Implications. Front. Genet. 2018, 9, 579. [Google Scholar] [CrossRef]
- Alagiakrishnan, K.; Gill, S.S.; Fagarasanu, A. Genetics and Epigenetics of Alzheimer’s Disease. Postgrad. Med. J. 2012, 88, 522–529. [Google Scholar] [CrossRef] [PubMed]
- Avner, P.; Heard, E. X-Chromosome Inactivation: Counting, Choice and Initiation. Nat. Rev. Genet. 2001, 2, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Keiser, A.A.; Wood, M.A. Examining the Contribution of Histone Modification to Sex Differences in Learning and Memory. Learn. Mem. 2019, 26, 318–331. [Google Scholar] [CrossRef] [PubMed]
- Ramzan, F.; Creighton, S.; Hall, M.; Baumbach, J.; Wahdan, M.; Poulson, S.; Michailidis, V.; Stefanelli, G.; Narkaj, K.; Tao, C.; et al. Sex-Specific Effects of the Histone Variant H2A.Z on Fear Memory, Stress-Enhanced Fear Learning and Hypersensitivity to Pain. Sci. Rep. 2020, 10, 14331. [Google Scholar] [CrossRef] [PubMed]
- Shen, E.; Ahern, T.; Cheung, I.; Straubhaar, J.; Dincer, A.; Houston, I.; de Vries, G.; Akbarian, S.; Forger, N. Epigenetics and Sex Differences in the Brain: A Genome-Wide Comparison of Histone-3 Lysine-4 Trimethylation (H3K4me3) in Male and Female Mice. Exp. Neurol. 2015, 268, 21–29. [Google Scholar] [CrossRef]
- Lawrence, M.; Daujat, S.; Schneider, R. Lateral Thinking: How Histone Modifications Regulate Gene Expression. Trends Genet. 2016, 32, 42–56. [Google Scholar] [CrossRef]
- Wang, C.M.; Tsai, S.N.; Yew, T.W.; Kwan, Y.W.; Ngai, S.M. Identification of Histone Methylation Multiplicities Patterns in the Brain of Senescence-Accelerated Prone Mouse. Biogerontology 2010, 11, 87–102. [Google Scholar] [CrossRef]
- Ryu, S.H.; Kang, K.; Yoo, T.; Joe, C.O.; Chung, J.H. Transcriptional Repression of Repeat-Derived Transcripts Correlates with Histone Hypoacetylation at Repetitive DNA Elements in Aged Mice Brain. Exp. Gerontol. 2011, 46, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Luo, M.; Jeong, M.; Rodriguez, B.; Xia, Z.; Hannah, R.; Wang, H.; Le, T.; Faull, K.F.; Chen, R.; et al. Epigenomic Profiling of Young and Aged HSCs Reveals Concerted Changes during Aging That Reinforce Self-Renewal. Cell Stem Cell 2014, 14, 673–688. [Google Scholar] [CrossRef] [PubMed]
- Dhawan, S.; Tschen, S.-I.; Bhushan, A. Bmi-1 Regulates the Ink4a/Arf Locus to Control Pancreatic β-Cell Proliferation. Genes Dev. 2009, 23, 906–911. [Google Scholar] [CrossRef]
- Cheung, I.; Shulha, H.P.; Jiang, Y.; Matevossian, A.; Wang, J.; Weng, Z.; Akbarian, S. Developmental Regulation and Individual Differences of Neuronal H3K4me3 Epigenomes in the Prefrontal Cortex. Proc. Natl. Acad. Sci. USA 2010, 107, 8824–8829. [Google Scholar] [CrossRef] [PubMed]
- Meyerhoff, J.; Muhie, S.; Chakraborty, N.; Naidu, L.; Sowe, B.; Hammamieh, R.; Jett, M.; Gautam, A. Microdissection of Mouse Brain into Functionally and Anatomically Different Regions. J. Vis. Exp. 2021, 168, e61941. [Google Scholar] [CrossRef]
- Cutler, A.; Corbett, A.; Pavlath, G. Biochemical Isolation of Myonuclei from Mouse Skeletal Muscle Tissue. Bio-Protocol 2017, 7, e2654. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, D.; Wilson, M.; Spyrou, C.; Brown, G.; Hadfield, J.; Odom, D. ChIP-Seq: Using High-Throughput Sequencing to Discover Protein-DNA Interactions. Methods 2009, 48, 240–248. [Google Scholar] [CrossRef]
- Amatori, S.; Persico, G.; Paolicelli, C.; Hillje, R.; Sahnane, N.; Corini, F.; Furlan, D.; Luzi, L.; Minucci, S.; Giorgio, M.; et al. Epigenomic Profiling of Archived FFPE Tissues by Enhanced PAT-ChIP (EPAT-ChIP) Technology. Clin. Epigenetics 2018, 10, 143. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and Accurate Short Read Alignment with Burrows-Wheeler Transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Quinlan, A.; Hall, I. BEDTools: A Flexible Suite of Utilities for Comparing Genomic Features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef]
- Stovner, E.; Sætrom, P. Epic2 Efficiently Finds Diffuse Domains in ChIP-Seq Data. Bioinformatics 2019, 35, 4392–4393. [Google Scholar] [CrossRef] [PubMed]
- Stark, R.; Brown, G. DiffBind: Differential Binding Analysis of ChIP-Seq Peak Data. Available online: https://bioconductor.org/packages/release/bioc/vignettes/DiffBind/inst/doc/DiffBind.pdf (accessed on 1 October 2021).
- Yu, G.; Wang, L.-G.; He, Q.-Y. ChIPseeker: An R/Bioconductor Package for ChIP Peak Annotation, Comparison and Visualization. Bioinformatics 2015, 31, 2382–2383. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, F.; Ryan, D.P.; Grüning, B.; Bhardwaj, V.; Kilpert, F.; Richter, A.S.; Heyne, S.; Dündar, F.; Manke, T. DeepTools2: A next Generation Web Server for Deep-Sequencing Data Analysis. Nucleic Acids Res. 2016, 44, W160–W165. [Google Scholar] [CrossRef] [PubMed]
- Rougeulle, C.; Chaumeil, J.; Sarma, K.; Allis, C.; Reinberg, D.; Avner, P.; Heard, E. Differential Histone H3 Lys-9 and Lys-27 Methylation Profiles on the X Chromosome. Mol. Cell. Biol. 2004, 24, 5475–5484. [Google Scholar] [CrossRef]
- Haizlip, K.M.; Harrison, B.C.; Leinwand, L.A. Sex-Based Differences in Skeletal Muscle Kinetics and Fiber-Type Composition. Physiology 2015, 30, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Welle, S.; Tawil, R.; Thornton, C.A. Sex-Related Differences in Gene Expression in Human Skeletal Muscle. PLoS ONE 2008, 3, e1385. [Google Scholar] [CrossRef]
- Ingenuity Downstream Effects Analysis in IPA®. Available online: http://pages.ingenuity.com/rs/ingenuity/images/0812%20downstream_effects_analysis_whitepaper.pdf (accessed on 4 November 2021).
- Takehashi, M.; Tanaka, S.; Masliah, E.; Ueda, K. Association of Monoamine Oxidase A Gene Polymorphism with Alzheimer’s Disease and Lewy Body Variant. Neurosci. Lett. 2002, 327, 79–82. [Google Scholar] [CrossRef]
- Bodily, P.M.; Fujimoto, M.S.; Page, J.T.; Clement, M.J.; Ebbert, M.T.W.; Ridge, P.G. A Novel Approach for Multi-SNP GWAS and Its Application in Alzheimer’s Disease. BMC Bioinform. 2016, 17, 455–463. [Google Scholar] [CrossRef]
- Alsema, A.M.; Jiang, Q.; Kracht, L.; Gerrits, E.; Dubbelaar, M.L.; Miedema, A.; Brouwer, N.; Hol, E.M.; Middeldorp, J.; van Dijk, R.; et al. Profiling Microglia From Alzheimer’s Disease Donors and Non-Demented Elderly in Acute Human Postmortem Cortical Tissue. Front. Mol. Neurosci. 2020, 13, 134. [Google Scholar] [CrossRef] [PubMed]
- Quitterer, U.; AbdAlla, S. Improvements of Symptoms of Alzheimer’s Disease by Inhibition of the Angiotensin System. Pharmacol. Res. 2020, 154, 104230. [Google Scholar] [CrossRef]
- Dos Santos, J.; Vizuete, A.; Hansen, F.; Biasibetti, R.; Gonçalves, C. Early and Persistent O-GlcNAc Protein Modification in the Streptozotocin Model of Alzheimer’s Disease. J. Alzheimer’s Dis. 2018, 61, 237–249. [Google Scholar] [CrossRef]
- Holmes, C.; Arranz, M.; Collier, D.; Powell, J.; Lovestone, S. Depression in Alzheimer’s Disease: The Effect of Serotonin Receptor Gene Variation. Am. J. Med. Genet. 2003, 119B, 40–43. [Google Scholar] [CrossRef]
- He, X.; Isaacs, C.; Yang, S. Roles of Mitochondrial 17β-Hydroxysteroid Dehydrogenase Type 10 in Alzheimer’s Disease. J. Alzheimer’s Dis. 2018, 62, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Chu, J.; Wisniewski, T.; Praticò, D. GATA1-Mediated Transcriptional Regulation of the γ-Secretase Activating Protein Increases Aβ Formation in Down Syndrome. Ann. Neurol. 2016, 79, 138–143. [Google Scholar] [CrossRef]
- Giuffrida, M.L.; Copani, A.; Rizzarelli, E. A Promising Connection between BDNF and Alzheimer’s Disease. Aging 2018, 10, 1791–1792. [Google Scholar] [CrossRef] [PubMed]
- Panula, P.; Rinne, J.; Kuokkanen, K.; Eriksson, K.; Sallmen, T.; Kalimo, H.; Relja, M. Neuronal Histamine Deficit in Alzheimer’s Disease. Neuroscience 1998, 82, 993–997. [Google Scholar] [CrossRef]
- Wang, W.; Tanokashira, D.; Fukui, Y.; Maruyama, M.; Kuroiwa, C.; Saito, T.; Saido, T.; Taguchi, A. Serine Phosphorylation of IRS1 Correlates with Aβ-Unrelated Memory Deficits and Elevation in Aβ Level Prior to the Onset of Memory Decline in AD. Nutrients 2019, 11, 1942. [Google Scholar] [CrossRef]
- Wang, J.; Tanila, H.; Puoliväli, J.; Kadish, I.; van Groen, T. Gender Differences in the Amount and Deposition of Amyloidbeta in APPswe and PS1 Double Transgenic Mice. Neurobiol. Dis. 2003, 14, 318–327. [Google Scholar] [CrossRef]
- Jiao, S.; Bu, X.; Liu, Y.; Zhu, C.; Wang, Q.; Shen, L.; Liu, C.; Wang, Y.; Yao, X.; Wang, Y. Sex Dimorphism Profile of Alzheimer’s Disease-Type Pathologies in an APP/PS1 Mouse Model. Neurotox. Res. 2016, 29, 256–266. [Google Scholar] [CrossRef]
- Li, X.; Feng, Y.; Wu, W.; Zhao, J.; Fu, C.; Li, Y.; Ding, Y.; Wu, B.; Gong, Y.; Yang, G.; et al. Sex Differences between APPswePS1dE9 Mice in A-Beta Accumulation and Pancreatic Islet Function during the Development of Alzheimer’s Disease. Lab. Anim. 2016, 50, 275–285. [Google Scholar] [CrossRef]
- Yang, J.-T.; Wang, Z.-J.; Cai, H.-Y.; Yuan, L.; Hu, M.-M.; Wu, M.-N.; Qi, J.-S. Sex Differences in Neuropathology and Cognitive Behavior in APP/PS1/Tau Triple-Transgenic Mouse Model of Alzheimer’s Disease. Neurosci. Bull. 2018, 34, 736–746. [Google Scholar] [CrossRef]
- Lopes-Ramos, C.M.; Chen, C.Y.; Kuijjer, M.L.; Paulson, J.N.; Sonawane, A.R.; Fagny, M.; Platig, J.; Glass, K.; Quackenbush, J.; DeMeo, D.L. Sex Differences in Gene Expression and Regulatory Networks across 29 Human Tissues. Cell Rep. 2020, 31, 107795. [Google Scholar] [CrossRef]
- Andreano, J.M.; Cahill, L. Sex Influences on the Neurobiology of Learning and Memory. Learn. Mem. 2009, 16, 248–266. [Google Scholar] [CrossRef]
- Skuse, D.H. X-Linked Genes and Mental Functioning. Hum. Mol. Genet. 2005, 14, 27–32. [Google Scholar] [CrossRef]
- Laumonnier, F.; Cuthbert, P.C.; Grant, S.G.N. The Role of Neuronal Complexes in Human X-Linked Brain Diseases. Am. J. Hum. Genet. 2007, 80, 205–220. [Google Scholar] [CrossRef] [PubMed]
- Campbell, R.; Wood, M. How the Epigenome Integrates Information and Reshapes the Synapse. Nat. Rev. Neurosci. 2019, 20, 133–147. [Google Scholar] [CrossRef]
- Gupta, S.; Kim, S.Y.; Artis, S.; Molfese, D.L.; Schumacher, A.; Sweatt, J.D.; Paylor, R.E.; Lubin, F.D. Histone Methylation Regulates Memory Formation. J. Neurosci. 2010, 30, 3589–3599. [Google Scholar] [CrossRef] [PubMed]
- Berletch, J.B.; Yang, F.; Xu, J.; Carrel, L.; Disteche, C.M. Genes That Escape from X Inactivation. Hum. Genet. 2011, 130, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Greenfield, A.; Carrel, L.; Pennisi, D.; Philippe, C.; Quaderi, N.; Siggers, P.; Steiner, K.; Tam, P.P.L.; Monaco, A.P.; Willard, H.F.; et al. The UTX Gene Escapes X Inactivation in Mice and Humans. Hum. Mol. Genet. 1998, 7, 737–742. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Histone Modification | Chromosomes | N° of db Sites in Cortex | N° db Sites in Gastrocnemius |
|---|---|---|---|
| Autosomes | 2 | 139 | |
| H3K4me3 | X chromosome | 65 | 102 |
| Autosomes | 2 | 781 | |
| H3K27ac | X chromosome | 56 | 69 |
| H3K27me3 | Autosomes | 5 | 206 |
| X chromosome | 2783 | 855 |
| Cortex Diseases or Functions (H3K27me3 X) | p-Value | Predicted Activation State | Activation Z-Score | Molecules | N° of Molecules |
|---|---|---|---|---|---|
| Spatial learning | 2.70 × 10−5 | Increased | 2.646 | AP1S2,ARHGEF9,CYBB,DLG3,Gprasp2,GRIPAP1,HTR2C, KDM5C, MECP2, OPHN1,PHF8,SLC6A8,UBE2A, ZDHHC9 | 14 |
| Growth of neurites | 4.98 × 10−3 | Increased | 2.138 | AR,ARX,CCDC120,CDKL5,DCX,EFNB1,ELK1,FRMD7, GJB1, MAO, MID1,OGT,PLXNA3,RAB33A,SLC25A5, SNX12,SYN1,TLR7,TRPC5 | 19 |
| Organization of cytoskeleton | 1.17 × 10−7 | Increased | 2.074 | AGTR2,AMOT,AR,ARHGAP4,ARHGAP6,ARHGEF9, ATP7A, BRWD3,CAPN6,CDK16,CDKL5,CETN2,CUL4B, CXCR3,CYBB, DCX,DGKK,DLG3,DOCK11,EFNB1,ELK1, F8A1 (includes others),FGD1,FLNA,FRMD7,GATA1,GDI1,GJB1,GPM6B, Gprasp2,HDAC6,HDAC8,HPRT1, IL1RAPL1,KDM5C,MAOA, MECP2,MID1,MID1IP1, mir-384,MPP1,MTM1,NR0B1,OFD1, OGT,OPHN1,PAK3, PCYT1B,PLS3,PLXNA3,PLXNB3,POF1B, PQBP1,RAB33A, RPGR,RPS6KA3,SH3KBP1,SHROOM2, SHROOM4, SLITRK2,SYN1,TLR7,Tmsb4x (includes others), TRPC5, USP9X | 65 |
| Organization of cytoplasm | 8.76 × 10−7 | Increased | 2.074 | AGTR2,AMOT,AR,ARHGAP4,ARHGAP6,ARHGEF9, ATP7A, BRWD3,CAPN6,CDK16,CDKL5,CETN2,CUL4B,CXCR3,CYBB, DCX,DGKK,DLG3,DOCK11,EFNB1,ELK1,F8A1(includes others),FGD1,FLNA,FRMD7,GATA1,GDI1,GJB1,GPM6B, Gprasp2,HCFC1,HDAC6,HDAC8,HPRT1,HSD17B10, IL1RAPL1,KDM5C,MAOA,MECP2,MID1,MID1IP1,mir-384, MPP1,MTM1,NR0B1,OFD1,OGT,OPHN1,PAK3,PCYT1B,PLS3,PLXNA3,PLXNB3,POF1B,PQBP1,RAB33A,RPGR,RPS6KA3, SH3KBP1,SHROOM2, SHROOM4, SLITRK, SYN1,TLR7, Tmsb4x (includes others),TRPC5,USP9X | 67 |
| Tremor | 1.14 × 10−3 | Decreased | −2.011 | ARAF,CA5B,GABRQ,GJB1,GPM6B,GRIA3,IKBKG,MECP2, PLP1, TIMP1 | 10 |
| Differentiation of Th2 cells | 1.09 × 10−2 | Decreased | −2.236 | FOXP3,GATA1,IL13RA2,let-7,TLR7 | 5 |
| Movement Disorders | 2.29 × 10−3 | Decreased | −2.242 | ABCB7,AIFM1,AMER1,AP1S2,AR,ARAF,ARHGEF9,ARMCX2, AR, ATP6AP2,BCAP31,CA5B,CDKL5,CETN2,CXCR3,F8A1 (includes others),GABRQ,GJB1,GPM6B,GRIA3,GRPR,HPRT1, HTR2C,IDS,IGSF1,IKBKG,KCND1,MAOA,MECP2,OGT,PDK3, PGK1,PGRMC1,PLP1,PRKX,PTCHD1,RGN,RS1,SRPX,SRPX2, SYN1,SYTL4,TIMP1,Tmsb4x (includes others),XIAP | 45 |
| Motor dysfunction or movement disorder | 1.72 × 10−3 | Decreased | −2.666 | ABCB7,AIFM1,AMER1,AP1S2,AR,ARAF,ARHGEF9,ARMCX2, AR,ATP6AP2,BCAP31,CA5B,CDKL5,CETN2,CXCR3,F8A1 (includes others),GABRQ,GJB1,GPM6B,GRIA3,GRPR,HPRT1, HTR2C,IDS,IGSF1,IKBKG,KCND1,MAOA,MECP2,MTM1, OGT, PDK3,PGK1,PGRMC1,PLP1,PRKX,PTCHD1,RGN,RS1, SRPX,SRPX2,SYN1, SYTL4,TIMP1,Tmsb4x (includes others), XIAP | 46 |
| Gastrocnemius Diseases or Functions (H3K27me3 X) | p-Value | Predicted Activation State | Activation Z-Score | Molecules | N° of Molecules |
|---|---|---|---|---|---|
| Organization of actin cytoskeleton | 27.23 × 10−3 | Increased | 2.000 | ARHGAP4,EFNB1,FGD1,GPM6B,MSN,OPHN1,PAK3,PLS3, Tmsb4x (includes others) | 9 |
| Abdominal cancer | 1.41 × 10−5 | Decreased | −2.000 | ABCB7,ACE2,AIFM1,AMER1,ARHGAP36,ARHGAP4,ARHGEF9, ARMCX1,ARMCX2,ARMCX3,ARMCX4,ARMCX5,ARX, ASB9, ATP2B3,AVPR2,AWAT2,BCOR,BCORL1,BEX1,BGN, BMX,BRS3, BTK,CA5B,CACNA1F,CCDC160,CD40LG, CDX4,CHRDL1, CLDN2,CNGA2,CNKSR2,COL4A5,DACH2,DCAF12L2,DGAT2L6,DOCK11, DUSP9, EFNB1,EGFL6,ELF4,ERCC6L,FAM155B, FGD1,FOXO4,FOXR2,FRMD7,FRMPD3,GABRA3,GABRQ,GATA1, GDPD2,GJB1,GPC3,GPC4,GPM6,GPR101,GPR143, GPR50,GRIA3, GRPR,GSPT2,GUCY2F,HCFC1,HDAC8, HPRT1,HSD17B1,HTR2C, IDH3G,IGSF1,IL13RA1,IL1RAPL1,IL1RAPL2, IQSEC2,IRAK1, KCND1,KCNE5,KLF8,KLHL15,L1CAM, LANCL3,LHFPL1, LONRF3,MAGEA10, MAGEA11,MAGED1,MAGEE1,MAGEE2, MAP7D2,MBNL3,mir-452,MSN,NAA10,NAP1L2,NEXMIF,NONO, NRK,NYX,OGT,OPHN1,OPN1LW,OTUD6A,PAK3,PCDH11X, PCDH19,PDK3,PDZD4,PHF6,PHKA1,PLAC1,PLS3,PLXNA3, PLXNB3,PNMA3,PNMA5,PRKX,PRPS1,PRPS2,PTCHD1, RAB33A, RAI2,RAP2C,RBM41,RNF128,RPS4Y1,RRAGB,SLC16A2,SLC25A43,SLC38A5,SLC6A8,SLC7A3,SLITRK4,SOWAHD,SOX3,SRPX, STARD8,SYN1,SYTL4,TAB3,TAF1,TBX22,TCEANC,THOC2, TMEM164,TMEM255A,TMEM47,TREX2, TRPC5,TSC22D3,TSPYL2, UPF3B,USP51,UTP14A,VGLL1,YIPF6,ZC4H2, ZCCHC12, ZDHHC9,ZFX,ZIC3,ZMYM3 | 162 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Casciaro, F.; Persico, G.; Rusin, M.; Amatori, S.; Montgomery, C.; Rutkowsky, J.R.; Ramsey, J.J.; Cortopassi, G.; Fanelli, M.; Giorgio, M. The Histone H3 K4me3, K27me3, and K27ac Genome-Wide Distributions Are Differently Influenced by Sex in Brain Cortexes and Gastrocnemius of the Alzheimer’s Disease PSAPP Mouse Model. Epigenomes 2021, 5, 26. https://0-doi-org.brum.beds.ac.uk/10.3390/epigenomes5040026
Casciaro F, Persico G, Rusin M, Amatori S, Montgomery C, Rutkowsky JR, Ramsey JJ, Cortopassi G, Fanelli M, Giorgio M. The Histone H3 K4me3, K27me3, and K27ac Genome-Wide Distributions Are Differently Influenced by Sex in Brain Cortexes and Gastrocnemius of the Alzheimer’s Disease PSAPP Mouse Model. Epigenomes. 2021; 5(4):26. https://0-doi-org.brum.beds.ac.uk/10.3390/epigenomes5040026
Chicago/Turabian StyleCasciaro, Francesca, Giuseppe Persico, Martina Rusin, Stefano Amatori, Claire Montgomery, Jennifer R. Rutkowsky, Jon J. Ramsey, Gino Cortopassi, Mirco Fanelli, and Marco Giorgio. 2021. "The Histone H3 K4me3, K27me3, and K27ac Genome-Wide Distributions Are Differently Influenced by Sex in Brain Cortexes and Gastrocnemius of the Alzheimer’s Disease PSAPP Mouse Model" Epigenomes 5, no. 4: 26. https://0-doi-org.brum.beds.ac.uk/10.3390/epigenomes5040026